6种铁线莲种质SSR遗传多样性分析
2024-05-13李勇慧
李勇慧
(洛阳师范学院 生命科学学院,河南 洛阳 471934)
铁线莲是毛茛科(Ranunculaceae)铁线莲属(ClematisL.)植物,是一种具有多种抗逆性的藤本植物,且观赏价值很高,享有“藤本花卉皇后之称”[1].它不仅具有园艺学价值,可做展览用切花或用作地被植物[2],同时该属植物的药用价值较高,具有抗炎、镇痛、通淋利尿等作用[3-4].全球已知的铁线莲属植物有350余种[5],国内该属植物超过140种,主要分布在华中和西南地区,尤其云南最多[6-7].调查显示,河南有35个类群的铁线莲(24个种,1个亚种,10个变种)[2].
随着分子生物学技术的快速发展,利用分子标记技术研究作物遗传多样性及其亲缘关系分析的报道也日渐增多.通过不同个体间核苷酸数有所区别的原理准确判断生物的种内、种间差异,是DNA分子标记技术的显著特点,且它与环境和发育阶段无关,多态性较强,多表现为共显性[8].其中,简单重复序列(SSR)标记具有数量丰富、多态性好、信息量高、准确度高等优点[9],该技术是一种用来构建遗传图谱、进行品种鉴定、谱系分析和遗传多样性分析的有效手段.
SSR技术已应用于研究多种植物的遗传结构和遗传多样性[10-11].熊阳阳等构建了部分铁线莲的cDNA文库,对聚类后得到的Unigene进行功能注释和同源搜索,查找分析了SSR位点,为铁线莲‘薇安’的不同花被类型的差异基因表达、筛选和探究重被花形成的分子机制提供了科学依据[12].陈文超等研究了铁线莲属其他野生品种和栽培品种中11对叶绿体SSR引物的通用性,据此进行cpSSR引物开发,还可以用于检测铁线莲野生种和栽培种之间的遗传信息,筛选出的cpSSR引物可作为铁线莲属植物系统发育、群体遗传结构等遗传信息检测和研究的有效工具[13].目前尚未有利用SSR技术对不同种质或不同居群的铁线莲进行遗传多样性分析的研究.因此,本试验通过SSR标记技术,利用6对引物对25个居群的120个铁线莲种质材料进行遗传多样性分析,确定不同材料间的遗传背景差异.
1 材料与方法
1.1 试验材料
本试验选取的材料共25个居群120个样品,其中包括陕西铁线莲(Clematisshensiensis)、大叶铁线莲(Clematisheracleifolia)、钝萼铁线莲(Clematispeterae)、尾叶铁线莲(Clematisurophylla)、太行铁线莲(Clematiskirilowii)、粗齿铁线莲(Clematisargentilucida)6种种质资源,具体如表1所示.
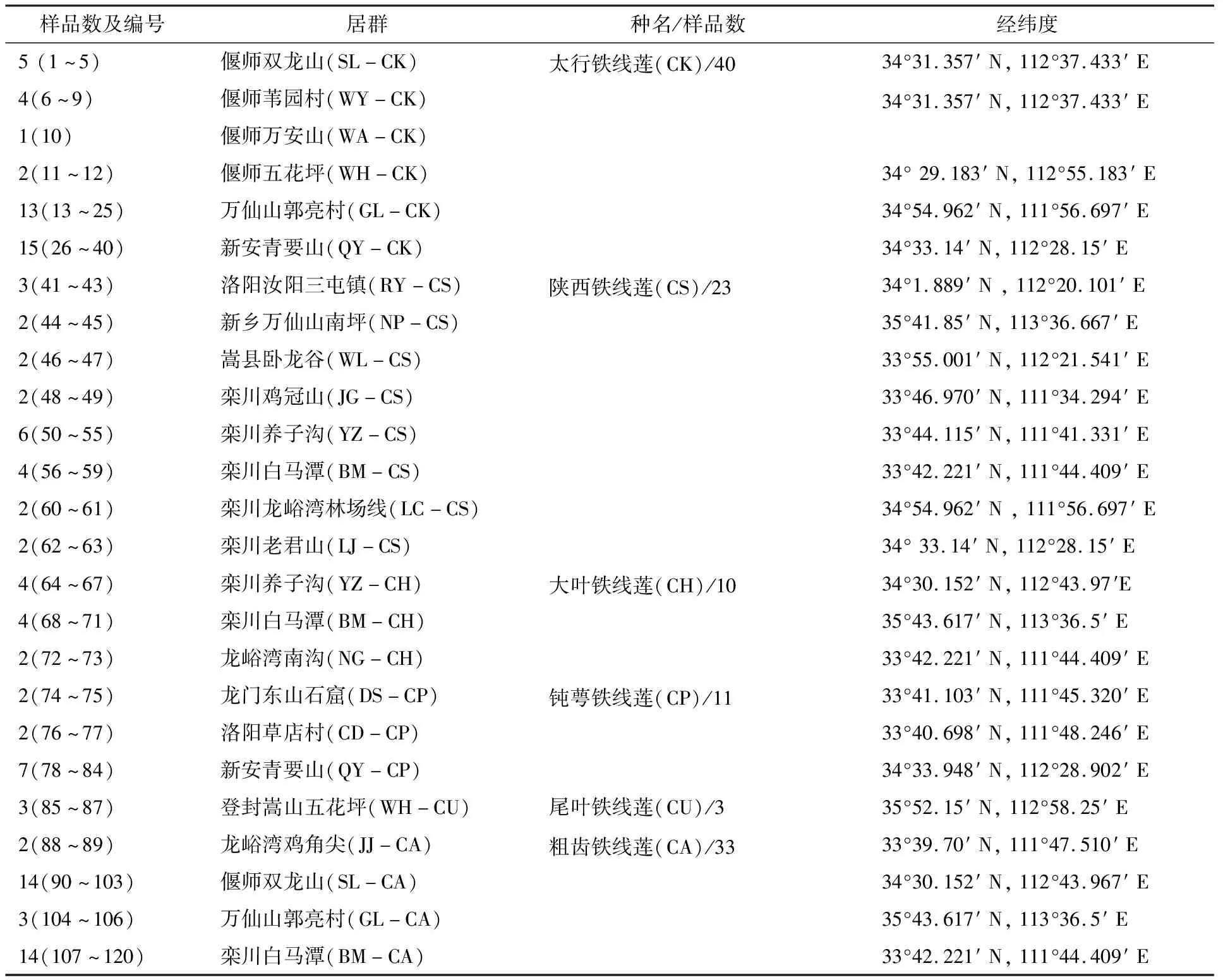
表1 样品及编号
1.2 主要试剂
试验所用的主要试剂:3 mol/L NaNC溶液,75%乙醇溶液,1×TBE缓冲液,2% CTAB溶液,10%聚丙烯酰胺凝胶母液,TBE,AgNO3,NaOH,甲醛.
1.3 试验方法
1.3.1 DNA的提取及结果检测
DNA的提取采用改良的CTAB法[14].用1%的琼脂糖凝胶电泳检测DNA的提取质量,通过超微量分光光度计(凯奥K5500 Plus,北京凯奥科技发展有限公司)进行测定,从中选出浓度与纯度较好(浓度在50 ng/mL左右)的DNA,放入冰箱备用.
1.3.2 PCR扩增
取出提取成功的120个DNA样品,从100对随机引物中筛选出多态性较好的6对引物进行PCR扩增,扩增完成后于4 ℃冰箱中保存.PCR扩增体系为:正反引物各0.5 μL,ddH2O 3.5 μL,2×pcr mixture 5 μL,DNA模板0.5 μL,总体系共计10.0 μL.铁线莲属SSR扩增程序如表2所示.由于不同引物的Tm值有所不同,退火温度也有差值,所以需要分批次进行PCR扩增.

表2 铁线莲属SSR扩增程序
1.3.3 聚丙烯酰胺凝胶电泳
将两个干净的玻璃板并齐重叠,一个玻璃板带耳朵,另一个带缝隙,将带缝隙的一面朝里面用夹子固定.按TBE∶琼脂粉=100 mL∶1 g的配比配制琼脂糖凝胶溶液,加热溶解后倒入凝胶槽,放固定好的玻璃板,凝固30 min.然后用玻璃棒将10%的聚丙烯酰胺凝胶工作液引流到玻璃板之间的缝隙,直到没过,插入梳子,凝固40 min.将玻璃板放入电泳槽中,带耳朵一侧朝里面,取下梳子,点样.电泳条件:恒压电压300 V,电流100 mA,3 h.
1.3.4 银染检测
取出玻璃板,将胶完整揭下,清水洗三遍.然后用250 mL 1%的硝酸银溶液染色10 min,水洗三遍至水清澈无白色浑浊.最后用250 mL显影液(0.2 g AgNO3,1 g NaOH,2 mL甲醛,纯水500 mL)在摇床上显色10 min.显示出条带后,放在胶片观察灯上观察,并拍照留档.
1.3.5 数据统计与分析
2 结果
2.1 多态性和遗传多样性分析
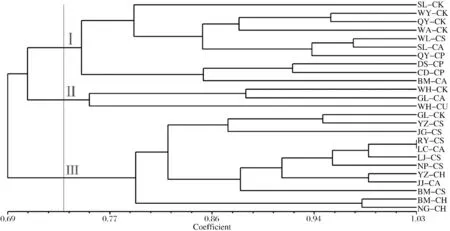
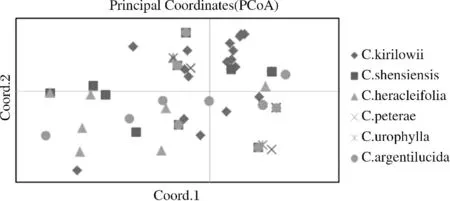
用筛选过的6对引物对25个居群的120个铁线莲样本进行SSR标记扩增,部分扩增图如图1所示.6对多态性引物在120个铁线莲中检测到33个等位基因,平均每对引物检测出5.5个等位基因.用POPGENE1.32软件计算出的平均等位基因数为2.00,平均有效等位基因数为1.577±0.233,Shan-non信息指数为0.527±0.136,Nei’s基因多样性指数为0.350±0.113.各引物的PIC值变化范围在 0.619到0.738之间,平均PIC值为0.638.PIC值在0到1之间,越接近1则多态性越好,反之越差.PIC>0.500时,是高度多态位点,PIC<0.250时,是低度多态位点,0.25 图1 SSR扩增图谱 用ACDsee软件对拍照留档的照片进行格式转换,再用Quantity one软件对多态性条带进行统计,整理出Excel里的0-1矩阵.用NTSYSpc-2.10e软件进行分析后得到120个铁线莲样品的UPGMA聚类图,120个样品分属于25个居群(见表1),聚类关系树状图如图2所示.遗传相似系数在0.58~1.00之间.遗传相似系数可以作为分类的依据,在遗传系数0.68处可以分为三个大类,再以0.77为界又可以将第Ⅰ大类继续分为四个亚类.Ⅰ-Ⅰ类包括太行铁线莲、陕西铁线莲和大叶铁线莲里的偃师双龙山(SL-CK)、偃师苇园村(WY-CK)、新乡万仙山郭亮村(GL-CK)、新安青要山(QY-CK)、洛阳栾川鸡冠山(JG-CS)、洛阳栾川白马潭(BM-CS)、洛阳龙峪湾南沟(NG-CH)等7个居群.Ⅰ-Ⅱ类包括太行铁线莲、陕西铁线莲、钝萼铁线莲、大叶铁线莲、粗齿铁线莲里的偃师苇园村(WY-CK)、万仙山郭亮村(GL-CK)、新安青要山(QY-CK和QY-CP)、嵩县卧龙谷(WL-CS)、栾川白马潭 (BM-CH和BM-CA)、龙门东山石窟(DS-CP)、偃师双龙山 (SL-CA)等 9个居群.Ⅰ-Ⅲ类包括太行铁线莲、粗齿铁线莲里的偃师双龙山(SL-CK)、栾川白马潭(BM-CA)两个居群.Ⅰ-Ⅳ类包括太行铁线莲、尾叶铁线莲、粗齿铁线莲和大叶铁线莲里的登封嵩山五花坪(WH-CU)、万仙山郭亮村(GL-CK和GL-CA)、栾川养子沟(YZ-CH)、栾川白马潭(BM-CH)等5个居群.同时,以0.78为界,第Ⅲ大类分为三个亚类,主要是陕西铁线莲和大叶铁线莲以及仅有的两个粗齿铁线莲. 图2 120个铁线莲样本的UPGMA聚类图 用POPGENE1.32软件计算出25个居群的铁线莲的遗传相似系数,其范围在0.59~1.03之间.根据遗传相似系数,用NTSYSpc-2.10e软件得到25个居群的UPGMA亲缘关系聚类图(见图3).在遗传系数0.74处分成三个大类,第一大类主要包括太行铁线莲里的偃师双龙山(SL-CK)、偃师苇园村(WY-CK)、新安青要山(QY-CK)、偃师万安山(WA-CK),钝萼铁线莲里的新安青要山(QY-CP)、龙门东山石窟(DS-CP)、洛阳草店村(CD-CP),陕西铁线莲里的嵩县卧龙谷(WL-CS),还有粗齿铁线莲里的偃师双龙山(SL-CA)和栾川白马潭(BM-CA).第二大类主要包括太行铁线莲里的五花坪(WH-CK),粗齿铁线莲里的新乡万仙山郭亮村(GL-CA),尾叶铁线莲里的登封嵩山五花坪(WH-CU).第三大类主要包括太行铁线莲里的新乡万仙山郭亮村(GL-CK)和距离较远的陕西铁线莲的其余居群,以及所有的大叶铁线莲和粗齿铁线莲里的龙峪湾鸡脚尖(JJ-CA). 图3 25个铁线莲居群的UPGMA聚类图 利用0-1矩阵通过GenAlex6.5软件进行铁线莲样品的PCoA分析(如图4所示).由图4可知,太行铁线莲主要集中在右上部,陕西、粗齿和大叶铁线莲大多分布在左下部,尾叶和钝萼铁线莲主要分布在左上和右下部.坐标(coordinate)cood.1和cood.2分别解释整体遗传变异的26.21%和46.02%. 图4 6个种质铁线莲的PCoA分析图 本研究中120个铁线莲的PIC值在0.619到0.738之间,其中96号引物的PIC值最高,为0.738,说明这对引物的扩增效率比较强.6对引物共扩增出33条带,其中96号引物扩增出的条带最多,为9条,15号只有2条,平均每个扩增出5.5条.在这6对引物中,3对含有二核苷酸重复序列,2对含有四核苷酸重复序列,1对含有六核苷酸重复序列,表明本研究所用的25个居群的铁线莲的多态性都比较好,SSR比较适合于铁线莲的遗传多样性分析. 根据NTSYSpc-2.10e软件得到120个个体的聚类图(见图2).由图2可知,在大多数情况下,以遗传相似系数作为分析的依据,同一种质的铁线莲会优先聚集在一个大类中,继而再按照地理位置进行聚类.譬如在第Ⅰ-Ⅰ大类中多是太行铁线莲,其中地理位置较近的偃师双龙山和偃师苇园村会优先聚集在一起.第Ⅲ大类中洛阳栾川养子沟的陕西铁线莲和同在栾川养子沟的大叶铁线莲优先聚集到一起.这也表明所处位置的地理差异对种群之间的遗传作用有一定影响.林立等及张杰等的研究也证实了它们的关联,提出两者之间是呈正相关的[17-18]. 用POPGENE1.32软件计算出的25个铁线莲居群遗传相似系数在0.59~1.03之间,平均值为0.77.遗传相似系数最小的是钝萼铁线莲的洛阳草店村居群和陕西铁线莲的新乡万仙山南坪居群,这说明这两个居群亲缘关系较远.遗传相似系数最大的为陕西铁线莲的洛阳汝阳三屯镇和栾川龙峪湾林场线居群,表明这两个居群的亲缘关系较近. 通过聚类图(见图3)可知,大多情况下,同一种的铁线莲会优先聚到一大类中,然后同一种内的铁线莲基本会按照地理位置的远近进行聚类,如第一类中的太行铁线莲先是距离较近的偃师双龙山、偃师苇园村、新安青要山、偃师万安山四个居群聚在一起,然后距离较远的万仙山郭亮村单独聚在第三类中.陕西铁线莲在第三类中也是按距离的远近进行聚类.其他种质也大体上能按距离远近进行聚类,表明种群间的遗传距离与地理距离有一定的关系.但万仙山郭亮村的太行铁线莲居群并没有跟太行铁线莲的其他居群聚类到一起,而是跟陕西铁线莲的居群聚类到一起,白马潭的陕西铁线莲也没有和其他陕西铁线莲聚在一起,其原因可能是距离较远或发生了基因交流.Fischer 等的研究也表明了种群内部的基因流会影响居群的遗传结构[19]. 利用GenAlex6.5软件进行PCoA分析(见图4),其结果与UPGMA个体与居群的聚类结果基本一致.Zhou等对Tapisciasinensis进行SSR分析,其PCoA分析结果与UPGMA聚类结果一致[20].可以看到,PCoA图中太行铁线莲主要集中在右上部,与居群第一大类大部分一致,陕西、粗齿和大叶铁线莲大多分布在左下部,与居群第三大类接近.尾叶和钝萼铁线莲主要分布在左上和右下部.这些都说明相同种质的铁线莲基本上能聚在一起. 本研究利用6对引物对25个居群的120个铁线莲进行基于SSR标记的遗传多样性分析研究.研究表明,用筛选出来的6对引物对120份铁线莲进行的SSR-PCR扩增,共得到33个等位基因,平均每对引物扩增的等位基因数为5.5个,PIC值均大于0.5,说明河南的这6种铁线莲具有较高的遗传多样性.相同种质的铁线莲优先聚在一起,继而再按照地理位置进行聚类.研究结果可为后续铁线莲种质创新和新品种选育提供参考.
2.2 铁线莲个体遗传关系聚类分析

2.3 铁线莲居群遗传关系聚类分析
2.4 铁线莲种质的主成分分析
3 讨论
4 结论
