黄海北部近岸海域渔业生物群落结构
2024-05-20崔培东卞晓东张雨轩单秀娟金显仕赵永松王惠宾
崔培东, 卞晓东, 张雨轩, 单秀娟,金显仕*, 赵永松, 王惠宾
(1. 上海海洋大学海洋科学学院,上海 201306;2. 中国水产科学研究院黄海水产研究所,农业农村部海洋渔业可持续发展重点实验室,山东省渔业资源与生态环境重点实验室,山东 青岛 266071;3. 山东长岛近海渔业资源国家野外科学观测研究站,山东 烟台 265800)
黄海北部近岸海域岛屿密布,有大洋河、辽河、鸭绿江等诸多河流汇入,同时受辽南沿岸流、辽东湾沿岸流、黄海暖流及黄海冷水团的交错影响,有机质和营养盐丰富,诸多海洋生物在此栖息繁衍,曾是黄海北部的重要产卵场和重要的渔业水域[1-5]。该海域地处暖温带又属近岸,海陆交互频繁,自然界各圈层相互影响活跃,气候复杂多变[3];同时受到人类捕捞活动、伏季休渔制度的实施、增殖放流活动和近海增养殖业发展等多重因素影响,渔业生物群落结构常呈现明显年际变动。
国内已开展过多次黄海北部近岸海域渔业生物资源调查,对海区渔获种类组成、区系特征、资源数量变动和群落结构等进行了相关研究[1,3,5-11]。20 世纪50、60 年代海域捕捞生产处于初级阶段,渔业资源相对稳定[3];随着渔业技术的不断发展和捕捞强度的增加,海域底层经济渔业资源逐步衰退,20 世纪60 年代后陆续转为开发大中型中上层渔业资源;20 世纪90 年代又对小型中上层鱼类进行大规模开发利用,渔业资源结构进一步发生改变[3]。在长期高强度的渔业捕捞压力下,传统经济种类亲体、当年生幼鱼均遭受过度捕捞,影响了渔业资源的可持续利用[1]。20 世纪90 年代以后,海域渔业资源逐渐趋于低龄化、小型化和低值化,短生命周期、低营养级的小型中上层鱼类、头足类和小型虾、蟹类逐步代替大中型经济种类[8]。在巨大捕捞压力下,小黄鱼(Larimichthys polyactis)、蓝点马鲛(Scomberomorus niphonius)等传统捕捞对象在2010—2017 年已基本无规模渔汛,鳀(Engraulis japonicus)捕捞量也呈下降趋势,个别年份上半年双拖网作业更是呈现停产状态[11]。历经几十年的开发利用,黄海北部近岸海域渔业生物群落结构已发生改变,有关该海域渔业生物群落结构变化的研究前述各调查虽均有涉及,但尚缺乏主要产卵期较为连续的观测资料,且有关海域生态环境及其对群落结构影响机制的描述较少。
因此,本实验于2021 年4—11 月在黄海北部近岸海域产卵场展开调查,开展了6 航次11 站次渔业资源调查和20 站次栖息环境调查,运用典范对应分析法对海区渔业资源种类组成及数量分布、群落结构稳定性及月际变动、生物多样性变化及群落结构时空变化与环境因子的关系进行了分析,以期阐明海域渔业资源种类组成和数量分布特征,并为海域渔业资源合理开发利用提供生态学基础数据。
1 材料与方法
1.1 数据来源与调查方法
2021 年4 月(4 月18—21 日)、5—6 月(5 月28 日—6 月1 日)、7 月(7 月21—24 日)、8 月(8月17—20 日)、9 月(9 月11—14 日)、10—11 月(10 月31 日—11 月4 日)于黄海北部近岸海域进行了底拖网调查。调查区域为38.75°~39.25°N,122°~124°E,总计11 站(图1,S1~S11)。期间采用 “中渔科102” 调查船单船底拖网取样,网目200 mm×440 目,网具规格46.6 m×58 m,网口高、宽度6 m × 21 m,上纲长度45 m,底纲长度47 m,囊网网目20 mm×620 目,每站拖网1 h,拖速为3 kn,网口宽度根据水深和曳纲长度,一般为18~22 m。现场鉴定和记录每站渔获物的种类与数量。调查与采样方法按照《海洋调查规范第 6 部分:海洋生物调查》(GB/T 12763.6—2007)执行。物种分类依据《中国海洋生物名录》[12]及《中国北部海洋无脊椎动物》[13]。实验期间,操作者严格遵守实验动物福利伦理规范。

图1 黄海北部近岸海域生态调查站位及拖网调查站位分布全部站位进行生态环境调查;S1~S11 表示拖网调查站位。Fig. 1 Distribution of ecological survey stations and trawl survey stations in offshore waters of the North Yellow Sea Ecological survey was carried out in all stations; S1-S11. trawl survey station.
将调查所获的鱼类资源按照FishBase (https: //www.fishbase.in/search.php)、Taiwan Fish Database(http: //fishdb.sinica. edu.tw)以及刘静等[14]划分的栖息类型判断分布水层。依据Tian 等[15]的研究将所获鱼类划分不同适温类型。参考《中国经济动物志-海产鱼类》[16]、《黄渤海近岸水域生态环境与生物群落》[3]等相关研究来衡量各渔业资源的经济价值。
对海域20 站位进行生态环境调查(图1),基本环境要素(温度、盐度)使用美国 Seabird 19 plus 及德国 Sea & Sun Technology CTD 60M 温盐深仪现场测量。分别选择海水表层0.5 m 内和海水底层0.5 m 内的温度、盐度平均值作为表、底温盐数据。根据船载声呐读取各站位的实测水深。使用采水器采取500 mL 表层海水并过滤至玻璃纤维滤膜(孔径 0.7 μm)上以获取表层叶绿素a(Chl.a)样品,冷冻保存带回实验室,用 90 %丙酮溶液于-20 °C 环境下冷冻萃取滤膜12 h,后放入Turner Designs Trilogy 测定Chl.a浓度。
由于7 月航次仪器故障,因此当月表层温度和叶绿素数据选用美国国家海洋和大气管理局(National Oceanic and Atmospheric Administration,NOAA) (https: //oceanwatch.pifsc.noaa.gov/erddap/griddap/index)所提供的月平均数据,底层温度和盐度数据选用哥白尼海洋环境监测服务中心(Copernicus Marine Environment Monitoring Service,CMEMS) (https: //resources.marine.copernicus.eu/product-detail/GLOBAL_ANALYSIS_FORECAST_PHY_001_024/INFORMATION)所提供的月平均数据,并应用ArcGIS 10.2 进行克里金空间插值统一空间分辨率。
1.2 群落物种优势度及相对资源密度
采用Pianka[17]的相对重要性指数(index of relative importance,IRI)作为群落优势度指标:
式中,N和W分别为每种渔获生物占所捕总量的个体数百分比和重量百分比,F为出现频率百分比。将IRI≥1 000 的种类定义为优势种,100 ≤IRI < 1 000 的种类定义为重要种,10≤IRI<100 的种类定义为常见种,1≤IRI<10 的种类定义为一般种,IRI<1 的种类定义为少见种[3]。本研究将IRI ≥10 (包括优势种、重要种和常见种)的渔业生物(鱼类、甲壳类及头足类)视为群落的主要种类[3]。
考虑到底拖网的选择作用,根据各站位扫海面积和各种类的可捕系数对现存渔业生物资源密度进行扫海面积法估算[18-20]:
式中,ρ为资源密度(单位面积资源生物量,kg/m2),C为平均每小时拖网渔获量(kg/h),a为每小时扫海面积(m2/h),q为捕捞系数。网具扫海面积(a)根据网口宽度(平均为0.02 km)与每小时的拖曳距离(5.556 km)计算,因此每网扫海面积为0.11 km2。中上层鱼类的可捕系数取0.3,近底层鱼类、枪乌贼类、口虾蛄(Oratosquilla oratoria)和虾类取0.5,底层鱼类、蛸类、蟹类等取0.8[8,21]。
1.3 群落结构生物多样性及稳定性
采用4 种生境内的多样性(within-habitat diversity),即α 多样性测度方法:
Shannon-Wiener 多样性指数H′[22]计算公式:
Margalef 丰富度指数D[23]计算公式:
Pielou 均匀度指数J′[24]计算公式:
Simpson 优势集中度指数λ[25]计算公式:
上述各式中,S为渔业生物种类总数,N为渔获物生物总生物量。Pi为群落中第i种的生物量占所有物种总生物量的比例。
应用丰度-生物量比较曲线(abundance - biomass comparison curves, ABC 曲线)来分析海域渔业生物群落结构的稳定性[26]。以W(W-statistic)作为ABC 曲线的统计量:
式中,Bi和Ai为 ABC 曲线中种类序号所对应的生物量和丰度的累积百分比,S为出现的物种数。该方法将生物量优势度曲线和丰度优势度曲线在同一坐标系中进行比较,依两条曲线的分布情况来判断群落所处干扰状况。当生物量优势曲线在丰度优势度曲线之上时W为正,反之为负。ABC曲线绘制和W统计量的计算均应用Primer 5.0 软件。
1.4 群落结构的时空变化及其与环境因子之间的相互关系
以春季(4 月及5—6 月调查)、夏季(7—8 月调查)、秋季(9—11 月调查)作为时间尺度,以各采样站位作为空间尺度。采用种类更替率E和Jaccard 群落种类组成相似性指数(coefficient of community, CC)[27]来表征群落结构季节间的物种更替[8,28]:
式中,E为种类更替率,A为季节间种类增加与减少数之和,即两季节间不同种类数,B为季节间相同的种类数。
式中,Ss为两个比较季节间共有种类数,Sj和Sk为两个比较季节各自拥有的种类数。当CC 为0~0.25 时, 群落间极不相似;当CC 为0.25~0.50时,群落间中等不相似;当CC 为0.50~0.75 时,群落间中等相似;当CC 为0.75~1.00 时,群落间极相似[27,29]。
应用Primer 5.0 软件进行CLUSTER 等级聚类和多维标度分析(non-metric multidimensional scaling ,NMDS),分别将春、夏、秋季各站点依物种丰度(4次方根转换)划分不同的群落结构。进一步应用单因子相似性分析(ANOSIM)对不同组群的群落结构进行差异显著性检验,并利用相似性百分比方法(SIMPER)检验群落结构组内相似的典型种,以及构成组间相异的分歧种[30-31]。
选取各样方中出现频率≥5%的渔业生物丰度数据作为物种矩阵[32],并进行[lg(x+1)] 转换以降低极端值影响;海水表层温度(SST)、海水底层温度(SBT)、海水表层盐度(SSS)、海水底层盐度(SBS)、深度(Depth)、Chl.a等作为环境因子矩阵,对除Chl.a以外的环境因子采取[lg(x+1)] 转换以使其符合正态分布[33-35]。首先,对物种数据进行趋势对应分析(detrended correspondence analysis,DCA),预分析显示最大梯度为4.33,表明数据更适合单峰模型进行排序[36],因此选择应用典范对应 分 析(canonical correspondence analysis,CCA)探究群落结构与环境因子的相互关系。
2 结果
2.1 黄海北部近岸海域物种组成及相对资源密度分布
本次资源调查捕获鱼类27 656 尾,无脊椎动物68 206 尾,共计89 种。包括鱼类50 种,隶属于12 目33 科48 属;无脊椎动物39 种,隶属于18 目30 科37 属,其中甲壳类及头足类25 种,共计11 目23 科30 属。底层鱼类主要包括大泷六线鱼(Hexagrammos otakii)、细纹狮子鱼(Liparis tanakae)、方氏云鳚(Pholis fangi)、黄鮟鱇(Lophius litulon)和绒杜父鱼(Hemitripterus villosus)等。中上层鱼类主要包括鳀、蓝圆鲹(Decapterus maruadsi)和黄鲫(Setipinna tenuifilis)等。头足类主要包括日本枪乌贼(Loligo japonica)和太平洋褶柔鱼(Todarodes pacificus)等。甲壳类主要包括脊腹褐虾(Crangon affinis)、戴氏赤虾(Metapenaeopsis dalei)和隆背黄道蟹(Cancer gibbosulus)等。本次调查5—6 月渔获资源丰度在全年占比最高,其次为8 月、10—11 月、9 月、7 月和4 月,其中4月黄鲫占比最高,5—6 月脊腹褐虾占比最高,7、8、9、10—11 月皆为鳀占比最高 (图2-a)。本次调查5—6 月渔获资源生物量(kg)在全年占比最高,其次为8 月、7 月、10—11 月、9 月和4 月,其中4 月黄鲫占比最高,5—6 月海葵(目) (Actiniaria)占比最高,7 月细纹狮子鱼占比最高,8 月太平洋褶柔鱼占比最高,9 月鳀占比最高,10—11月蓝圆鲹占比最高 (图2-b)。

图2 2021 年4—11 月黄海北部近岸海域各生物类群丰度(a, 尾)及生物量(b, kg)变化M1. 4 月,M2. 5—6 月,M3. 7 月,M4. 8 月,M5. 9 月,M6. 10—11 月。S1. 黄鲫,S2. 鳀,S3. 大泷六线鱼,S4. 蓝圆鲹,S5. 细纹狮子鱼,S6. 日本枪乌贼,S7. 太平洋褶柔鱼,S8. 脊腹褐虾,S9. 海葵(目),S10. 其他鱼种,S11. 其他无脊椎动物。Fig. 2 Abundance (a, ind) and biomass (b, kg) changes of various biota in offshore waters of the North Yellow Sea from April to November, 2021 M1. April, M2. May to June, M3. July, M4. August, M5. September, M6. October to November. S1. S. tenuifilis, S2. E. japonicus, S3. H. otakii, S4. D.maruadsi, S5. L. tanakae, S6. L. japonica, S7. T. pacificus, S8. C. affinis, S9. Actiniaria, S10. other fish species, S11. other invertebrates.
将4 至11 月调查周期内出现的渔业生物合并计算,IRI≥10 的渔业生物共计16 种,包含鱼类11 种,鳀、细纹狮子鱼、大泷六线鱼、黄鮟鱇、绒杜父鱼、蓝圆鲹、玉筋鱼(Ammodytes personatus)、黄鲫、长绵鳚(Zoarces elongatus)、大头鳕(Gadus macrocephalus)、方氏云鳚。甲壳类3 种,脊腹褐虾、戴氏赤虾、隆背黄道蟹。头足类2 种,日本枪乌贼、太平洋褶柔鱼。细纹狮子鱼、鳀、大泷六线鱼、日本枪乌贼和戴氏赤虾为4—11 月的共有种。
4 月主要种类包含16 种,其中优势种为黄鲫、日本枪乌贼和细纹狮子鱼3 种。5—6 月主要种类包含10 种,其中优势种为脊腹褐虾。7 月主要种类包含15 种,其中优势种为鳀、细纹狮子鱼和大泷六线鱼3 种。8 月主要种类包含16 种,其中优势种为鳀和太平洋褶柔鱼2 种。9 月主要物种包含20 种,其中优势种为鳀和日本枪乌贼2 种。10—11 月主要物种包含20 种,其中优势种为日本枪乌贼、蓝圆鲹和鳀3 种(表1)。

表1 2021 年4—11 月黄海北部近岸海域渔业资源主要物种相对重要性指数(IRI)Tab. 1 Index of relative importance (IRI) for the main species of fishery resources in offshore waters of the North Yellow Sea from April to November, 2021
4—11 月相对资源密度及底层温度、盐度分布如图3 所示。5—8 月调查所获渔业生物相对资源密度较高,9、10—11 月次之,4 月最低。其中5—6 月渔业生物高密度聚集区主要出现在獐子岛西南海域及海洋岛东部海域,7 月主要出现在海洋岛周边海域和鸭绿江口以南远岸海域,8 月主要出现在调查海域西南侧及海洋岛周边海域,其余月份均未形成明显聚集区。调查海域底层海水在4—9 月处于增温期间,直至10 月后才有所下降,4 月及10—11 月底层水温整体分布相对均匀,其余各月内基本皆呈现由近岸端向离岸端逐步降低的趋势(图3-a~f)。海域海水底层盐度月间变化幅度较小,5—7 月调查海域南部和东南部有高值区出现,整体分布离岸端低于近岸端(图3-g~l)。
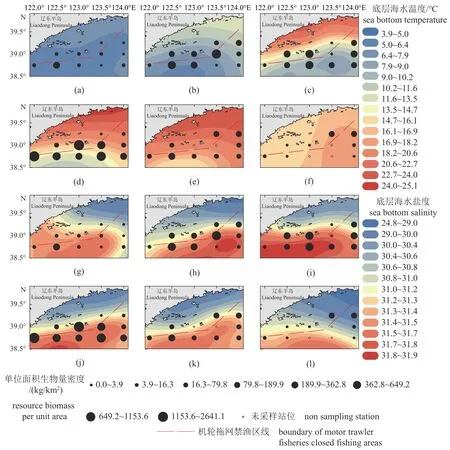
图3 2021 年4—11 月黄海北部近岸海域单位面积资源生物量及底层温度(a~f)和底层盐度(g~l)分布(a) 4 月,(b) 5—6 月,(c) 7 月,(d) 8 月,(e) 9 月,(f) 10—11 月,(g) 4 月,(h) 5—6 月,(i) 7 月,(j) 8 月,(k) 9 月,(l) 10—11 月。Fig. 3 Distribution of resource biomass per unit area with sea bottom temperature (a-f) and sea bottom salinity (g-l) in offshore waters of the North Yellow Sea, From April to November, 2021(a) April, (b) May to June, (c) July, (d) August, (e) September, (f) October to November, (g) April, (h) May to June, (i) July, (j) August, (k) September,(l) October to November.
2.2 黄海北部近岸海域鱼类亲、幼体种类及其季节变化
将4—11 月底拖网所获鱼类依据体长、体重和性腺发育期等生物学参数[37],初步判定产卵亲体共24 种,主要经济种类包括白姑鱼(Pennahia argentata)、小黄鱼、带鱼(Trichiurus japonicus)、镰鲳(Pampus echinogaster)、高眼鲽(Cleisthenes herzensteini)、圆斑星鲽(Verasper variegatus)、角木叶鲽(Pleuronichthys cornutus)、半滑舌鳎(Cynoglossus semilaevis)、大泷六线鱼、绿鳍鱼(Chelidonichthy kumu)和鲬(Platycephalus indicus)等。当年生(0 龄)幼体共29 种,主要经济种类包括油魣(Sphyraena pinguis)、蓝圆鲹、真鲷(Pagrus major)、带鱼、鲐(Scomber japonicus)、镰鲳、褐牙鲆(Paralichthys olivaceus)、高眼鲽、角木叶鲽、半滑舌鳎、绿鳍马面鲀(Thamnaconus modestus)、许氏平鲉(Sebastes schlegelii)、绒杜父鱼和大泷六线鱼等(表2)。其中产卵亲体种类5—6 月最多,出现15 种,其余依次为10—11 月出现5 种,4 月出现2 种,7 月出现3 种,8、9 月各出现2 种。整体看来产卵亲体种数占比在春季(4—6 月)最高(47.2%),在夏季(7—8 月)大幅减少(11.1%),而后于秋季(9—11 月)略有升高(15.4%)。0 龄幼体种类在5—6 月最多,出现14 种,其余依次为9月出现13 种,4 月出现9 种,10—11 月出现8 种,7 月出现7 种,8 月出现5 种。0 龄幼体种数占比随季节更替变化幅度较为平缓,其在春季为47.2%,夏季降低至33.35%,秋季回升至43.6%。此外,已达生物学最小型的非繁殖期成体在9 月最多,出现16 种,其余依次为7 月出现14 种,8月出现13 种,10—11 月出现12 种,4 月出现11 种,5—6 月出现9 种。未达生物学最小型的非0 龄鱼在9—11 月最多,皆出现6 种,其余依次为5—6 月出现5 种,8 月出现3 种,4 月和7月各出现2 种。

表2 4—11 月黄海北部近岸海域调查所获鱼类资源发育阶段Tab. 2 Development stages of fishery resources obtained from the investigation in offshore waters of the North Yellow Sea from April to November
2.3 黄海北部近岸海域群落多样性及ABC 曲线
2021 年4—11 月,黄海北部近岸海域渔业生物群落多样性有较为明显的月度变动(图4)。期间,Marglef 丰富度指数(D)基本呈现 “双峰” 分布,5—6 月和9 月群落D先升至顶峰与次高峰。Shannon-Wiener 多样性指数(H′)变化趋势与D基本一致,群落丰富度较高的月份H′也处于较高水平。Pielou 均匀度指数(J′)和Simpson 优势集中度指数(λ)整体变化幅度不大,二者呈现相对的变化趋势,峰值分别出现在9 和4 月,谷值分别出现在4 和9 月。

图4 2021 年4—11 月黄海北部近岸海域渔业生物群落多样性指数D. Marglef 丰富度指数,J′. Pielou 均匀度指数,H′. Shannon-Wiener多样性指数,λ. Simpson 优势集中度指数。Fig. 4 Diversity index of fishery biological community in offshore waters of the North Yellow Sea from April to November, 2021 D. Margalef richness index, J′. Pielou evenness index, H′. Shannon-Wiener diversity index, λ. Simpson dominance concentration index.
黄海北部近岸海域渔业生物群落丰度-生物量比较曲线如图5 所示。4—6 月,ABC 曲线中生物量优势度曲线与丰度优势度曲线相对位置变化明显,4 月丰度优势度曲线起点低于生物量优势度曲线,而后与之交叉,W为0.006。5—6 月丰度优势度曲线起点较4 月有所升高,并始终高于生物量优势度曲线,W为-0.095。7—9 月丰度优势度曲线皆高于生物量优势度曲线,W均为负值,10—11 月丰度优势度曲线起点大幅下降,优势不明显,与生物量优势度曲线极为接近,W为-0.05。

图5 2021 年4—11 月黄海北部近岸海域丰度-生物量比较曲线(a) 4 月,(b) 5—6 月,(c) 7 月,(d) 8 月,(e) 9 月,(f) 10—11 月。Fig. 5 Abundance biomass comparison curve in offshore waters of the North Yellow Sea from April to November(a) April, (b) May to June, (c) July, (d) August, (e) September, (f) October to November.
2.4 黄海北部近岸海域群落结构时空变化
通过计算季节间Jaccard 群落种类组成相似性指数和物种更替率(表3)可知,春(4—6 月)夏季(7—8 月)群落之间中等不相似,有28 种相同物种,物种更替较为明显;夏季秋季(9—11 月)群落之间中等相似,共有物种数达31 种,季节间物种更替率小于春夏季间更替率;春秋季间群落中等相似,物种更替率低于春夏季,与夏秋季相当,虽季节跨度较大,但仍有36 种相同物种出现。

表3 黄海北部近岸海域群落结构季节间物种相似性及更替率Tab. 3 Seasonal species similarity and replacement rate of community structure in offshore waters of the North Yellow Sea
春季黄海北部近岸海域渔业生物群落在50.6%的相似性水平上可分为3 组,其中鸭绿江口南部及海洋岛西南部(S1、S2、S6、S9、S11)为组1,獐子岛西南部及海洋岛东南部(S8、S10)为组2,海域西南端至獐子岛、海洋岛一带(S3、S4、S5、S7)为组3 (图6)。NMDS 分析结果与CLUSTER 结果一致,所得Stress 值小于0.1,表明分类结果较好[38]。ANOSIM 也验证春季各组间群落种类组成差异极显著(R=0.802,P<0.01)。后利用SIMPER 分析得出(表4,表5),组1 内各站位间的平均相似性(average similarity)为48.9%,组内典型种(相似性贡献大于4%)完全由甲壳类组成,主要为冷温性种,相似性累积贡献率达到91.9%。组2 内各站位间的平均相似性为9.7%,相似性累积贡献率达到79.5%,中上层暖水性和暖温性种类(黄鲫和日本枪乌贼等)为主要典型种(累积贡献率达33.2%)。组3 内各站位间的平均相似性为20.5%,相似性累积贡献率达到88.3%,玉筋鱼等底层冷温性种类为主要典型种(累积贡献率达58.1%)。组1 与组2、组3 的平均相异性(average dissimilarity)分别为98.4%和94.9%,导致组1 与其余两组群落产生差异的分歧种(相异性贡献大于4%)主要为底层种类(脊腹褐虾、戴氏赤虾和细纹狮子鱼等);组2 与组3 间的平均相异性为87.5%,分歧种主要为中上层(黄鲫和鳀)及底层鱼类(玉筋鱼)。

表4 黄海北部近岸海域各季节的典型种及其贡献率Tab. 4 Typical species and their contribution rate in each season in offshore waters of the North Yellow Sea
表5 黄海北部近岸海域各季节的分歧种及其贡献率Tab. 5 Discriminating species and their contribution rate in different seasons in offshore waters of the North Yellow Sea
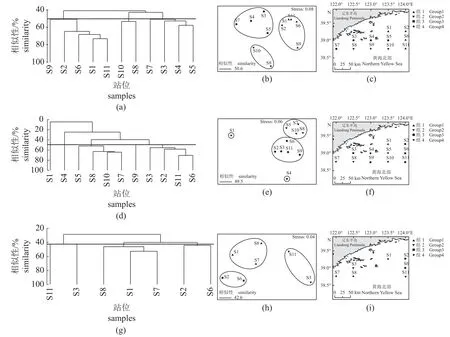
图6 春、夏、秋季黄海北部近岸海域渔业生物群落(a) (d) (g) 聚类图,(b) (e) (h) NMDS 排序图,(c) (f) (i) 站位示意图。(a) (b) (c) 春季,(d) (e) (f) 夏季,(g) (h) (i) 秋季。Fig. 6 Fishery biological community in offshore waters of the North Yellow Sea in spring, summer and autumn(a) (d) (g) cluster diagram, (b) (e) (h) NMDS ordination diagram, (c) (f) (i) station diagram. (a) (b) (c) spring, (d) (e) (f) summer, (g) (h) (i) autumn.
夏季群落在49.5%的相似性水平上可分为4 组,大洋河口周边海域(S1)为组1,獐子岛西南及海洋岛东南海域(S5、S7、S8、S10)为组2,鸭绿江口南部、獐子岛西部和东南部(S2、S3、S6、S9、S11)为组3,獐子岛与海洋岛相邻海域(S4)为组4 (图6)。NMDS 分析结果与CLUSTER 结果一致,所得Stress 值小于0.1,各组间群落种类组成差异极显著(R=0.872,P<0.01)。组1 内仅有蓝圆鲹一个种类(暖水性种);组2 平均相似性为57.7%,相似性累积贡献率达到88.9%,底层冷温性种类(大泷六线鱼和细纹狮子鱼等) 为主要典型种(累积贡献率达80.0%);组3 平均相似性为55.1%,相似性累积贡献率达到91.3%,中上层暖温性种类(鳀和太平洋褶柔鱼等)为主要典型种(累积贡献率达49.3%);组4 内仅有太平洋褶柔鱼(暖温性)和鲐(暖水性)两个种类。组1、组2 间的平均相异性为100%,分歧种为底层种类(隆背黄道蟹和大泷六线鱼等);组1、组3 间的平均相异性为91.0%,分歧种为中上层种类(鳀和太平洋褶柔鱼等);组1、组4 间的平均相异性为100%,分歧种为中上层种类(鳀和蓝圆鲹等);组2 与组3、组4 间的平均相异性分别为60.5%和86.4%,主要分歧种均为底层种类(脊腹褐虾和隆背黄道蟹等);组3、组4 间的平均相异性为66.6%,主要分歧种为中上层种类(鳀和鲐等)。
秋季部分站位因禁渔区、养殖区等问题未能作业,选取两月共现站位(S1、S2、S3、S6、S7、S8、S11)。秋季渔业生物群落在42.6%的相似性水平上可分为3 组,大洋河口南部及獐子岛西南部(S1、S7、S8) 为组1,调查海域东南端及獐子岛西部(S3、S11)为组2,鸭绿江口南部(S2、S6)为组3 (图6)。NMDS 分析结果与CLUSTER 结果一致,所得Stress 值小于0.1,各组间群落种类组成差异极显著(R=0.775,P<0.01)。各组内典型种均由暖温性和暖水性种类组成:组1 内各站间的平均相似性为48.5%,相似性累积贡献率达到79.3%,中上层种类(鳀和日本枪乌贼等)为主要典型种(累积贡献率达52.6%);组2 内平均相似性为42.6%,相似性累积贡献率达到100%,中上层种类(蓝圆鲹和鳀等)为主要典型种(累积贡献率达87.2%);组3 内平均相似性为44.4%,相似性累积贡献率达到94.5%,底层种类戴氏赤虾和三疣梭子蟹等为主要典型种(累积贡献率达58.0%)。组1 与组2、组3 群间的平均相异性分别为69.2%和59.8%,主要分歧种为中上层种类(鳀和日本枪乌贼等); 组2、组3 群间的平均相异性为74.4%,主要分歧种为中上层种类(日本枪乌贼和赤鼻棱鳀等)。
2.5 黄海北部近岸海域渔业生物群落与环境因子的关系
CCA 分析结果显示(图7),对于物种样本,第一、二排序轴间的相关系数仅为-0.04,表明两排序轴近乎垂直;对于环境因子,第一、二排序轴的相关系数为0,表明排序结果有效[39]。前两个排序轴的特征值分别为0.342 和0.313,共解释了物种群落变异程度的20.0%。44 个物种与环境因子排序轴(第一排序轴和第二排序轴)的相关系数都达到了0.86 以上,表明排序能够较好地反映物种与环境因子间的关系。物种编号见表6。

表6 黄海北部近岸海域CCA 分析所用物种编号Tab. 6 Species numbers used for CCA analysis in offshore waters of the North Yellow Sea
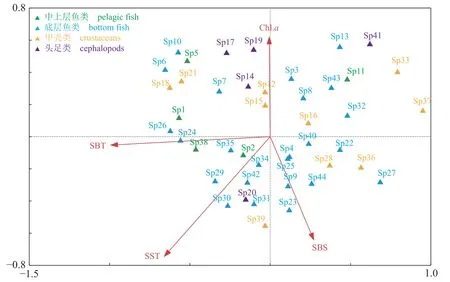
图7 黄海北部近岸海域主要生物类群丰度与主要环境因子间的CCA 排序图SBT.海水底层温度、SST.海水表层温度、SBS. 海水底层温度、Chl.a.叶绿素。Fig. 7 CCA ordinationof main biota and environmental factors in offshore waters of the North Yellow Sea SBT. sea bottom temperature, SST. sea surface temperature, SBS. sea bottom salinity, Chl.a. chlorophyll-a.
在所选环境因子中,SST、SBT、SBS、Chl.a是显著影响黄海北部近岸海域渔业资源结构时空变化的环境因子,其中水温条件(SST 和SBT)尤为重要。蓝圆鲹、鳀、太平洋褶柔鱼等物种的丰度与SST 成较强的正相关,双喙耳乌贼(S41)的适宜SST 最低,红鳍东方鲀(Sp30)的适宜SST 最高;细纹狮子鱼和方氏云鳚等物种丰度与SBT 成负相关,枯瘦突眼蟹(Sp37)的适宜底温最低,长蛇鲻(Sp6)的适宜底温最高;蓝圆鲹、鲐等物种丰度与Chl.a成正相关,隆背黄道蟹(Sp39)适宜叶绿素浓度最低,双喙耳乌贼(Sp41)适宜叶绿素浓度最高;大头鳕和隆背黄道蟹等物种丰度与SBS 成正相关,绿鳍马面鲀(Sp10)的适宜底盐最低,高眼鲽(Sp27)的适宜底盐最高。
3 讨论
3.1 黄海北部近岸海域渔业生物群落基本属性、特征与年代际演变
黄海北部近岸海域渔业生物群落并非作为独立群落存在,在黄海沿岸、辽南沿岸流和黄海暖流此消彼长的作用下,海区水温季节变化显著且盐度较低,游泳动物多为暖温种,暖水性种类所占比例远小于东海和南海海区[40],底栖动物多以沿岸广温低盐种为主,海域整体呈现暖温带区系特点,属北太平洋温带区东亚亚区[37]。
对比与本调查站位设置相似的早年(1998—2000 年)[3,8]和近年(2006—2007 年、2014—2017年)[5,11]调查,2021 年黄海北部近岸海域渔业生物群落优势种仍以底层、暖温性鱼类为主,种类总数亦变化不大,但不同水层的渔业生物群落结构存在明显变动。中上层鱼类在本次调查中渔获量占比较小(23.2%),总体优势地位有所降低。小型中上层鱼类生命周期短,对环境变化的响应高度敏感,1990 年以后对小型中上层鱼类的大规模开发利用[3]应为本次调查中上层渔获量显著下降的重要人为影响因素。在中上层鱼类群落中,斑鰶和青鳞小沙丁鱼(Sardinella zunas)不作为优势种类出现,鳀在近年来逐渐占据优势地位,但8 月盛期内资源存量(13 553.7 t)仍远低于1998—2000早期调查(33 515.9 t ,统一海域面积为4.73×104km2[8]),呈波动式下降趋势,各种群数量变动规律和差异仍需长时间的持续观测。蓝圆鲹为暖水种,在黄渤海区往年调查中甚少分布,夏季或为东海群系,成体曾偶见于黄海南部,秋季亦有幼鱼索饵[37],群体规模小且不稳定;2021 年8—11 月调查采集到一定规模的蓝圆鲹0 龄幼鱼,且于10—11 月出现大量集群现象,推测附近海域内有其育幼场存在,水文条件的年际变动及生物自身补充、摄食等特性的综合作用是育幼场形成的重要因素,然而,历史资料中少有蓝圆鲹在黄海北部集群的记录,因此无法确定该现象是否为网具选择性差异导致的偶然事件,其育幼场形成的主导因子以及该鱼种的利用潜力亦须开展深入研究。与历史调查相比[3,11],2021 年调查具有相近的网具规格及拖速,然而,作为历史优势鱼种的蓝点马鲛[3]及在2017 年资源量明显回升的鲐[11]在本次调查中却少有捕获,此2 种中上层重要经济鱼种在黄海北部的资源量呈波动式衰退迹象。底层种类中,脊腹褐虾、细纹狮子鱼和大泷六线鱼在黄海北部海域早年调查中占比较少,或受北黄海冷水团和海域渔业生物群落结构改变的影响,其虽为冷温性种类,但常于春夏季升温期至夏季高温期占据优势地位,成为季节性主要渔业生物。真鲷作为重要经济鱼种,其在历史调查中少有捕获,资源破坏严重[10],但随着人工养殖技术发展,增殖放流活动的逐步展开,加之伏季休渔期政策的充分保护,其在近年来复现于黄海北部近岸海域[11]。本次9 月调查于鸭绿江口以南海域捕获6 尾真鲷0 龄幼体,推测附近水域或存在其育幼场,真鲷资源呈恢复趋势。小黄鱼在20 世纪50—60 年代为本海区主要经济种类[41],其种群资源在高强度和无节制的捕捞后已遭受严重损害[1],20 世纪90年代开始实施的全面伏季休渔制度,缓解了部分捕捞压力,但小黄鱼种群的小型化、低龄化问题仍较显著[8-9,42],自2010 年始又有衰退趋势,后期难以形成规模渔汛[11]。小黄鱼种群在本次调查中的优势度亦较低,为各月海洋生物群落的少见种,种群资源呈现持续衰退迹象。对于头足类而言,太平洋褶柔鱼种群优势度于7 月开始升高,在8月调查中成为绝对优势种,推测该洄游群体或为东海产卵场发生的太平洋褶柔鱼分支[43]。黄海北部海域作为产卵育幼场,夏季鱼类稚幼体众多,饵料资源丰富[44],有利于其群体栖息生存。
3.2 黄海北部近岸海域渔业生物群落生物多样性与季节性演替
黄海北部近岸海域地处暖温带,在中纬度季风影响下气候季节变化明显,生境的季节变动在一定程度上影响了群落生物的季节性演替。各渔业生物类群为维持自身生命以及争夺繁衍后代所需饵料,会在产卵、越冬及索饵等各个时期依据自身生物学特性的需求,移动并占据一定的空间,以畅通其物质流动和能量流动[45]。由春季向夏季过渡时,黄海北部海域逐渐进入升温期,暖温性种类开始向海域内迁移,该海域作为重要产卵场也开始迎来产卵洄游盛期[46],因此春季末期(5—6 月)群落生物多样性及物种丰富度整体呈现上升趋势,鸭绿江口南部及海区中部海洋岛周边海域渔业生物群落典型种逐渐由冷温性底层鱼类和甲壳类演替为鳀和太平洋褶柔鱼等中上层暖温性种类。随着温度的不断升高,加之受北黄海冷水团边缘影响[47-49],獐子岛及海洋岛以南外部深水区群落逐渐演替为大泷六线鱼和细纹狮子鱼等底层冷温性种类,调查区域内冷温性种类个体的减少(如脊腹褐虾等)使得夏季群落生物多样性及物种丰富度大幅降低。秋季海域初入降温期,各种类产卵活动逐步进入末期,产卵后亲体及幼体在产卵场附近进行分散索饵,部分种类边索饵边逐渐向海域南部外海移动,在此期间,海域不同地理位置、不同水层的渔业生物群落基本更替为暖温性及暖水性种类,历经产卵盛期大量物种的迁入和幼体资源的补充,海域秋季群落丰富度和多样性都有明显提高。
综合来看,春夏季黄海北部近岸海域正处于渔业生物集中产卵洄游时期[46],生殖群体的大量迁入致使春-夏季物种更替幅度达全调查月份最高水平,而夏季海域冷水团边缘的低温水域为夏-秋季海洋环境变动提供了缓冲区,削弱了环境变化速率,为冷温性鱼种持续迁入及留存提供了适宜生境,进而缩小了夏-秋季物种更替幅度。
3.3 影响黄海北部近岸海域渔业生物群落稳定性的主要因素
本次调查自4 月中旬持续到11 月上旬,因渔业生物群落在不同程度上受到海洋环境、气候变化、种群迁入迁出以及人类活动的干扰,群落稳定性存在明显的月际变动。4 月黄海北部近岸海域正值春季,尚未进入大幅升温期,气候条件相对稳定,渔业生物群落中r选择种类丰度优势度有增加趋势,人为捕捞与生境变化对群落的干扰处于中度水平[50-52],群落相对稳定。5—6 月獐子岛和海洋岛南部海域出现大规模海葵暴发现象,泛滥繁殖的海葵极大地压缩了底栖生物的空间生态位,其复杂的食性也限制着其他渔业生物的生存繁衍[53],为北黄海底栖生态系统带来巨大压力,因此,黄海北部近岸海域渔业生物群落在5—6 月受到较强的自然扰动,群落处于极不稳定状态。春夏季为产卵洄游盛期[46],大量渔业生物的产卵洄游行为形成了较为持续的种群迁入扰动,加之辽南沿岸水系的增强,海域生境剧烈变化,7 月群落所受干扰进一步加剧。8 月海区内大量分布的水母(单次网获最高可达2 000 kg)极大地占据了海洋上层空间生态位,并使得海域生境遭受一定程度破坏,生物及非生物环境扰动大幅增加,加之产卵洄游种类的不断迁入扰动,群落处于严重干扰状态,不过,诸多洄游种类的聚集形成了更为复杂的营养通道[3],使得群落具备较强的抗干扰能力,且由于伏季休渔期内人为捕捞扰动大幅降低,有效缓冲了海域生境干扰,8 月群落整体稳定性较7 月有所提高。9—11 月,随着休渔期的结束,人为捕捞扰动骤增,群落结构遭受严重干扰,其稳定性进一步降低。综合看来,伏季休渔制度的设立可为海域渔业生物群落结构稳定性提供良好保障,但黄海北部近岸海域渔业生物群落稳定性整体仍处于较低水平。
(作者声明本文无实际或潜在的利益冲突)
