不同授粉品种花粉对湘猕枣果实生长和品质的影响
2024-04-30黄晗羽莫沙陈梦洁唐育辉王仁才罗飞雄
黄晗羽 莫沙 陈梦洁 唐育辉 王仁才 罗飞雄


摘 要:【目的】研究授粉品種对软枣猕猴桃湘猕枣坐果和果实品质的影响,筛选出湘猕枣的最佳授粉品种。【方法】选用软枣猕猴桃雄株DWSR1、NER1、R2px2的花粉、美味猕猴桃花粉和长叶猕猴桃雄株AASAH1花粉为湘猕枣授粉,研究授粉处理对湘猕枣不同时期子房内源激素含量、坐果率及果实品质的影响。【结果】AASAH1授粉处理落果严重,坐果率最低(9.68%~33.33%);其他授粉处理的坐果率均在40%以上,其中NER1和DWSR1授粉处理坐果率在70%以上。2023年,NER1和DWSR1授粉处理的果实单果质量、果形指数、干物质和可溶性固形物含量显著高于其他授粉处理,且在贮藏期硬度下降速度和呼吸速率更慢。授粉后75 h,AASAH1授粉处理的子房中GA3含量、IAA含量、IAA/ABA、GA3/ABA和(GA3+IAA)/ABA均处于较低水平,而ABA含量较高。【结论】湘猕枣种内授粉亲和性好于种间授粉,NER1和DWSR1可配置为湘猕枣适宜授粉雄株。
关键词:猕猴桃;授粉;果实品质;内源激素
中图分类号:S663.4 文献标志码:A 文章编号:1009-9980(2024)04-0725-13
Effects of pollinizer cultivars on berry growth and quality of Xiangmizao kiwifruit
HUANG Hanyu1, MO Sha1, CHEN Mengjie1, TANG Yuhui2, WANG Rencai1*, LUO Feixiong1*
(1College of Horticulture, Hunan Agricultural University, Changsha 410128, Hunan, China; 2Yulin Fruit Production Professional Cooperative of Liuyang City, Liuyang 410308, Hunan, China)
Abstract: 【Objective】 Kiwifruit, a dioecious plant, must be effectively pollinated to obtain fruit quality and optimal yield because their complete flowers produce unviable pollens and their male flowers have abortive pistils. Besides, pollinizer cultivars confer “xenia” effect on the fruit, which affects a range of important traits including fruit set, berry size, berry shape, color, developmental timing, metabolites composition, etc. In kiwifruit vineyard, the ratio of male and female plants is generally recommended as 1∶8. Compatible male cultivars for Actinidia arguta production are lacking and more studies are needed to select compatible pollinizer cultivars for any specific kiwiberry female cultivars. For selecting a compatible male cultivar, its flowering time, amount of pollen in each flower and pollen viability need to be considered. There have been few studies investigating the effects of pollen parents on berry quality in kiwiberry. Xiangmizao (A. arguta) is a new kiwiberry cultivar, which was selected from the wild kiwifruit population growing in Dawei Mountain, Hunan Province. To grow this new cultivar properly, compatible pollinizer cultivars are needed. To select the best pollinating male cultivars compatible for Xiangmizao, four pollinizer cultivars were used to investigate pollen parent effect on fruit set, berry growth, berry quality, and berry storability of this new female cultivar. 【Methods】 Pollens from a tetraploid A. arguta male named DWSR1 collected from Dawei mountain (T1-22/23), a tetraploid A. arguta male named NER1 collected from Northeast China (T2-22/23), a tetraploid A. arguta male named R2px2 (T2-23), a tetraploid and a hexaploid A. deliciosa males (T3Ad-22) and a diploid A. hemsleyana male named AASAH1 (T4-23) were used to pollinate the female cultivar Xiangmizao. This study compared the differences in endogenous hormones in the ovaries at different developmental stages. Ovary samples were collected at different time points after pollination. Fruit set, berry growth, berry quality, and berry storability were recorded for the pollinated cultivar. 【Results】 T4-23 showed serious fruit drop after pollination and had the lowest fruit set rate in the end. Compared to T4-23, the other treatments had higher fruit set above 40%, with T1-22/23 and T2-22/23 having fruit set above 70%. Seventy-five hours after pollination, endogenous hormones and their ratios such as GA3, IAA, IAA/ABA, GA3/ABA and (GA3+IAA)/ABA were all at relatively lower levels in the ovaries of T4-23, while its ABA content was the highest, compared to the other treatments. In contrast to T4-23, except for ABA content, endogenous hormones and their ratios in ovaries collected from T1-23 and T2-23 were higher than those from the other two treatments. In 2022, average berry weight of T1-22 (12.18 g) and T3Ad-22 (11.78 g) was significantly higher than that of T2-22 (10.26 g). Fruit shape index was above 1.2 in all treatments, with T1-22 having significantly higher fruit shape index (1.40) than T2-22 (1.30) and T3Ad-22 (1.28). Fruit firmness of berries from T1-22 (43.21 N) was significantly higher than from the other treatments. T1-22 also had a higher titratable acid content (1.13 %), which was 79.37% higher than T3Ad-22, which had the lowest titratable acid (TA) content. Berry firmness of T3Ad-22 declined more slowly than the other two treatments, T1-22 and T2-22, and their total soluble solids (TSS) were consistently lower than that of T1-22. In 2023, average berry weight of both T1-23 and T2-23 was significantly higher than that in T3Aa-23, with T2-23 being 2.37 times higher than T3Aa-23. There was no significant difference in berry shape index between the T1-23 and T2-23, which both reached 1.4 and significantly higher than that of T3Aa-23 (1.16). Fruit dry matter content was higher than 20% in all treatments at harvest, with T3Aa-23 (21.03%) being the lowest. TSS contents of T1-23 (7.63%) and T2-23 (7.88%) were higher than that of T3Aa-23 (6.64%). Berry TA content of T1-23 (1.10%) was significantly (1.46 times) higher than that of T2-23 (0.75%) and also significantly (1.45 times) higher than that of T3Aa-23 (0.76%). Fruit firmness declined more rapidly in T1-23 and T2-23 than in T3Aa-23. Berry respiratory rate of T3Aa-23 was consistently higher than that of T1-23 and T2-23. 【Conclusion】Results from this study suggest both DWSR1 and NER1 could be used as suitable pollinizers for Xiangmizao fruit production.
Key words: Actinidia arguta; Pollination; Fruit quality; Endogenous hormones
猕猴桃属(Actinidia)在全世界范围内有66种,在中国有62种且自然分布广泛[1]。软枣猕猴桃(A. arguta)是所有猕猴桃中分布最广的种之一,在中国、俄罗斯远东区和日本均有发现[2]。猕猴桃为雌雄异株植物,雌株的花虽是完全花,但其花粉不具生活力;雄株的花雌蕊败育,故需要配置授粉树或进行人工授粉[3]。目前在生产栽培时,一般采用雌雄树比例为8∶1的形式配置授粉树,软枣猕猴桃配套的雄性品种混乱,选择雄性品种时大多只考虑其花期是否相宜,而忽视其对果实品质的影响。
花粉来源对果实品质产生影响的现象广泛存在于水果(苹果[4]、葡萄[5]、柑橘[6]、蓝莓[7]、梨[8])、蔬菜(黄瓜[9]、番茄[10])等农作物中。陈庆红等[11]首次在金魁猕猴桃上发现不同花粉授粉后,金魁的果形、单果质量、可溶性固形物含量、维生素C含量、贮藏期等均有变化,后续诸多研究也显示中华猕猴桃[12]、美味猕猴桃[13]、软枣猕猴桃[14]等都存在此类现象。
植物内源激素在植物生长发育过程中起重要作用,且不同激素之间也会相互作用,以相互促进或相互拮抗的方式對植物的生长发育过程进行调控[15-16]。针对猕猴桃的内源激素检测方面的研究,目前对花蕾[17]、芽[18]、枝条[19]和果实[20-21]进行了相关研究,而对授粉后子房内源激素的研究鲜少报道。
软枣猕猴桃新品种湘猕枣是从湖南省浏阳市大围山野生软枣猕猴桃群体中选育而成的,果实风味独特,抗逆性强[22]。目前实际生产中面临的主要问题是缺少适宜的授粉树,导致产量低,果实品质良莠不齐。笔者通过使用不同花粉对湘猕枣进行授粉试验,以探究其对坐果和果实品质的影响,筛选出适宜的授粉品种以提高产量与品质,以期为生产上授粉树的配置提供理论依据。
1 材料和方法
1.1 材料
试验于2022年5月—2023年5月在湖南省浏阳市大围山进行,供试雌性品种为软枣猕猴桃新品种湘猕枣,均为10年生树,砧木为美味猕猴桃实生苗。授粉雄株包括软枣猕猴桃NER1、DWSR1、AASAH1、美味猕猴桃(来源于商业花粉)和软枣猕猴桃R2px2(来源于中国农业科学院郑州果树研究所)。分别于授粉后20、30、45、55、75、100和120 h采集子房并迅速进行液氮处理,存于-80 ℃以备内源激素含量测定。
1.2 方法
1.2.1 试验处理 各授粉处理选择3株树势一致的湘猕枣,在大蕾期将树体不同方位的结果枝上进行套袋处理并编号,授粉一周后解开套袋,每种花粉授粉花朵数均在300朵左右。授粉方式分别采用人工点授(2022年)和液体授粉(2023年),花粉液为10%蔗糖+0.1 g·L-1硼酸+0.2 g·L-1羧甲基纤维素钠+1.0 g·L-1花粉+纯水[23],授粉后立即套袋,记录授粉花朵数,并于授粉后15和30 d时统计各处理坐果率。
1.2.2 子房内源激素含量测定 采用高效液相色谱法测定吲哚乙酸(IAA)、赤霉素(GA3)和脱落酸(ABA)含量[24]。将样品于液氮中研磨至粉碎,准确称量适量试样置于试管中,加入10倍体积乙腈溶液,并加入4 μL内标母液;4 ℃提取过夜,12 000 r·min-1离心5 min,取上清液;沉淀再次加入5倍体积乙腈溶液,提取两次,合并所得上清液;加入200 mg C18填料,剧烈震荡30 s,10 000 r·min-1离心5 min,取上清液;浓缩至干,以200 μL甲醇复溶,过0.22 μm有机相滤膜,放入-20 ℃冰箱待上机检测。色谱条件为色谱柱WATERS ACQUITY UPLC HSS T3(2.1 mm×100 mm,1.8 ?m),柱温为30 ℃,使用的流动相A为0.1%甲酸水,流动相B为乙腈,流速设置为0.3 mL·min-1,进样量为2 μL。
1.2.3 果实品质指标测定 授粉后14 d,每隔7 d追踪测定12个果实的生长情况,用游标卡尺[MNT-150,上海美耐特实业(集团)有限公司,中国]测量果实纵径和横径。果实采收后立即送至实验室内,分别置于25 ℃(2022年)和4 ℃(2023年)中贮藏,定期测定果实品质。果实质量采用电子天平(XY500C,常州市幸运电子设备有限公司,中国)测定;果实硬度采用数显推拉力计(ZP-50,香港艾固仪器仪表有限公司,中国)测定;果实可溶性固形物(TSS)含量使用数显糖酸一体机(PAL-BX∣ACID8,Atago,日本)测定;干物质含量参考刘磊等[25]的方法测定;可滴定酸(TA)含量参考张艳霞等[26]采用的酸碱滴定法测定;维生素C含量参考田彦龙等[27]采用的2,6-二氯靛酚法测定。
1.2.4 数据统计分析 采用Excel 2016进行数据整理及统计,采用SPSS 25进行差异显著性分析,采用Origin 2022作图。
2 结果与分析
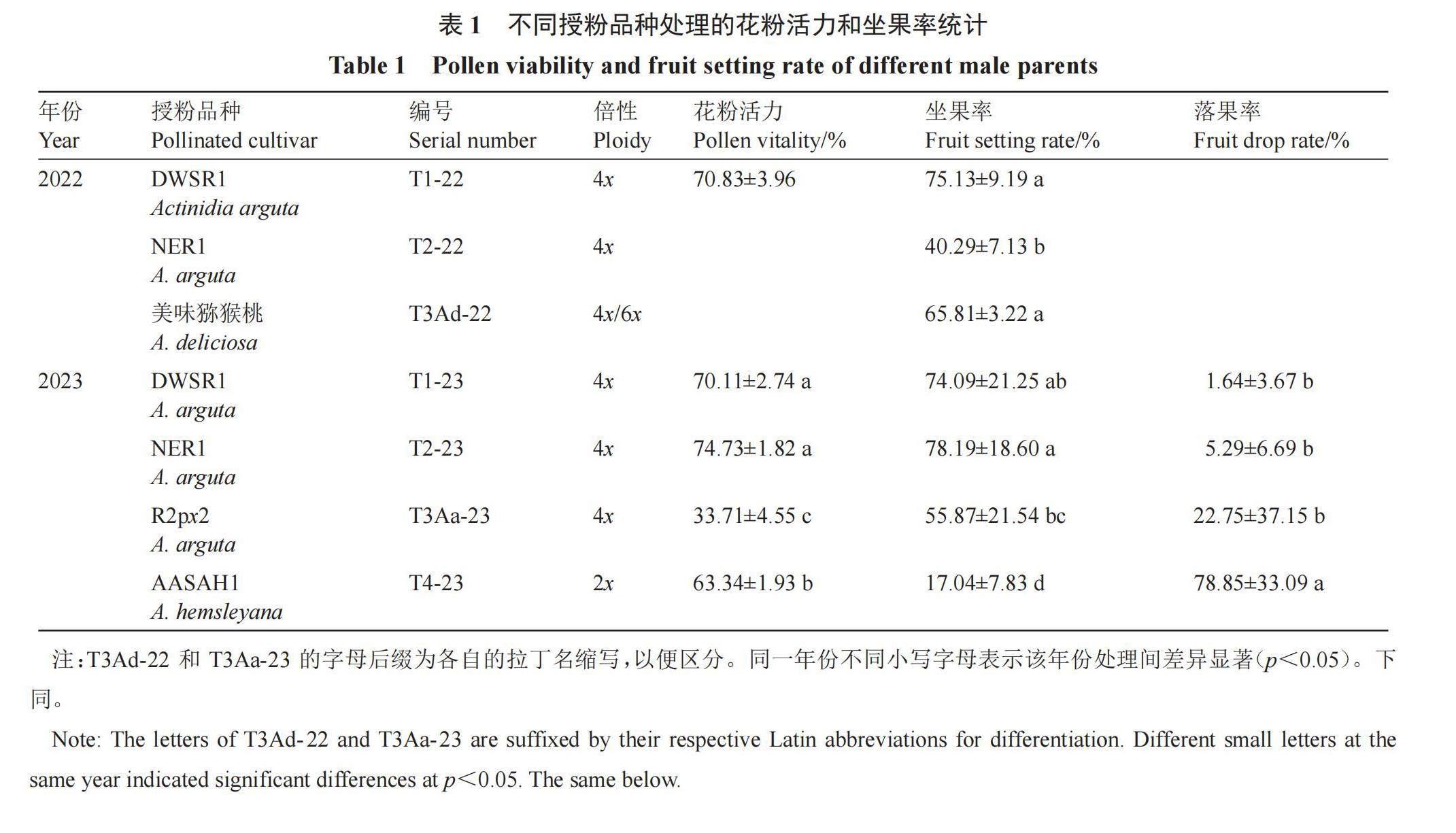
2.1 不同授粉品种处理的花粉活力和坐果率统计
2022年T1-22和T3Ad-22处理的坐果率分别比T2-22显著提高34.84和25.52个百分点。2023年T1-23和T2-23的花粉活力显著高于其他处理,且均在70%以上,其中T2-23的花粉活力相对更高,分别是T3Aa-23和T4-23的2.22倍和1.18倍;T1-23和T2-23间的坐果率差异不显著,且均在70%以上,显著高于T4-23,分别是T4-23处理的4.35倍和4.59倍;T3Aa-23的坐果率显著高于T4-23,前者是后者的3.28倍;T4-23处理的植株落果率显著高于其他处理,为78.85%,T1-23处理的落果率最低,仅有1.64%,前者落果率是后者的48.08倍(表1)。
2.2 不同授粉品种处理对湘猕枣授粉后子房内源激素含量的影响
授粉后30 h,T1-23和T4-23处理的子房GA3含量分别是授粉前的19.18倍和2.17倍,而T2-23和T3Aa-23处理的子房GA3含量分别下降了94.30%和78.51%。授粉后75 h,T1-23处理的子房GA3含量最高,是含量最低处理T4-23的10倍。授粉后120 h,T3Aa-23和T1-23处理的子房GA3含量均有不同程度的上升,此时各处理的子房GA3含量从高到低排序为T1-23>T3Aa-23>T2-23>T4-23,其中T1-23是T4-23的8.82倍(图1-A)。
授粉后30 h,T4-23处理的子房IAA含量较授粉前上升了54.26%,而T2-23、T3Aa-23和T1-23处理的子房IAA含量分别下降了62.55%、31.70%和54.35%。授粉后75 h,T2-23处理的子房IAA含量最高,分别是T1-23、T4-23和T3Aa-23处理的1.52倍、3.22倍和8.61倍。授粉后120 h,各处理的子房IAA含量从高到低排序为T2-23>T4-23>T3Aa-23>T1-23,其中T2-23是T1-23的2.42倍(图1-B)。
授粉后30 h,各处理的子房ABA含量均大幅下降,T1-23处理的子房ABA含量最高,是含量最低处理T4-23的5.31倍;此后T4-23处理的子房ABA含量逐渐上升且始终处于较高水平,而T3Aa-23处理与之相反。授粉后120 h,各处理子房ABA含量从高到低排序为T4-23>T2-23>T1-23>T3Aa-23,其中T4-23是T3Aa-23的7.17倍(图1-C)。
2.3 不同授粉品种处理对湘猕枣授粉后子房内源激素比值的影响
授粉后30 h,T1-23处理的子房GA3/ABA比值远高于其他处理,分别是T4-23、T3Aa-23和T2-23的1.67倍、77.5倍和155倍。授粉后75 h,各处理的子房GA3/ABA比值均下降到较低水平,其中T1-23处理的比值最高,是最低的T4-23的33.75倍。授粉后120 h,各处理的GA3/ABA比值高低排序为T1-23>T3Aa-23>T2-23>T4-23,除T1-23外,其他处理的子房GA3/ABA比值均在0.1以下(图1-D)。
授粉后30 h,T4-23处理的子房IAA/ABA比值远高于其他处理,其中T1-23最低,前者是后者的19.43倍。授粉后120 h,各处理的IAA/ABA比值从高到低排序为T3Aa-23>T1-23>T2-23>T4-23,其中T3Aa-23是T4-23的4.20倍(圖1-E)。
授粉后30 h,T4-23处理的子房(GA3+IAA)/ABA比值远高于其他处理,分别是T1-23、T3Aa-23和T2-23的1.42倍、15.27倍和25.44倍。授粉后75 h,T1-23处理的子房(GA3+IAA)/ABA比值最大,是比值最小处理T4-23的9.00倍。授粉后120 h,各处理的(GA3+IAA)/ABA比值从高到低排序为T3Aa-23>T1-23>T2-23>T4-23,其中T3Aa-23是T4-23的5.18倍(图1-F)。
2.4 不同授粉品种处理对果实生长发育的影响
2022年,各授粉处理的果实生长发育曲线在各个时期基本一致,大致可分为三个时期,即快速膨大期(授粉后14~35 d)、缓慢生长期(授粉后35~63 d)和生长停滞期(授粉后63~77 d);授粉后第35天,T1-22、T2-22和T3Ad-22处理的果实纵径相较于授粉后14 d分别增长了78.86%、75.14%和45.03%,果实横径分别增长了105.86%、84.98%和88.63%;果实缓慢生长期较长,相较于授粉后第35天,第63天时T1-22、T2-22和T3Ad-22处理的果实纵径分别增长了6.73%、16.64%和19.90%,果实横径分别增长了19.96%、13.92%和24.39%;授粉后第77天,T1-22、T2-22和T3Ad-22处理的果实纵径、横径无显著差异,果实纵径均在30 mm以上,果实横径均在20 mm以上(图2-A~B)。2023年,各授粉处理果实生长发育曲线与2022年相似,即分为快速膨大期(14~42 d)、缓慢生长期(42~70 d)和生长停滞期(70~77 d)。T2-23处理的果实纵径整体显著大于T1-23和T3Aa-23;授粉后第42天,T1-23、T2-23和T3Aa-23处理的果实纵径相较于授粉后第14天,增长率分别为57.51%、93.52%和86.75%,果实横径增长率分别为116.03%、116.50%和113.78%;授粉后第70天,相较于第42天,T1-23、T2-23和T3Aa-23处理的果实纵径增长率分别为5.36%、7.42%和10.00%,果实横径增长率分别为9.38%、15.71%和15.93%;授粉后第77天,T2-23处理的果实纵径分别显著高出T1-23和T3Aa-23处理20.95%和25.91%,T2-23和T3Aa-23处理的果实横径显著高出T1-23处理的13.44%和7.36%(图2-C~D)。
2.5 不同授粉品种处理对采收时果实品质的影响
2022年,T1-22和T3Ad-22处理的平均单果质量显著高于T2-22处理,其中T1-22的平均单果质量最大,是T2-22的1.19倍;各授粉处理的果形指数均大于1.20,T1-22授粉处理的果形指数较T2-22显著高出了7.69%,也较T3Ad-22授粉处理显著高出了9.38%。T1-22处理的果实硬度和可滴定酸含量均显著高于T2-22和T3Ad-22处理,其中T1-22处理的果实可滴定酸含量分别高出T2-22和T3Ad-22处理31.40%和79.37%;T3Ad-22处理的果实可溶性固形物含量显著低于其他处理,T1-22和T2-22处理的果实可溶性固形物含量分别比T3Ad-22显著提高59.34%和57.40%(表2)。2023年,T1-23和T2-23处理的平均单果质量均显著大于T3Aa-23,其中T2-23处理的平均单果质量最大,是T3Aa-23处理的2.37倍;T1-23和T2-23处理的果形指数无显著差异,均达到1.40,但两者显著高于T3Aa-23,分别高出26.72%和20.69%;T2-23和T3Aa-23处理的果实硬度显著大于T1-23,其中T2-23处理的果实硬度最大,比T1-23提高11.27%。T1-23和T2-23处理的果实干物质及可溶性固形物含量均显著高于T3Aa-23,其中T2-23处理的果实干物质和可溶性固形物含量比T3Aa-23分别提高1.93%和1.24%;T1-23处理的果实可滴定酸含量比T2-23和T3Aa-23分别显著提高46.67%和44.74%(表2)。
2.6 不同授粉品种处理对贮藏期果实品质的影响
2.6.1 不同授粉品种处理对贮藏期果实外观形态的影响 2022年,T3Ad-22处理的果实最小且畸形;各授粉处理的果皮顏色随贮藏时间由绿转红后逐渐加深至深红色,色泽由明渐暗;果心均从果蒂端至果脐端逐渐转红;果肉颜色则是由翠绿色转为深绿色,且果实软化(图3-A)。2023年,所有处理的果皮和果肉颜色的变化相较于2022年更为缓慢,但变化趋势一致,其中T1-23处理的果皮颜色最早(贮藏后第8天)开始变红,T2-23果心在贮藏后第16天逐渐变红;T3Aa-23处理的果实最小且大多为畸形果,缺乏商品性(图3-B)。
2.6.2 不同授粉品种处理对贮藏期果实硬度的影响 两年各授粉处理的果实在不同贮藏温度下的硬度都随时间变化呈先下降后趋于平缓的趋势。2022年,T1-22和T2-22处理的果实室温贮藏(25 ℃)2 d后,硬度分别下降了89.75%和87.38%;贮藏4 d后,T3Ad-22处理的果实硬度降低了90.85%,此时三者果实硬度无显著差异;贮藏6 d后,T3Ad-22处理的果实硬度分别显著高出T1-22和T2-22处理26.17%和36.17%(图4-A)。2023年,各授粉处理的果实低温贮藏(4 ℃),果实硬度在贮藏第4~24天中无显著差异;贮藏24 d后,T2-23、T3Aa-23和T1-23处理的果实硬度相较于第0天分别降低了90.90%、89.74%和71.02%(图4-B)。
2.6.3 不同授粉品种处理对贮藏期猕猴桃果实呼吸速率的影响 2023年,贮藏期各处理的果实呼吸速率随时间呈“升-降-升-降”的变化趋势,其中T3Aa-23处理的果实呼吸速率始终高于T1-23和T2-23,T1-23和T2-23的呼吸速率相近;贮藏第24天,T1-23、T2-23和T3Aa-23处理的果实呼吸速率分别是第0天的1.61倍、1.46倍和3.24倍,T3Aa-23处理的果实呼吸速率分别高出T1-23和T2-23处理110.10%和152.39%(图5)。
2.6.4 不同授粉品种处理对贮藏期果实可溶性固形物含量的影响 2022年贮藏期间,T1-22处理的果实TSS含量始终显著高于T3Ad-22;贮藏第8天,T1-22处理的果实TSS含量分别比T2-22和T3Ad-22显著提高21.94%和15.97%(图6-A)。2023年贮藏第24天,T1-23、T2-23和T3Aa-23处理的果实TSS含量分别为16.67%、16.43%和16.73%,三者无显著差异;分别是第0天的2.19倍、2.12倍和2.47倍(图6-B)。
2.6.5 不同授粉品种处理对贮藏期果实维生素C和可滴定酸含量的影响 2023年贮藏期间各授粉处理果实的维生素C含量变化趋势不一致。贮藏第24天,各授粉处理无显著差异,相较于第0天,T1-23、T2-22和T3Aa-23处理的果实维生素C含量分别降低了25.86%、32.76%和39.68%(图7-A)。
2023年贮藏期间各授粉处理果实的TA含量变化趋势不一致。在贮藏第0、4、8和24天,T3Aa-23处理的果实TA含量显著高于T2-23和T1-23。贮藏第24天,T1-23、T2-23和T3Aa-23处理的果实TA含量相较于第0天分别降低了34.14%、22.43%和37.41%(图7-B)。
3 讨 论
湘猕枣作为一个软枣猕猴桃新品种,品质优良,风味独特,但目前还未配置授粉树,要求授粉树与雌株花期相遇、花粉量大且亲和性好,湘猕枣的广泛推广还需解决授粉树配置问题。笔者在本研究中的结果表明,DWSR1和NER1可作为湘猕枣的适宜授粉树,授粉后果大质优、风味浓郁。
花粉活力对授粉成效有重要影响,而花粉类型[28]、环境条件[29]、不同发育阶段[30]、脱药时间[31]、干燥方式[32]、贮藏条件[33]等影响花粉活力。笔者在本研究中使用的R2px2花粉并非当年收集,故其花粉活力最低,可能是其坐果率在种内花粉中最低的原因。猕猴桃种间杂交的杂交不亲和性体现在种间授粉速度慢和种胚生长异常导致的坐果率低、种子数量少和单果质量较小[13],与笔者在本研究中发现的AASAH1授粉处理的坐果率低的情况相似,说明长叶猕猴桃和软枣猕猴桃杂交不亲和。
不同倍性的花粉显著影响猕猴桃坐果率、单果质量和种子千粒质量等性状。二倍体中华猕猴桃雄株花粉给四倍体软枣猕猴桃授粉处理的坐果率远低于四倍体软枣猕猴桃和六倍体美味猕猴桃[34],表明低倍性花粉和高倍性花粉授粉处理所产生的影响是有差异的,这与本研究结果一致。不同授粉时间影响果实生长,海沃德和徐香开花后前3 d授粉的坐果率和单果质量均大于开花后4~7 d [35],AU Fitzgerald有效授粉期为4~5 d,且在第5天授粉的单果质量和种子数量降低[36]。笔者在本研究中发现,2023年DWSR1因花期稍晚于湘猕枣雌株,授粉时间为花后第7天,其余处理为开花后前3 h,授粉时间的不同可能是T1-23与其他处理的坐果率和果实品质存在差异的原因之一。而T2-22的坐果率远低于T1-22的原因可能是T2-22与T1-22授粉时间相同,此时NER1处于盛花期,而DWSR1处于大蕾期,故T2-22花粉活力可能下降,导致坐果率较低。
授粉后子房内源激素的含量和比值是影响雌株坐果率的关键因素。笔者在本研究中对湘猕枣授粉后子房中内源激素分析的结果表明,在授粉后100~120 h内,T1-23和T3Aa-23处理的子房GA3含量都有上升的趋势;同时各处理的子房IAA含量均保持上升的趋势;虽然T4-23处理的子房IAA含量在各处理之间相对不低,但其ABA含量在授粉后30 h之后逐渐上升,T1-23和T2-23处理的子房ABA含量呈波动式变化,T3Aa-23处理的子房ABA含量逐渐下降,直至授粉后120 h时,T4-23授粉处理的子房ABA含量远高于其他处理。相关研究表明,火龙果的花、果从盛花期到坐果期内GA3、IAA和ABA含量均呈上升趋势[37]。百合在授粉后8 h内,雌蕊的ABA含量变化与IAA、GA3含量变化呈相反的趋势[38],与本研究中猕猴桃在授粉后45 h内子房激素含量变化相似。由此可见,花粉不亲和性与授粉后高水平的ABA有关。油茶在授粉48 h后的雌蕊中GA3/ABA、IAA/ABA和(GA3+IAA+ZR)/ABA的比值均呈上升的趋势[39],这一结果与本研究中T1-23和T3Aa-23授粉处理在授粉100 h后的结果相似;虽然T2-23授粉处理的各激素间比值在该时期呈下降趋势,但其在授粉后55 h和100 h的GA3和IAA含量较高,达到一定积累量,为后续有效坐果奠定基础。
在本研究中,不同授粉品種对湘猕枣果实质量、果实形状、硬度、干物质含量、可溶性固形物含量和可滴定酸含量都有显著影响,其中T3Aa-23的平均单果质量显著小于其他处理,推测可能是由于花粉为2022年采集,贮藏时间较长降低了花粉活力,花粉管萌发力下降从而受精不充分,这与关于猕猴桃花粉耐贮性影响果实品质性状的研究结果相同[40]。干物质和可溶性固形物含量作为软枣猕猴桃的采收指标,在采收时,T3Aa-23处理果实的这两个指标水平均显著低于其他处理,表明不同花粉来源对果实成熟期存在明显影响,这与授粉品种对黑穗醋栗成熟期影响显著的研究结果一致[41]。在果实贮藏期间,T3Aa-23果实呼吸速率较高,且硬度下降速度相对更快,说明授粉品种可能影响果实耐贮性,这与关于金魁猕猴桃授粉品种影响其贮藏性能的研究结果相似[11]。在本研究中,2023年DWSR1和NER1授粉处理的单果质量和果形指数均大于2022年,产生差异的原因可能是授粉方式和环境气候的不同,2023年果实的可溶性固形物和干物质含量均低于2022年的原因可能是采收时间较早,2022年采收时出现了一定的落果现象,故2023年采收时间提前。
笔者在本研究中探讨了植物内源激素在授粉不亲和过程中的作用,研究了授粉品种对果实成熟期的影响,为从分子生物学水平研究软枣猕猴桃花粉直感提供了生理基础,也为该新品种的后续栽培管理及优质高效生产奠定了基础。但笔者在本研究中的授粉品种的数量较少,未来可进一步探索增加种内品种授粉组合,并重复美味猕猴桃花粉及NER1授粉处理。
4 结 论
通过使用不同授粉品种花粉处理软枣猕猴桃湘猕枣,分析不同授粉处理间果实生长与品质的差异。结果表明,以长叶猕猴桃花粉授粉处理作为种间授粉,坐果率极低;种内授粉中DWSR1和NER1授粉处理的果实品质均较高,大围山软枣猕猴桃雄株DWSR1和NER1可作为湘猕枣的适宜授粉树。
参考文献 References:
[1] 黄宏文,龚俊杰,王圣梅,何子灿,张忠慧,李建强. 猕猴桃属(Actinidia)植物的遗传多样性[J]. 生物多样性,2000,8(1):1-12.
HUANG Hongwen,GONG Junjie,WANG Shengmei,HE Zican,ZHANG Zhonghui,LI Jianqiang. Genetic diversity in the genus Actinidia[J]. Chinese Biodiversity,2000,8(1):1-12.
[2] 黄宏文. 猕猴桃属:分类、资源、驯化、栽培[M]. 北京:科学出版社,2013:50-52.
HUANG Hongwen. Actinidia taxonomy germplasm domestication cultivation[M]. Beijing:Science Press,2013:50-52.
[3] COIMBRA S,TORR?O L,ABREU I. Programmed cell death induces male sterility in Actinidia deliciosa female flowers[J]. Plant Physiology and Biochemistry,2004,42(6):537-541.
[4] NEBEL B R. Metaxenia in apples[J]. Journal of Heredity,1936,27(9):345-350.
[5] SABIR A. Xenia and metaxenia in grapes:Differences in berry and seed characteristics of maternal grape cv. ‘Narince (Vitis vinifera L.) as influenced by different pollen sources[J]. Plant Biology,2015,17(2):567-573.
[6] ZHANG H P,LIU C H,YAO J L,DENG C H,CHEN S L,CHEN J J,WANG Z H,YU Q M,CHENG Y J,XU J. Citrus mangshanensis pollen confers a Xenia effect on linalool oxide accumulation in pummelo fruit by enhancing the expression of a cytochrome P450 78A7 gene CitLO 1[J]. Journal of Agricultural and Food Chemistry,2019,67(34):9468-9476.
[7] GUPTON C L. Evidence of xenia in blueberry[C]//International Society for Horticultural Science,VI International Symposium on Vaccinium Culture,August 12,1996,Maine,Orono:Acta Horticulturae,1996,446:119-124.
[8] CALLAN N W,LOMBARD P B. Pollination effects on fruit and seed development in ‘Comice pear[J]. Journal of the American Society for Horticultural Science,1978,103(4):496-500.
[9] OLFATI J A,SHEYKHTAHER Z,QAMGOSAR R,KHASMAKHI-SABET A,PEYVAST G,SAMIZADEH H,RABIEE B. Xenia and metaxenia on cucumber fruit and seed characteristics[J]. International Journal of Vegetable Science,2010,16(3):243-252.
[10] PIOTTO F A,BATAGIN-PIOTTO K D,DE ALMEIDA M,OLIVEIRA G C X. Interspecific xenia and metaxenia in seeds and fruits of tomato[J]. Scientia Agricola,2013,70:102-107.
[11] 陈庆红,张忠慧,秦仲麒,蒋迎春. 金魁猕猴桃的雄株选配及其花粉直感研究[J]. 中国果树,1996(2):23-24.
CHEN Qinghong,ZHANG Zhonghui,QIN Zhongqi,JIANG Yingchun. Study on the selections of male plants and its xenia effects in Jinkui kiwifruit[J]. China Fruits,1996(2):23-24.
[12] SEAL A G,DUNN J K,JIA Y L. Pollen parent effects on fruit attributes of diploid Actinidia chinensis ‘Hort16A kiwifruit[J]. New Zealand Journal of Crop and Horticultural Science,2013,41(4):219-229.
[13] 齐秀娟,徐善坤,张威远,林苗苗,方金豹. 美味猕猴桃‘徐香与长果猕猴桃远缘杂交亲和性的解剖学研究[J]. 园艺学报,2013,40(10):1897-1904.
QI Xiujuan,XU Shankun,ZHANG Weiyuan,LIN Miaomiao,FANG Jinbao. Studies on compatibility of interspecific hybridization between Actinidia deliciosa ‘Xuxiang and A. longicarpa by anatomy[J]. Acta Horticulturae Sinica,2013,40(10):1897-1904.
[14] STASIAK A,LATOCHA P,DRZEWIECKI J,HALLMANN E,NAJMAN K,LEONTOWICZ H,LEONTOWICZ M,?ATA B. The choice of female or male parent affects some biochemical characteristics of fruit or seed of kiwiberry (Actinidia arguta)[J]. Euphytica,2019,215(3):52.
[15] 李伟才,魏永赞,胡会刚,石胜友,王一承,谢江辉. 3种无核荔枝果实发育过程中内源激素含量变化动态[J]. 热带作物学报,2011,32(6):1042-1045.
LI Weicai,WEI Yongzan,HU Huigang,SHI Shengyou,WANG Yicheng,XIE Jianghui. Dynamic changes of endogenous hormone contents in the pericarp of seedless litchi during fruit growth and development[J]. Chinese Journal of Tropical Crops,2011,32(6):1042-1045.
[16] CHENG X,RUYTER-SPIRA C,BOUWMEESTER H. The interaction between strigolactones and other plant hormones in the regulation of plant development[J]. Frontiers in Plant Science,2013,4:199.
[17] 李曉艳,王振兴,秦红艳,范书田,艾军. 软枣猕猴桃雌雄株花蕾发育过程中内源激素的动态变化[J]. 吉林农业大学学报,2016,38(3):281-286.
LI Xiaoyan,WANG Zhenxing,QIN Hongyan,FAN Shutian,AI Jun. Dynamic variation of endogenous hormone during male and female flower buds development of Actinidia arguta[J]. Journal of Jilin Agricultural University,2016,38(3):281-286.
[18] 李尧,张坤,周源洁,刘芯伶,夏惠,邓红红,梁东. 枝蔓牵引对金艳猕猴桃生长的影响[J]. 果树学报,2022,39(3):406-415.
LI Yao,ZHANG Kun,ZHOU Yuanjie,LIU Xinling,XIA Hui,DENG Honghong,LIANG Dong. Effect of traction of branches on the growth of Jinyan kiwifruit[J]. Journal of Fruit Science,2022,39(3):406-415.
[19] 袁月,代志国,张丙秀,王天鹤,张昭,姜婷,蔚明月. 自然越冬期软枣猕猴桃枝条组织结构及内源激素的变化特征[J]. 西北植物学报,2020,40(2):279-286.
YUAN Yue,DAI Zhiguo,ZHANG Bingxiu,WANG Tianhe,ZHANG Zhao,JIANG Ting,YU Mingyue. Changes in tissue structure and endogenous hormones of Actinidia arguta branches during over-wintering period[J]. Acta Botanica Boreali-Occidentalia Sinica,2020,40(2):279-286.
[20] 鲁敏,黄亚欣,王国立,安华明. ‘贵长猕猴桃果实内源激素的动态分布及含量变化与果实形状发育的关系[J]. 植物生理学报,2020,56(10):2159-2167.
LU Min,HUANG Yaxin,WANG Guoli,AN Huaming. The correlation between the dynamic distribution and content of endogenous hormones and the kiwi fruit shape during ‘Guichang fruits development[J]. Plant Physiology Journal,2020,56(10):2159-2167.
[21] 王利新. 软枣猕猴桃果实发育与内源激素含量变化关系的研究[D]. 长春:吉林农业大学,2023.
WANG Lixin. Study on the relationship between fruit development and changes of endogenous hormone content of Actinidia arguta[D]. Changchun:Jilin Agricultural University,2023.
[22] 牟建莉,黄晗羽,陈梦洁,唐育辉,王仁才,罗飞雄. 软枣猕猴桃新品种湘猕枣的选育[J]. 果树学报,2024,41(1):187-192.
MOU Jianli,HUANG Hanyu,CHEN Mengjie,TANG Yuhui,WANG Rencai,LUO Feixiong. Breeding report of a new Actinidia arguta cultivar Xiangmizao[J]. Journal of Fruit Science,2024,41(1):187-192.
[23] 钱峥. 猕猴桃液体授粉配方的优化及授粉效果评价[D]. 杨凌:西北农林科技大学,2021.
QIAN Zheng. Optimization of liquid pollination formula & evaluation of pollination effect of kiwifruit[D]. Yangling:Northwest A & F University,2021.
[24] 朱莉莉,陈雅雯,王棚涛,马同森,张君丽. HPLC-MS/MS同时测定植物6种内源激素含量方法的优化[J]. 河南大学学报(自然科学版),2020,50(3):298-304.
ZHU Lili,CHEN Yawen,WANG Pengtao,MA Tongsen,ZHANG Junli. Optimization of HPLC-MS/MS method for simultaneous quantification of six classes of endogenous hormones in plants[J]. Journal of Henan University (Natural Science),2020,50(3):298-304.
[25] 刘磊,李争艳,雷华,高本旺,赵佳,李薇. 30个猕猴桃品种(单株)主要果实品质特征的综合评价[J]. 果树学报,2021,38(4):530-537.
LIU Lei,LI Zhengyan,LEI Hua,GAO Benwang,ZHAO Jia,LI Wei. Comprehensive evaluation of main fruit quality characteristics with 30 kiwifruit cultivars (strains)[J]. Journal of Fruit Science,2021,38(4):530-537.
[26] 張艳霞,吕丹桂,耿康奇,王宁,王瑞,王振平. 水分胁迫对赤霞珠葡萄果实品质和甲氧基吡嗪含量的影响[J]. 果树学报,2022,39(6):1017-1028.
ZHANG Yanxia,L? Dangui,GENG Kangqi,WANG Ning,WANG Rui,WANG Zhenping. Effects of water stress on grape quality and content of methoxypyrazines in Cabernet Sauvignon[J]. Journal of Fruit Science,2022,39(6):1017-1028.
[27] 田彦龙,马永强,王磊,郭青云,陈红雨. 西北不同生态区甜樱桃果实品质分析[J]. 果树学报,2021,38(4):509-519.
TIAN Yanlong,MA Yongqiang,WANG Lei,GUO Qingyun,CHEN Hongyu. Quality analysis of sweet cherry fruits in different ecological areas in Northwest China[J]. Journal of Fruit Science,2021,38(4):509-519.
[28] 胡晋. 花粉的保存和生活力测定[J]. 种子,1992,11(6):33-35.
HU Jin. Pollen preservation and viability determination[J]. Seed,1992,11(6):33-35.
[29] 李钊. 开花期低温对花生开花及结实特性的影响[D]. 沈阳:沈阳农业大学,2022.
LI Zhao. Effects of low temperature at flowering stage flowering and fruiting characteristics of peanut[D]. Shenyang:Shenyang Agricultural University,2022
[30] 蔡昭艳,董龙,王葫青,邱文武,苏伟强,任惠,王小媚,方位宽,黄章保,邓彪,刘业强. 百香果花不同发育阶段花粉活力、柱头可授性及其对坐果的影响[J]. 果树学报,2023,40(5):969-977.
CAI Zhaoyan,DONG Long,WANG Huqing,QIU Wenwu,SU Weiqiang,REN Hui,WANG Xiaomei,FANG Weikuan,HUANG Zhangbao,DENG Biao,LIU Yeqiang. Pollen viability,stigma receptivity and their effect on fruit set of passionfruit at different flower developmental stages[J]. Journal of Fruit Science,2023,40(5):969-977.
[31] 陈厚锡,杨技超,王胜艳,潘丽珊,罗充,李苇洁. 影响猕猴桃花粉活力的因素探讨[J]. 中国果树,2021(9):59-62.
CHEN Houxi,YANG Jichao,WANG Shengyan,PAN Lishan,LUO Chong,LI Weijie. Discussion on the factors of affecting the activity of kiwifruit polle[J]. China Fruits,2021(9):59-62.
[32] 姚春潮,龙周侠,刘旭峰,王西芳. 不同干燥及贮藏方法对猕猴桃花粉活力的影响[J]. 北方園艺,2010(20):37-39.
YAO Chunchao,LONG Zhouxia,LIU Xufeng,WANG Xifang. Effects of different dryness and storage methods on pollen viability in Actinidia deliciosa[J]. Northern Horticulture,2010(20):37-39.
[33] 陈永安,陈鑫,刘艳飞. 采粉期及贮藏条件对猕猴桃花粉生活力的影响[J]. 西北农林科技大学学报(自然科学版),2012,40(8):157-160.
CHEN Yongan,CHEN Xin,LIU Yanfei. Effects of different collecting pollen periods and storage conditions on pollen viability of kiwifruit[J]. Journal of Northwest A & F University (Natural Science Edition),2012,40(8):157-160.
[34] 李志,方金豹,齐秀娟,林苗苗,陈锦永,顾红. 不同倍性雄株对软枣猕猴桃坐果及果实性状的影响[J]. 果树学报,2016,33(6):658-663.
LI Zhi,FANG Jinbao,QI Xiujuan,LIN Miaomiao,CHEN Jinyong,GU Hong. Effects of male plants with different ploidy on the fruit set and fruit characteristics in Actinidia arguta kiwifruit[J]. Journal of Fruit Science,2016,33(6):658-663.
[35] 白雪,刘占德,李建军,王西芳,蒋宏勤,姚春潮. 猕猴桃花后不同天数授粉效果研究[J]. 江苏农业科学,2020,48(7):166-168.
BAI Xue,LIU Zhande,LI Jianjun,WANG Xifang,JIANG Hongqin,YAO Chunchao. Study on pollination effect of different days after flowering for kiwifruit[J]. Jiangsu Agricultural Sciences,2020,48(7):166-168.
[36] BRANTLEY A K,SPIERS J D,THOMPSON A B,PITTS J A,KESSLER J R,WRIGHT A N,CONEVA E D. Effective pollination period of Actinidia chinensis ‘AU Golden Sunshine and A. deliciosa ‘AU Fitzgerald kiwifruit[J]. HortScience,2019,54(4):656-660.
[37] 张瀚,杨福孙,胡文斌,孙会举,李洪立. 火龙果生长发育过程中内源激素含量变化[J]. 江苏农业科学,2022,50(10):110-116.
ZHANG Han,YANG Fusun,HU Wenbin,SUN Huiju,LI Hongli. Changes of endogenous hormone content during growth of pitaya[J]. Jiangsu Agricultural Sciences,2022,50(10):110-116.
[38] 楊晓苓,杨利平,尚爱芹,刘凤栾. 百合授粉亲和性与雌蕊中保护酶和激素的关系[J]. 园艺学报,2009,36(6):855-860.
YANG Xiaoling,YANG Liping,SHANG Aiqin,LIU Fengluan. Relationship between protective enzymes,endogenous hormones in pistil and pollination compatibility of Lilium[J]. Acta Horticulturae Sinica,2009,36(6):855-860.
[39] 杜明,于旭东,吴繁花. 海南油茶授粉后雌蕊内源激素的动态变化[J]. 热带生物学报,2023,14(2):173-177.
DU Ming,YU Xudong,WU Fanhua. Dynamic changes of endogenous hormones in self-pollinated and cross-pollinated pistils of two Camellia species in Hainan[J]. Journal of Tropical Biology,2023,14(2):173-177.
[40] SEYREK U A,LUO M,ZHONG M,HUANG C H,TAO J J,QU X Y,XU X B. Effects of stored pollens from wild Actinidia eriantha vines on some fruit quality traits[J]. Agricultural Sciences,2017,8:465-478.
[41] 高洪娜,张武杰,刘凤芝,周文志,汤佳翰,徐德海,张帅. 黑穗醋栗晚丰花粉直感效应研究[J]. 种子,2023,42(7):91-95.
GAO Hongna,ZHANG Wujie,LIU Fengzhi,ZHOU Wenzhi,TANG Jiahan,XU Dehai,ZHANG Shuai. Study on the Xenia of Wanfeng Ribes nigrum L.[J]. Seed,2023,42(7):91-95.
