两个小麦品种的抗旱性鉴定与评价
2024-04-27郑祥博白昕晨王伟伟刘曙东奚亚军
郑祥博 白昕晨 王伟伟 刘曙东 奚亚军

摘 要 为探索小麦品种‘西农100和‘周麦18的抗旱特性,采用大田和盆栽组合试验,通过测定根部性状、萌发率、生理指标和显微结构等,分析其抗旱性。结果表明:‘西农100的根质量、平均根数、根表面积与体积显著低于‘周麦18,在水培25 d时,‘西农100的总根长与最大根长显著高于‘周麦18。两个品种在PEG胁迫下的发芽率都达到了90%以上,苗期抗旱性等级均为极强。两个品种在苗期干旱胁迫下的幼苗存活率较高,相对含水量无显著差异。‘西农100在水分临界期受到干旱胁迫后,其产量显著低于‘周麦18;两个品种的旗叶抗氧化酶(SOD、POD、CAT)活性、渗透调节物质(Pro、可溶性蛋白)含量与小麦的抗旱性呈正相关,复水后抗氧化酶活性随时间逐渐降低至对照水平,渗透调节物质含量显著降低;‘西农100和‘周麦18在干旱胁迫下的叶片均出现明显结构变化;总体来说,在水分临界期‘西农100抗旱性整体弱于‘周麦18。
关键词 小麦(Triticum aestivum L.);西农100;周麦18;抗旱性;生理指标
干旱缺水已成为影响植物健康生长和制约农业生产发展的主要逆境因素之一 ,旱灾不仅会造成粮食作物减产,还会促使其他次生灾害的发生[1-2]。作物的抗旱性是指作物在缺水干旱的环境下生长发育和形成籽粒产量的能力,它是一种由多基因相互调控而表现在各种形态和生理变化中的复杂生物特征[3-4]。旱灾一般可分成避旱型、耐旱型和抗旱型3种类型,快速生长发育与较早成熟是避旱类型作物一般的特点,从而避免了旱害[5]。植株根系发达且生命力旺、分蘖期长且有较强分蘖力是抗旱性作物的一般特点,其在水分亏缺条件下,还能够比较顺利的生长与发育。而耐旱型作物由于渗透调节力较强,对不同组织器官的抗脱水处理能力较强,它在干旱条件下耐受时间长[6-7]。
作物在遇到逆境胁迫时,往往会有大量活性氧产生而形成氧化逆境,这会使植物细胞的细胞膜和某些生物大分子化合物受到破坏,植物细胞在抗氧化威胁时生成除去活性氧的酶和抗氧化物质,从而高效的除去活性氧,比如过氧化物酶(POD)、超氧化物歧化酶(SOD)、过氧化氢酶(CAT)等都是常见的抗氧化酶,这些抗氧化酶活性增高可以提高植物的抗逆能力[8-9]。阎成仕等[10]研究表明超氧化物歧化酶(SOD)活力、过氧化氢酶(CAT)活力和谷胱甘肽(GSH)浓度在小麦水分亏缺下均呈现出下降的趋势,抗旱性弱的品种下降幅度大于抗旱性强的品种。周雪英等[11]发现小麦拔节期水分亏缺后复水可以使超氧化物歧化酶(SOD)活力、谷胱甘肽还原酶(GR)和抗坏血酸过氧化物酶(AP)的活性显著下降。王敏等[12]研究发现小麦孕穗期和抽穗期水分较为敏感,这两个时期水分亏缺对小麦产量的影响最大,小麦蜡熟期水分亏缺有增产效果。黄正金等[13]研究发现小麦在抽穗期干旱胁迫时会使产量变化,旗叶抗氧化酶(SOD、CAT、POD)活力均显著增加,丙二醛(MDA)的浓度显著上升,且与胁迫程度呈正比。
小麦是仅次于水稻和玉米的第三大粮食作物,为全球超过1/3人口提供了主食,不断增长的环境压力给小麦研究和育种者带来了重大挑战,研究小麦的抗旱性能对其应用推广具有重要价值[14-15]。‘周麦18是国家黄淮南片及河南省冬水组区域试验对照品种,具有高产、稳产、适应性广等显著特点[16];‘西农100是2020年通过审定的国审品种,由于其矮杆、大穗、高产、适应性强,已在黄淮南片麦区展示出强劲的推广势头。本研究以‘周麦18作为对照,通过对国审小麦品种‘西农100的根系结构、叶片抗逆性生理指标和显微结构等进行测定,比较了‘西农100和‘周麦18的抗旱能力,以期为‘西农100的推广应用提供参考依据。
1 材料与方法
1.1 试验材料
供试小麦品种为2020年通过国审的小麦品种‘西农100(国审麦20200076)和黄淮麦区南片小麦区域试驗对照品种‘周麦18(国审麦2005006),均由农业农村部西北地区小麦生物学与遗传育种重点实验室提供。
1.2 品种苗期根部发育特性的研究试验
‘西农100和对照‘周麦18各选大小均匀、籽粒饱满的种子150粒,先用无菌水浸泡3 h,再用8%的次氯酸钠溶液(体积比)灭菌10 min后蒸馏水洗涤至无刺鼻性异味,后置于规格为长×宽×高=10 cm×10 cm×5 cm的发芽盒(单层滤纸)中,诱导萌发7 d并保持发芽盒中水分充足, 7 d后将发芽完成的小麦幼苗移至7 000 mL的包含1/2 Hoagland营养液黑色塑料盒(规格为34 cm×27 cm×13 cm)并置于光照培养箱中(温度周期20/18 ℃,光周期14/10 h,相对湿度 70%±5%),培养10 d、15 d、20 d、25 d后取材进行试验,2个品种各重复3次。
测定小麦幼苗根部性状,采用根系扫描仪(MicrotekScanmakeri800 plus)扫描幼苗根部并保存图像,利用万深LA-S 系列植物图像分析系统对图像进行分析并获取根系形态数据:根数(RN)、总根长(TRL)、根表面积(RSA) 和根体积(RV) ;用直尺人工测量最大根长(MRL);幼苗自根茎相接处分开,称量地上部鲜质量(SFW)和根鲜质量(RFW),烘箱 105 ℃杀青 20 min,70 ℃烘干至恒质量,称量地上部干质量(SDW)和根干质量(RDW),并计算根冠比(RSR)和地上部含水量(SWC)和根含水量(RWC)。每个品种不同时期各测定30株。
1.3 小麦品种抗旱性鉴定试验
1.3.1 小麦品种种子萌发期抗旱性鉴定试验 采用高渗溶液法,利用PEG-6000(-0.5 MPa)水溶液对种子进行水分胁迫处理,对照处理采用蒸馏水培养,以底部放置单层滤纸的塑料盒(10 cm×10 cm×5 cm)为发芽盒。‘西农100和‘周麦18品种各取大小均匀、籽粒饱满的种子800粒,其中胁迫处理400粒,对照400粒,100粒为一个重复置于一个发芽皿中,各重复四次。将发芽盒放入培养箱中(温度周期20/18 ℃,光周期14/10 h,相对湿度70%±5%),第8天调查发芽种子数。
种子发芽率计算:
1.3.2 小麦品种苗期抗旱性鉴定试验 苗期抗旱性鉴定试验采用盆栽实验,于2021年10月9日在西北农林科技大学南校区温室进行,采用底部带孔的塑料盆(直径30 cm,高度26 cm)中装入10 cm厚中等肥力壤土,灌水(田间持水量 85%±5%),‘西农100与‘周麦18品种各播300粒,每盆30粒各10盆(6处理+3取样+1 CK),其中3次重复,每个重复60苗,温室温度在 20 ℃±5 ℃。每天18:00-23:00进行补光,顶部补光光强为500 μmol/(m2·s)。
第1次干旱胁迫及复水处理:
幼苗长至三叶时,进行干旱胁迫(停止供水),持续测定盆栽土壤含水量,土壤含水量为田间持水量的15%~20%时复水,复水后使土壤水分在田间持水量的80%±5%左右,复水120 h后调查存活苗数,判定幼苗存活标准为叶色转为鲜绿色。
第2次干旱胁迫及复水处理:
第1次复水后进行第2次干旱胁迫(停止供水),后续方法与第1次干旱胁迫相同。
幼苗第1次干旱存活率记为DS1(%),第2次干旱存活率记为DS2(%)。
叶片含水量测定:
测定叶片相对含水量(RWC)。剪取长势一致的小麦旗叶(第1次处理与第2次处理),收获后立即进行鲜质量(Wf)测定,将叶片放入去离子水中浸泡过夜充分吸水,取出用吸水纸擦干表面水分进行称质量(Wt),在105 ℃杀青15 min, 80 ℃烘干至恒质量(Wd),每个处理3个生物学重复。
RWC=(Wf-Wd)/(Wt-Wd ) ×100%
逆境生理指标的测定:
在第1次干旱胁迫复水120 h后和第2次干旱胁迫复水120 h后分别选取长势一致的叶片取样,取样后液氮速冻,保存在-80 ℃。进行相关逆境生理指标的测定,每个处理重复3次。其中超氧化物歧化酶(SOD)活性测定参照氮蓝四唑(NBT)光化还原法,过氧化物酶(POD)活性测定参照愈创木酚法,过氧化氢酶(CAT)活性测定参照过氧化氢还原法,抗坏血酸过氧化物酶(APX)活性参考Vicente等[17]的方法测定,可溶性蛋白质含量测定参照考马斯亮蓝G-250染色法,可溶性总糖和还原糖的测定参照蒽酮比色法,脯氨酸含量参照酸性茚三酮法测定,丙二醛(MDA)含量参照硫代巴比妥酸法测定,超氧阴离子自由基 (O-·2)产生速率的测定参照羟胺氧化法。
1.3.3 水分临界期抗旱性鉴定试验 采用盆栽试验,试验于2020-10-09在西北农林科技大学南校区温室展开,盆底(直径30 cm,高度26 cm)底部有小孔,保證水分和空气流通,2020-10-09种植,每盆6株,两个品种各12盆(6处理+6 CK),播种后将盆埋入田间,待返青期(2021-01-13)移栽温室,拔节期前温室温度控制在16 ℃,拔节期后温度白天20 ℃夜晚16 ℃,每天8:30-22:30进行补光,小麦植株顶部补光光强为500 μmol/(m2·s)。
胁迫处理:在抽穗期、灌浆期浇水使土壤相对含水量保持在80%±5%(表2)。
对照处理:在拔节-孕穗期、抽穗期、灌浆期浇水使土壤相对含水量保持在80%±5%。
考察性状:籽粒产量
抗旱指数计算:DI=(GY2S.T/GYS.W)×(GYCK.W/GY2CK.T)
DI为抗旱指数(表3);GYS.T为待测材料胁迫处理籽粒产量;GYS.W为待测材料对照处理籽粒产量;GYCK.W为对照品种对照处理籽粒产量;GYCK.T为对照品种胁迫处理籽粒产量。
在胁迫复水前、胁迫复水后12 h、24 h、48 h,从对照和干旱处理中选取长势一致的旗叶取样,其中部分旗叶立刻进行叶片相对含水量的测定,剩余旗叶取样后液氮速冻,保存在-80 ℃备用,用于逆境生理指标的测定。待小麦自然生长至成熟,对其穗部性状进行调查:第1、2小花结实粒数、小穗数、穗粒数。每个处理调查3盆,求平均结实率。
1.4 干旱胁迫对旗叶显微结构的影响
采用CaseViewer植物显微成像分析系统观察切片。在干旱前与复水后12 h分别从干旱处理和对/照中选取长势一致的倒二叶片进行取样,再截取距主脉中心1 cm 处的叶片固定于FAA固定液中,采用番红固绿染色法制作石蜡切片,切片厚度为在120 mm,采用OLYMPUS实体显微镜(SZX16)观察叶片解剖结构变化。
2 结果与分析
2.1 小麦品种苗期根部性状分析
根据图1,两个品种苗期地上部鲜质量(SFW)、地上部干质量(SDW)、根鲜质量(RFW)、根干质量(RDW)、鲜根冠比(FRSR)、干根冠比(DRSR)、平均根数(RN)、体积(RV)、总根长(TRL)、最大根长(MRL)与根表面积(RSA)。SFW、SDW与培养时间成正比,‘西农100显著小于‘周麦18;RFW与RDW整体变化趋势与SFW和SDW相同,在10、15、20 d时,‘西农100与‘周麦18具有显著差异;‘西农100的FRSR显著高于‘周麦18,25 d内两个品种的FRSR变化没有差异,DRSR有明显差异变化,在10 d时达到最大0.32左右,15~25 d下降到0.2左右;在10~20 d中,两个品种RSA与RV增速平缓,每5 d增速为20.3%,20~25 d增速为56.7%;‘西农100与‘周麦18的TRL值随时间增长而变大,且在 10 d、15 d、20 d时‘周麦18显著高于‘西农100,而在25 d时‘西农100显著高于‘周麦18,达到了400 cm;两个品种的MRL值在10 d时差异不显著,长度达到 10.5 cm,15 d时‘周麦18显著大于‘西农100,20 d和25 d时‘西农100显著大于‘周麦18 ,25 d ‘西农100的MRL达到了24 cm。RN在10 d时两个品种都在6.7根左右,25 d时‘西农100显著小于‘周麦18,RN平均少1.8根。
2.2 种子萌发期抗旱性鉴定
从表4和图2可以看出,干旱胁迫(PEG-6000)下小麦‘西农100与‘周麦18种子萌发率(GerT)为96.75%和96.5%,比对照(未处理)的萌发率低2.5%和3.25%,两个品种的相对发芽率(RGer)都达到了90%以上。在种子萌发期,两个品种的抗旱性等级均为极强(HR)。
2.3 苗期干旱胁迫对幼苗存活率的影响
苗期干旱胁迫对‘西农100与‘周麦18的影响如表5所示,苗期第1次干旱胁迫‘西农100与‘周麦18的幼苗存活率都在96%以上,第2次干旱胁迫后‘西农100的幼苗存活率下降了 4.44%,‘周麦18幼苗存活率为93.89%,较第1次干旱胁迫下降了3.89%。‘西农100的幼苗存活率略低于‘周麦18,两个品种整体的幼苗存活率高。
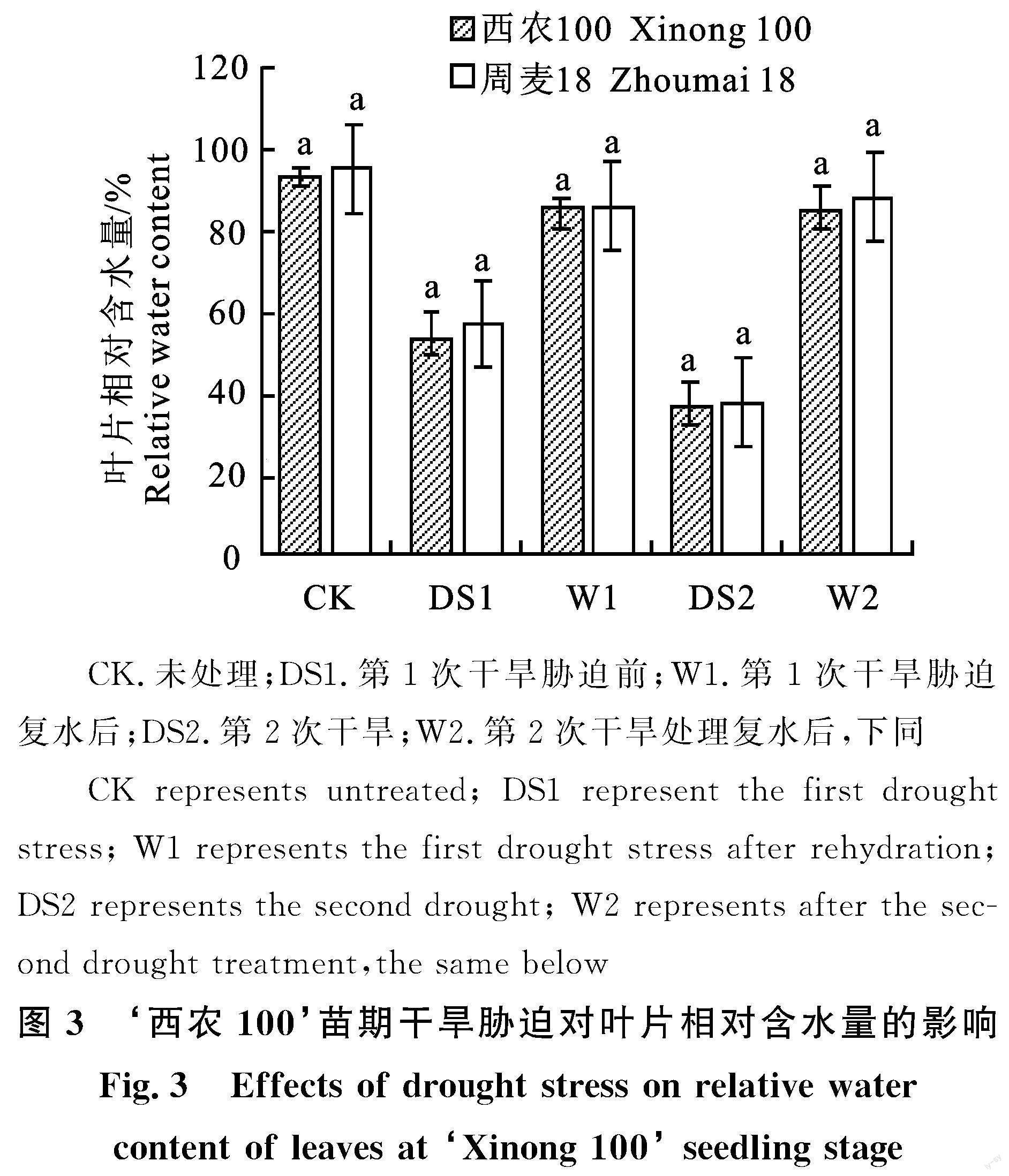
2.4 苗期干旱胁迫对叶片相对含水量的影响
苗期干旱胁迫后小麦叶片相对含水量(RWC)变化如图3所示,CK(未处理)的RWC在93%以上,第1次干旱胁迫后小麦叶片RWC下降至55%,复水后RWC有所回升,较CK相差10%,第2次干旱胁迫叶片RWC降至40%以下,较第1次胁迫时下降45%,复水后叶片RWC恢复至第1次胁迫复水后水平,‘西农100各时期叶片RWC略低于‘周麦18,两个品种整体叶片RWC无显著差异。
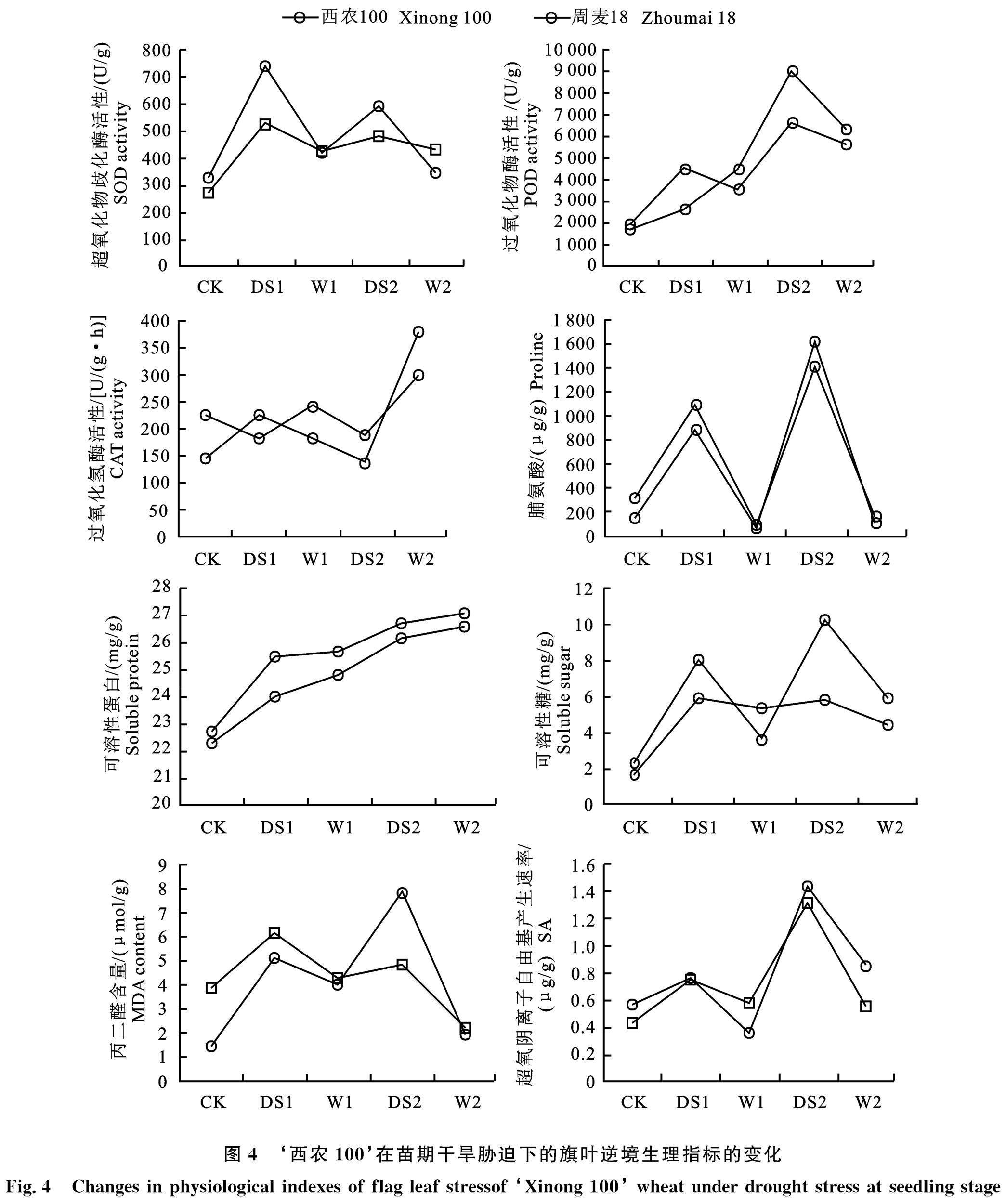
2.5 苗期干旱胁迫对旗叶逆境生理指标的影响
由图4可知,‘西农100与‘周麦18在苗期干旱胁迫后旗叶逆境生理指标变化,SOD(超氧化物歧化酶)活性整体呈“双峰”变化趋势,‘西农100较‘周麦18的 SOD活性变化更大,第1次干旱胁迫后两个品种的SOD活性较CK(未处理)明显增大,复水后SOD活性下降,第2次干旱后‘西农100的SOD活性明显增大,‘周麦18的SOD活性小于‘西农100,复水后两个品种SOD活性下降至第1次复水后水平;苗期干旱胁迫对POD(过氧化物酶)活性有明显影响,‘西农100 的POD值受干旱胁迫后增加,复水后下降,而‘周麦18在第1次干旱胁迫与复水后POD活性增大不明显,第2次干旱胁迫后‘周麦18 的POD活性明显增大,复水后下降;在第2次干旱复水前,‘西农100 的CAT活性先升后降,而‘周麦18表现为先降后升,两个品种POD活性变化范围都在150~250 U/(g·h),第2次干旱复水后两个品种的CAT活性显著上升。
干旱胁迫使脯氨酸含量变化明显,两个品种受到干旱胁迫时,脯氨酸含量显著升高(900~ 1 500 μg/g),复水后又迅速降低与CK相同水平(300 μg/g),第2次干旱脅迫较第1次明显增高,两个品种脯氨酸含量变化无明显差异;可溶性蛋白含量在小麦苗期受干旱胁迫后明显上升,复水后增速减小,‘西农100可溶性蛋白含量大于‘周麦18;可溶性总糖含量表现为受干旱胁迫后上升,复水后下降,两个品种变化趋势相同,‘西农100变化速率整体大于‘周麦18,第2次干旱胁迫后‘西农100 可溶性糖含量较‘周麦18增加60%;两个品种受到干旱胁迫后MDA(丙二醛)含量明显增大,复水后MDA含量下降,‘西农100不同于‘周麦18,表现在第1次干旱胁迫MDA含量大于第2次干旱胁迫,‘周麦18在第2次干旱胁迫时MDA含量达到最大。超氧阴离子自由基产生速率在受干旱胁迫时有明显增大,第2次干旱胁迫大于第1次,且复水后速率下降,‘西农100与‘周麦18无明显差异。
2.6 水分临界期抗旱性鉴定
2.6.1 水分临界期抗旱性判定 拔节-孕穗期干旱胁迫对对籽粒产量的影响如表6所示,‘西农100与‘周麦18在水分临界期抗旱性存在差异,‘西农100的抗旱指数(DI)为0.91,在0.90~ 1.09之间属于水分临界期抗旱性中等(MR),‘周麦18较‘西农100抗旱指数(DI)更高,达到了 1.11,判定水分临界期抗旱性强。
2.6.2 水分临界期干旱后复水对品种旗叶逆境生理指标的影响 由图5可知,‘西农100与‘周麦18在水分临界期干旱胁迫复水前后旗叶逆境生理指标的动态变化,SOD(超氧化物歧化酶)活性胁迫处理较CK(未胁迫处理)变化更明显,干旱胁迫使SOD活性明显升高,复水后受胁迫处理的SOD活性明显降低,复水48 h后与CK无明显差异,SOD活性在两个品种间差异较小。POD(过氧化物酶)活性在复水后干旱处理与CK无明显差异,复水前(0 h)干旱处理POD活性明显小于CK,两个品种的POD活性存在差异,‘西农100 POD活性在复水后呈先上升后下降趋势,而‘周麦18 的POD活性整体呈先下降后上升趋势。CAT(过氧化氢酶)活性整体呈先上升后下降的趋势,干旱胁迫后‘西农100较‘周麦18 的CAT活性反应更大,两个品种在复水后24 h的CAT活性达到最大峰值,CK的CAT活性变化起伏较小。
干旱后复水两个小麦品种的MDA(丙二醛)含量有所不同,复水前‘西农100的MDA含量高于‘周麦18,复水后‘西农100 MDA含量呈先下降后上升趋势,而‘周麦18 MDA含量呈先上升后下降趋势,两个品种在复水后24 h的MDA含量降至与CK处理相近程度。胁迫处理与CK的脯氨酸含量差异明显,CK的脯氨酸含量一直保持在200 μg/g 以下,而干旱处理后小麦的脯氨酸含量在900 μg/g左右,复水后脯氨酸含量有所下降,在复水后48 h达到了最低为400 μg/g,明显高于CK。干旱胁迫后旗叶的可溶性蛋白明显大于CK,‘西农100大于‘周麦18,复水后‘西农100的可溶性蛋白含量明显下降,与CK相差较小,而‘周麦18在复水后24 h后的可溶性蛋白含量有所下降。‘西农100与‘周麦18可溶性总糖与可溶性还原糖的变化趋势相近,可溶性总糖与可溶性还原糖的CK处理变化平缓,而干旱处理下两者都呈先下降后上升的变化趋势,其中‘西农100的可溶性总糖与还原糖在干旱胁迫后含量较‘周麦18高,复水后两个品种的可溶性总糖与可溶性还原糖有所下降,在12 h后上升至与复水前相同状态。
2.6.3 水分临界期干旱胁迫对品种叶片显微结构的影响 叶片距主脉中心1 cm 处的横切面整体与局部结构如图6和图7所示。干旱胁迫使叶片失水过多,泡状细胞发生萎蔫,‘西农100与‘周麦18叶片较CK(未处理)内卷,复水后细胞吸水膨胀、叶片展开,两个品种间无明显差异。局部结构图中可以看出干旱胁迫降低了小麦的叶片厚度,叶肉细胞出现明显萎缩,细胞间出现空洞,复水后叶片明显增厚,复水使得叶片中细胞膨大,表皮细胞與叶肉细胞间隙缩小,保持了细胞膨压,缓解干旱胁迫对叶片细胞的损伤。
3 结论与讨论
‘周麦18是河南省周口市农业科学院选育的半冬性、稳产、广适的小麦新品种,于2008年被定为国家黄淮南片及河南省冬水组区域试验对照种。‘西农100则是2020年通过审定的国审品种,具有矮杆、大穗、高产、适应性强等优点。本研
究以‘周麦18作为对照,通过测定‘西农100的根部性状、萌发率、显微结构等指标,解析‘西农100的抗旱能力,可以为‘西农100的推广应用提供了一定的参考依据。
小麦根系是吸收水分与养分的主要器官,苗期初生根在整个生育期均发挥重要作用,其形态特征影响小麦的产量性状[18]。研究发现‘西农100苗期根系前期生长慢,后期最大根长(MRL)达到400 cm,且显著大于‘周麦18,根质量(RFW和RDW)、根表面积(RSA)与体积(RV)明显增大。赵佳佳等[19]研究发现最大根长(MRL)越长,根生物量(RFW和 RDW)越大,小麦苗期的抗旱性越高,MRL、RFW和RDW可以作为小麦品种早期筛选抗性和产量的指标。因此,推断出‘西农100苗期抗旱性好、田间产量高与品种苗期MRL长、RFW和RDW大有密切 关系。
干旱胁迫是环境中最普遍的逆境胁迫,使植物细胞遭受氧化威胁,超氧化物歧化酶(SOD)、过氧化物酶(POD)、过氧化氢酶(CAT)等保护酶类能维持活性氧自由基产生与清除系统的平衡[20-21]。大量研究表明小麦的抗旱性与水分胁迫下体内的抗氧化酶(SOD、POD、CAT)活性高低呈正相关[22-23]。通过对‘西农100干旱胁迫的研究,发现干旱胁迫后小麦旗叶抗氧化酶(SOD、POD、CAT)活性较CK(未处理)明显增加,复水后抗氧化酶活性降低至正常水平。
渗透调节是作物对逆境胁迫的一种重要适应方式,作物通过自身细胞的渗透调节作用来抵抗外界渗透胁迫,从而降低细胞水势以保持植物体内膨压正常,以维持植物对土壤水分的吸收利用[24]。脯氨酸(Pro)含量和活性的增加有利于保持植物蛋白质和细胞膜的稳定性,清除逆境胁迫下细胞内大量积累的活性氧,维持亚细胞结构,特别是液泡膜的功能及完整性,以保护细胞功能的正常发挥[25]。本研究中,水分临界期(拔节—孕穗期)干旱胁迫后,渗透调节物质脯氨酸(Pro)含量明显高于CK(未处理),复水后Pro含量明显降低,但仍显著高于CK,品种间无明显差异。Pro是有效的渗透调节物质,可以增加植株组织的保水力,但脯氨酸与抗旱性之间的关系仍有争论,大部分研究认为逆境胁迫会使脯氨酸含量增加,只是植株对外部环境的适应表现,并不能代表抗逆性强,脯氨酸的积累与抗逆性的关系需进一步研究。可溶性蛋白是有效的渗透调节物质之一[26]。本研究中‘西农100苗期干旱胁迫后,其叶片可溶性蛋白含量大幅增加,水分临界期干旱胁迫后复水,‘西农100叶片可溶性蛋白含量变化呈先升后降的趋势,这与赵丽英[27]研究出干旱导致蛋白水解并使含量下降的结果不一致,也有研究表明,植物适度干旱时,可溶性蛋白含量会增加,亲水性的蛋白可以增强原生质的水合度,而植物在受到重度干旱胁迫下,可溶性蛋白合成受阻,并含量降低[28],由此推断出本研究干旱胁迫程度对于‘西农100与‘周麦18均不属于重度胁迫。研究表明叶片的可溶性糖含量可以作为抗旱性鉴定的重要指标,干早胁迫会使叶片中可溶性糖含量的增高,为植株生长提供正常生长所需的物质基础,胁迫解除后,叶片中可溶性糖快速降低,从而解除叶片同化物积累对光合作用的抑制,提高生长速率并补偿物质积累量,与本研究中苗期和水分临界期干旱胁迫会使可溶性糖明显增高结论一致[29]。
叶片是小麦蒸腾和同化的主要器官,研究表明在干旱胁迫下其形态结构会受到的影响,己经发育完全叶可能会萎蔫、保持与太阳辐射方向平行和卷曲,从而减轻过热和水分过度散失。小麦整体叶片数减少,叶片相对含水量降低,衰老加速,叶片细胞增大受抑制,表皮细胞和叶肉细胞变小而密,海绵细胞变小,当水分恢复后叶片也很快恢复原状,适度干旱胁迫对小麦叶片属于可逆变化。本研究发现‘西农100与‘周麦18受干旱胁迫后叶片相对含水量有显著降低,叶片显微结构较CK出现卷曲与变薄,液泡细胞出现萎缩,叶肉细胞间出现空洞,复水后表皮细胞膨大,叶片明显增厚,基本与前人的研究结果相符[29-30]。
本研究从形态学、生理学角度出发解析了‘西农100的根系特征、叶片抗逆性生理指标和显微结构等,确定了‘西农100和‘周麦18都具有一定的抗旱能力,但‘西农100的抗旱性整体要稍弱于‘周麦18。研究结果为国审小麦品种‘西农100的推广应用提供了初步的理论参考。
参考文献 Reference:
[1] 景蕊莲.作物抗旱节水研究进展[J].中国农业科技导报,2007,9(1):1-5.
JING R L.Advances of research on drought resistance and water use efficiency in crop plants[J].Journal of Agricultural Science and Technology,2007,9(1):1-5.
[2] MUELLER B,SENEVIRATNE S I.Hot days induced by precipitation deficits at the global scale[C]//EGU General Assembly Conference Abstracts,2013:12398-1240.
[3] 王 静,康林玉,刘周斌,等.干旱对植物影响的研究进展[J].湖南农业科学,2017(7):123-126,130.
WANG J,KANG L Y,LIU ZH B,et al.Research progress on the impact of drought on plant[J].Hunan Agricultural Sciences,2017(7):123-126,130.
[4] 馬孝松,曾贤军,李恩熙,等.水稻耐旱性及其研究进展[J].上海农业学报,2022,38(4):36-45.
MA X S,ZENG X J,LI E X,et al.Advances of drought tolerance researches in rice[J].Acta Agriculturae Shanghai,2022,38(4):36-45.
[5] [WB]田艳丽,种培芳,陆文涛,等.不同光合途径植物红砂和珍珠猪毛菜幼苗对氮沉降及降水变化的光合响应[J].草地学报,2021,29(1):121-130.
[DW]TIAN Y L,ZHONG P F,LU W T,et al.Photosynthetic responses of seedlings of Reaumuria soongorica and Salsola passerina with different photosynthetic pathway to nitrogen deposition and precipitation changes[J].Acta Agrestia Sinica,2021,29(1):121-130.
[6] 庄 晔,葛嘉雪,汪孝国,等.干旱胁迫后复水对烤烟生长及其生理特性的影响[J].中国烟草学报,2022,28(4):48-58.
ZHUAN Y,GE J X,WANG X G,et al.Effect of rehydration after drought stress on the growth and physiological characteristics of flue-cured tobacco[J].Acta Tabacaria Sinica,2022,28(4):48-58.
[7]张梦飞,李 爽,李运盛,等.9种绿化树种幼苗抗旱性评价[J].中国农学通报,2022,38(20):38-46.
ZHANG M F,LI SH,LI Y SH,et al.Evaluation on the drought resistance of seedlings of nine greening tree species[J].Chinese Agricultural Science Bulletin,2022,38(20):38-46.
[8] 刘 畅,王维领,赵 灿,等.小麦倒春寒研究现状及展望[J].江苏农业学报,2022,38(4):1115-1122.
LIU CH,WANG W L,ZHAO C,et al.Research progress and prospect of cold spell in later spring in wheat[J].Jiangsu Journal of Agricultural Sciences,2022,38(4):1115-1122.
[9] 梁培鑫,唐 榕,郭晨荔,等.油莎豆对自然盐碱胁迫的生长及生理响应[J].中国农学通报,2022,38(26):1-8.
LIANG P X,TANG R,GUO CH L,et al.Growth and physiological response of Cyperus esculentus L. to natural saline-alkali stress[J].Chinese Agricultural Science Bulletin,2022,38(26):1-8.
[10] 阎成仕,李德全,张建华.冬小麦旗叶旱促衰老过程中氧化伤害与抗氧化系统的响应[J].西北植物学报,2000, 20(4):568-576.
YAN CH SH,LI D Q,ZHANG J H.Oxidative damage and antioxidant responses during drought-induced winter wheat flag leaf senescence[J].Acta Botanica Boreali-Occidentalia Sinica,2000,20(4):568-576.
[11]周雪英,邓西平.旱后复水对不同倍性小麦光合及抗氧化特性的影响[J].西北植物学报,2007,27(2):278-285.
ZHOU XY,DENG X P.Effect of post-drought rewatering on leaf photosynthetic characteristics and antioxidation in different wheat genotypes[J].Acta Botanica Boreali-Occidentalia Sinica,2007,27(2):278-285.
[12]王 敏,张从宇,姚维传,等.不同生育期干旱胁迫对小麦产量的影响[J].安徽农业科学,2001,29(5):605-607,610.
WANG M,ZHANG C Y,YAO W CH,et al.Effects of drought stress in different development stages on wheat yield[J].Journal of Anhui Agricultural Sciences,2001,29(5):605-607,610.
[13] 黄正金,丁锦峰,李春燕,等.抽穗期干旱胁迫对小麦产量及生理特性的影响[J].中国科技论文,2017,12(18):2141-2145.
HUANG ZH J,DING J F,LI CH Y,et al.Effects of drought stress at heading stage on grain yield and physiological characteristics of wheat[J].China Sciencepaper,2017,12(18):2141-2145.
[14] 吴秀宁,敬 樊,张 军,等.PEG-6000模拟干旱胁迫对黑小麦种子萌发特性的影响[J].作物研究,2022,36(4):307-312.
WU XN,JING F,ZHANG J,et al.Effects of PEG-6000 on seed germination of black-grain wheat varieties under simulated drought condition[J].Crop Research,2022, 36(4):307-312.
[15] 蘇培森.小麦赤霉病抗病机制研究进展[J].生物技术进展,2021,11(5):599-609.
SU P S.Research advances in wheat FHB resistance mechanism[J].Current Biotechnology,2021,11(5):599-609.
[16] 吴和平.不同播种密度对周麦18号小麦生长发育及产量的影响[J].现代农业科技,2019(23):3.
WU H P.Effects of different sowing densities on growth and yield of Zhoumai 18 wheat[J].Modern Agricultural Science and Technology,2019(23):3.
[17] VICENTE A R,GUSTAVO A M,CHAVES A R,et al.Effect of heat treatment on strawberry fruit damage and oxidative metabolism during storage[J].Postharvest Biology and Technology,2006,40(2):116-122.
[18] LIAO M,FILLERY IRP,PALTA J A.Early vigorous growth is a major factor influencing nitrogen uptake in wheat[J].Functional Plant Biology,2004,31(2):121-129.
[19]赵佳佳,乔 玲,武棒棒,等.山西省小麦苗期根系性状及抗旱特性分析[J].作物学报,2021,47(4):714-727.
ZHAO J J,QIAO L,WU B B,et al.Seedling root characteristics and drought resistance of wheat in Shanxi province[J].Acta Agronomica Sinica,2021,47(4):714-727.
[20] 张立新,李生秀.氮,钾,甜菜碱对水分胁迫下夏玉米叶片膜脂过氧化和保护酶活性的影响[J].作物学报,2007, 33(3):482-490.
ZHANG L X,LI SH X.Effects of nitrogen,potassium and glycinebetaine on the lipid peroxidation and protective enzyme activities in water-stressed summer maize[J].Acta Agronomica Sinica,2007,33(3):482-490.
[21] 彭昌操,孙中海.低温锻炼期间柑桔原生质体SOD和CAT酸活性的变化[J].华中农业大学学报,2000,19(4):384-387.
PENG CH C,SUN ZH H.Changes of SOD and CAT enzyme activities in citrus protoplasts during low temperature exercise[J].Journal of Huazhong Agricultural University,2000,19(4):384-387.
[22] 武仙山,昌小平,景蕊莲.小麦灌浆期抗旱性鉴定指标的综合评价[J].麦类作物学报,2008,28(4):626-626.
WU X SH,CHANG X P,JING R L.Screening indexes for drought resistance of wheat at grain-filling stage[J].Journal of Triticeae Crops,2008,28(4):626-626.
[23] 陳 军,高贵珍,方雪梅,等.干旱胁迫对小麦萌发期POD、CAT活性的影响研究[J].农业科学与技术:英文版,2013,14(11):5360-5361.
CHENJ,GAO G ZH,FANG X M,et al.Effects of drought stress on activity of POD and CAT in wheat during generation period[J].Agricultural Science & Technology,2013,14(11):5360-5361.
[24] 蒋明义,郭绍川.氧化胁迫下稻苗体内积累的脯氨酸的抗氧化作用[J].植物生理学报,1997,23(4):347-352.
JIANG M Y,GUO SH CH.Antioxidant effects of proline accumulation in rice seedlings under oxidative stress[J].Plant Physiology Journal,1997,23(4):347-352.
[25] 赵 璞,李 梦,及增发,等.植物干旱响应生理对策研究进展[J].中国农学通报,2016,32(15):86-92.
ZHAOP,LI M,JI Z F,et al.Counter-measures of drought physiology response in plant[J].Chinese Agricultural Science Bulletin,2016,32(15):86-92.
[26] 颜 华,贾良辉,王根轩.植物水分胁迫诱导蛋白的研究进展[J].生命的化学,2002,22(2):5355-5357.
YANH,JIA L H,WANG G X.Research progress of water stress inducible proteins in plants[J].Chemistry of life,2002,22(2):5355-5357.
[27] 赵丽英.小麦对水分亏缺的阶段性反应及其机制研究[D].陕西杨凌:西北农林科技大学,2005.
ZHAO L Y.Study on the phase responses and mechanisms of different winter wheat varieties to drought[D].Yangling Shaanxi:Northwest A&F University,2005.
[28] 马旭凤.水分亏缺对玉米生理指标、形态特性及解剖结构的影响[D].陕西杨凌:西北农林科技大学,2010.
MA X F.Effects of water deficit on maize physiology,morphology and anatomical structure[D].Yangling Shaanxi:Northwest A&F University,2010.
[29] 彭祚登,李吉跃,沈熙环.林木抗旱性育种的现状与策略思考[J].北京林业大学学报,1998,20(4):101-106.
PENG Z D,LI J Y,SHEN X H.Recent status and strategy analysis of drought resistant breeding of forest tree species[J].Journal of Beijing Forestry University,1998,20(4):101-106.
[30] 苟作旺,杨文雄,刘效华.水分胁迫下旱地小麦品种形态及生理特性研究[J].农业现代化研究,2008(4):503-505.
GOU Z W,YANG W X,LIU X H.Configuration and physiological characteristics of dryland spring wheat under water stress conditions[J].Research of Agricultural Modernization,2008(4):503-505.
Identification and Evaluation of Two Wheat Varieties under Drought Stress
Abstract To explore the drought resistance characteristics of wheat varieties ‘Xinong 100 and ‘Zhoumai 18 , a combination of field and pot experiment was conducted to analyze the drought resistance of the wheat varieties by measuring the root character, germination rate, physiological index and microscopic structure. The results showed that the root mass, root average number, root surface area and volume of ‘Xinong 100 were significantly lower than those of ‘Zhoumai 18 , and the total root length and maximum root length of ‘Xinong 100 were significantly higher than those of ‘Zhoumai 18 after 25 days of hydroponics. The germination rate of the two varieties under PEG stress reached more than 90%, and the drought resistance grade of the seedling stage was identified as very strong. The seedling survival rate of the two varieties was extremely strong under drought stress, and there was no significant difference in relative water content. The yield of ‘Xinong 100 was significantly lower than that of ‘Zhoumai 18 after drought stress at critical moisture stage. The activities of antioxidant enzymes (SOD, POD, CAT) and the contents of osmotic regulatory substances (Pro, soluble protein) in flag leaves of the two varieties were positively correlated with the drought resistance of wheat. The activity of antioxidant enzymes decreased gradually to the control level after rehydration, and the content of osmotic regulatory substances decreased significantly. The leaves of ‘Xinong 100 and ‘Zhoumai 18 showed obvious structural changes under drought stress. In general, the drought resistance of ‘Xinong 100 is weaker than that of ‘Zhoumai 18 duing the critical moisture period. The comparison between the drought resistance of ‘Xinong 100 and ‘Zhoumai 18 was conducted by measuring several traits and indexes.This study provides a reference for the popularization and application of these two varieties.
Key words Wheat(Triticum aestivum L.); Xinong 100; Zhoumai 18; Drought resistance; Physiological indexes
