ABO 血型变异的分子基础研究
2024-04-26雷航王学锋程晓文张慧蔡晓红
雷航 王学锋 程晓文 张慧 蔡晓红
(1.上海交通大学医学院附属瑞金医院 输血科,上海 200025;2.复旦大学附属闵行医院 输血科)
ABO 血型系统由血型之父Karl Landsteiner 于1900 年发现,在临床输血、器官移植、新生儿溶血病的诊断和筛查中发挥着重要作用[1-3]。 ABO 亚型是1 种具有以下血清学特征的遗传性ABO 表型:红细胞上抗原的强度减弱、消失、出现混合视野,血清中伴有不规则抗-A 或抗-B 出现[4-5]。 而类孟买血型因H 抗原的缺失,影响A、B 抗原的合成,进而影响着ABO 血型的准确鉴定[6-7]。
ABO 亚型与类孟买血型是导致ABO 血型变异的重要原因,临床上均可以导致ABO 血型定型困难、血型误判、甚至急性溶血性输血反应的发生[8-9]。 ABO 亚型与类孟买血型复杂的血清学表现,我们仅依靠常规血清学实验已无法满足临床安全供血和器官移植等精准医学发展需求,随着近年来分子生物学的发展,基因分析逐渐成为ABO 亚型和类孟买血型鉴定的重要组成部分[10-13],但也有相关文献报道:1 种亚型表型可对应多种基因型,如B(A)血型有6 种等位基因(BA.01—BA.06)[14];同1 种基因型也可对应多种表现型,如等位基因BW.12 血清学上可以出现:Bx、Bm、ABx、ABm、AB3等多种表型[3];甚至同1 个基因位点突变也可以产生不同的表型,如c.425T>C 发生在等位基因B.01 上突变可产生混合视野B3表型[15],而c.425T>C 若发生在等位基因A1.02 基础上的突变则会产生Ael表型[16]。 这提示我们在遇到ABO 血型变异时,需要将传统经典的血清学方法同现代的基因检测方法结合起来,方能提高对血型变异的鉴定准确率。 我们对在临床工作中遇到的ABO 血型变异标本进行了表型与基因型的关联性分析,现报告如下。
1 材料与方法
1.1 标本来源
纳入标准:1)2022 年2 月至2022 年12 月上海交通大学医学院附属瑞金医院(以下简称瑞金医院)门急诊/住院部2.42 万名血型鉴定患者(重复检测的患者只计入首次);2)2022 年9 月外院送检瑞金医院的ABO 疑难血型鉴定标本10 例,其中疑似ABO 亚型3 例,疑似类孟买血型7 例。 以上标本类型均为EDTA 抗凝外周静脉全血。
排除标准:1)新生儿血型标本;2)血液科疾病患者标本;3)近2 月内有ABO 非同型输注史的患者标本。 研究对照:随机选取120 名ABO 血型正常的外科患者的标本。
1.2 试剂与仪器
单克隆抗-A(批号:20220102)、抗-B(批号:20220102)、抗-A1(批号:20211128)、抗-H 血清(批号:20211110),人ABO 血型反定型红细胞试剂(批号:20225303), 不规则抗体筛查细胞(批号:20227004),均为上海血液生物医药有限公司产品;人源多克隆抗-A、抗-B 来自本课题组自制,效价128;抗-Lea(批号:8000451442)、抗-Leb血清(批号:8000262294)均为荷兰Sanquin 公司产品;血液基因组DNA 抽提试剂盒(批号:X0615)、DNA 琼脂糖凝胶纯化试剂盒(批号:X1014)均为北京天根生物; Ex TaqTMVersion 2.0 plus dye ( 批 号:ALE2528A)、pMD18-T 克隆载体(批号:AL51603A)均为日本TAKARA 公司产品。 全自动血型仪(Ortho AutoVue,强生);KUBOTA 离心机(KA-2200,久保田);瞬时离心机(5424R,Eppendorf);DNA 扩增安全柜(SCV 4A1,Airtech); PCR 扩增仪(Proflex3,ABI);基因测序仪(3730xl,ABI)。
1.3 方法
1.3.1血型血清学检测
采用全自动血型分析仪对患者的ABO 血型及不规则抗体筛查进行检测,对初筛中显示ABO 正反定型不符的疑为ABO 亚型标本和外地送来已初筛过疑似ABO 亚型和类孟买血型的标本按如下方法进行ABO 亚型和类孟买血型的确认试验:弱ABO(H)抗原或抗体检测法、增强法、吸收放散试验,根据血型血清学国际标准方法AABB[17]对ABO 亚型、类孟买血型进行鉴定。 由于未获取到患者的唾液标本,故我们使用Lewis 血型鉴定方法来间接地对个体的分泌状况进行判断。 亚型分类判断按文献[4]判断。
1.3.2基因组DNA 抽提与PCR 扩增
血清学鉴定为ABO 亚型、类孟买血型的患者标本,进行外周血基因组DNA 抽提,对ABO基因的7 个外显子(E1—E7)及其侧翼序列、增强子、启动子进行PCR 扩增,类孟买血型则对FUT1、FUT2基因序列进行扩增。ABO、FUT1、FUT2 基因突变筛查的扩增引物序列及扩增条件按参考文献[3,18]进行。
1.3.3PCR 产物纯化、测序和克隆
采用DNA 琼脂糖凝胶纯化试剂盒对PCR 产物进行纯化后测序,单倍体鉴定时,将ABO基因E6、E7、Intron6 的PCR 产物或FUT1 基因扩增产物克隆到pMD18-T 载体做TA 克隆,挑取多个阳性菌落,抽提质粒DNA 进行测序鉴定,确定标本的单体型[19]。
1.3.4ABO基因、FUT1 和FUT2 基因新突变与等位基因的命名方式
对测序结果分析到的含有基因突变的序列,进行再一次的PCR 扩增和测序确认,以排除PCR 扩增引入基因突变的可能性。 根据国际输血协会(ISBT)命名方式对新突变及ABO、FUT1 和FUT2等位基因进行命名。
2 结果
2.1 ABO 亚型的表型检测
2022 年2—12 月,瑞金医院门急诊/住院部2.42万血型鉴定患者中,检出7 例ABO 亚型(编号1—7)。 外院送检瑞金医院的ABO 疑难血型鉴定10 例标本中,共鉴定出2 例ABO 亚型:编号Ⅰ-1、Ⅱ-1(第3 例鉴定为正常A 型,故未纳入亚型统计结果内)。 一共检出的9 例ABO 亚型中含Ael亚型2 例、B3亚型3 例、B(A)亚型1 例、Bweak亚型2 例、A2Bweak亚型1 例。 其中B 亚型6 例,占比67%(6/9),A 亚型2 例,占比22%(2/9),血清学鉴定结果如表1。
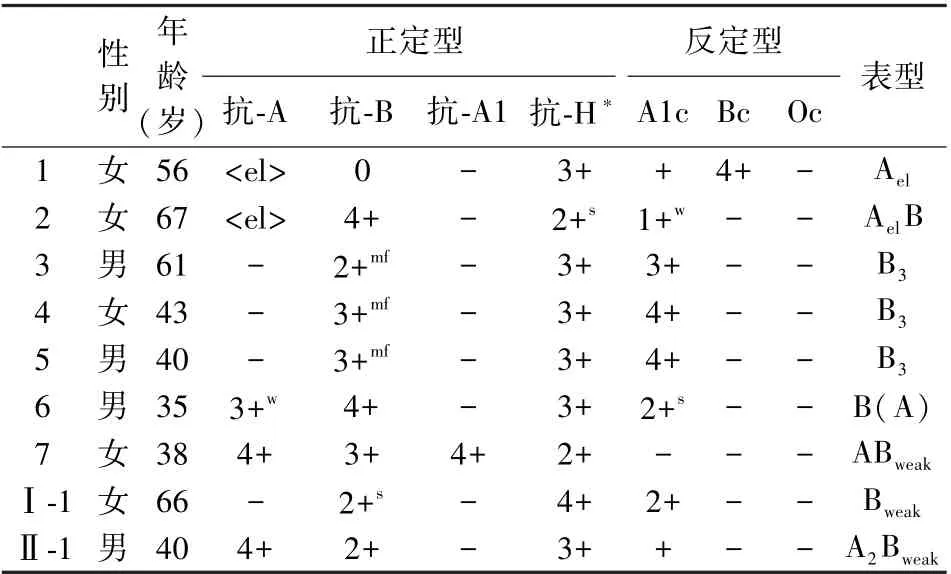
表1 9 例ABO 亚型血清学鉴定结果Table 1 Results of serological identification of ABO subgroup in 9 cases
2.2 ABO 亚型的基因型检测
9 例ABO 亚型标本,经DNA 直接测序、克隆后测序共鉴定出7 种罕见等位基因,9 例ABO 亚型的表型及其对应基因型分别为:1 例Ael(AEL.02/O.01.02)、1 例AeB(AEL.05/B.01)、3 例B3(2 例B3.03/O.01.01、1 例B3.03/O.01.02)、1 例B(A)(BA.02/O.01.01)、1 例ABweak(A1.02/BW.07)、1 例Bweak(BW.31/O.01.02)、1 例A2Bweak(A2.05/BW.31),见表2。
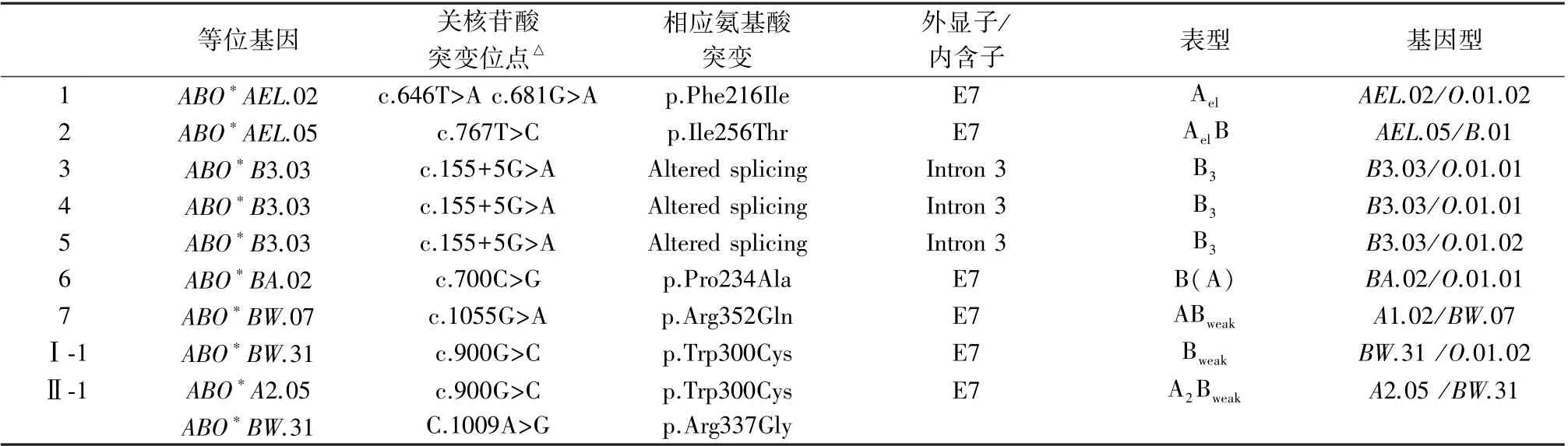
表2 9 例ABO 亚型基因型鉴定结果Table 2 Results of genotype identification of ABO subgroup in 9 cases
2.3 7 例类孟买血型鉴定
来自外院送检瑞金医院ABO 疑难血型鉴定10例标本中,共鉴定出7 例类孟买血型,血清学结果及测序结果见表3,其中7 名患者红细胞上均未检出A、B、H 抗原,仅通过吸收放散试验检出红细胞上极微弱的A、B 抗原;Lewis 血型鉴定1 例为Lewis(a-b-),余6 例均为Lewis(a-b+),间接证明以上均为分泌型;7 名患者血清中均检出抗-H。FUT1 的2 个主要突变位点c.547delAG (FUT1*01N.06)、c.880delTT(FUT1*01N.13)的测序结果如图1。
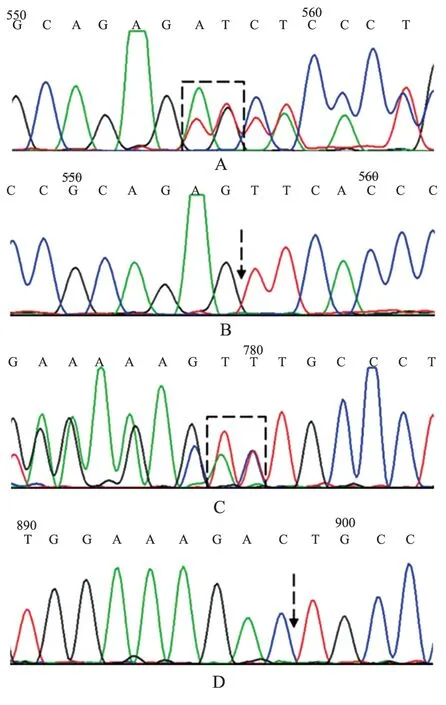
图1 FUT1 基因部分测序结果Figure 1 Results of FUT1 gene sequencing
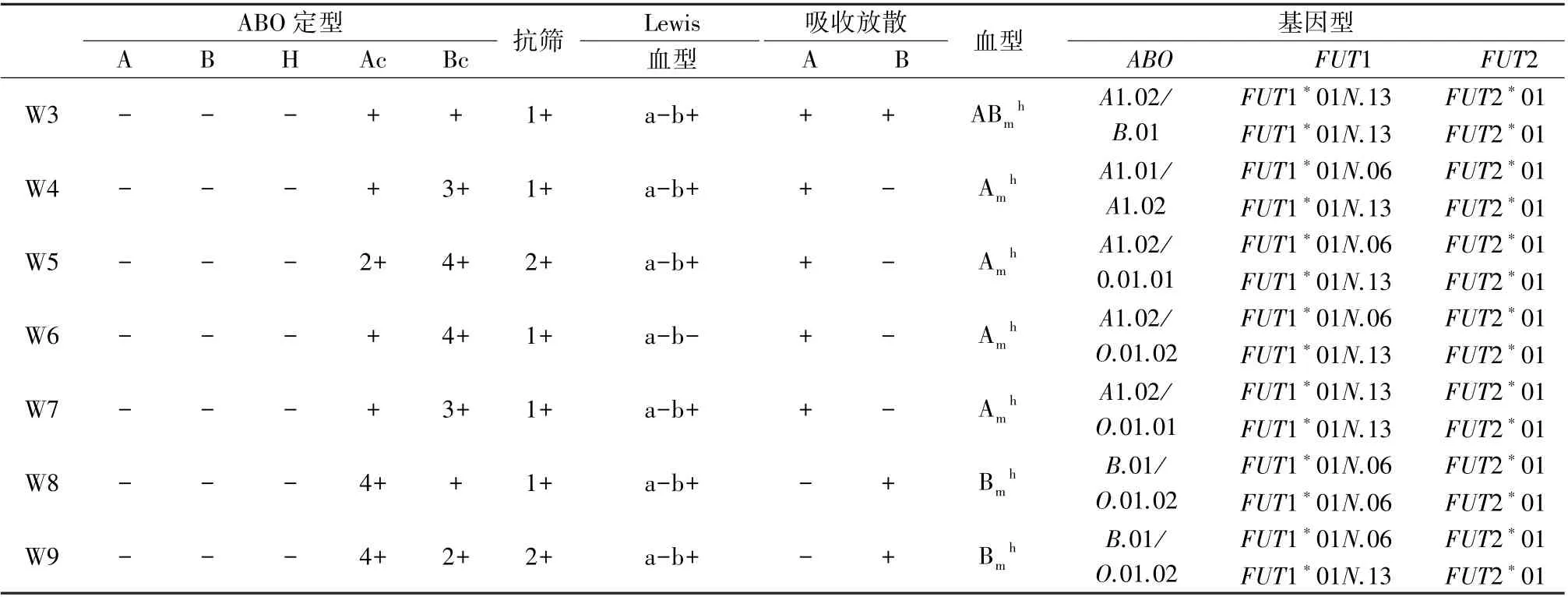
表3 7 例类孟买血型鉴定结果Table 3 Results of para-Bombay blood group identification in 7 cases
3 讨论
ABO 血型抗原表达受控于ABO基因,但ABO基因表达的初级产物是α1,3-N-乙酰氨基半乳糖转移酶(GTA)或α1,3-D-半乳糖基转移酶(GTB),在GTA、GTB 催化下,分别将供体底物UDP-Gal-NAc、UDP-Gal 转移到H 糖链上形成A、 B 抗原,人类ABO基因位于9 号染色体,由7 个外显子和6个内含子组成[20-21]。 目前已知与ABO 亚型有关的ABO基因突变类型包括:点突变、插入、缺失和基因重排等,由于突变类型以及突变位点的不同,导致ABO 亚型产生的机理也不尽相同[14]。 本研究将对工作中检出的亚型进行表型与基因型关联性分析,以探讨其潜在的分子机制,从而达到准确鉴定血型,保障临床输血、器官移植等安全。
本研究检出2 例Ael亚型,Ael血清学特点为正定型通常不被抗-A、抗-AB 及抗-A1凝集,但经吸收放散试验常可检出A 抗原[16];与抗-H 试剂反应凝集强度明显增强,常与正常O 型红细胞接近[4];血清中存在不规则抗-A,这与正定型的A 亚型或者抗-A 减弱的O 型血清学上常难以区分,故有必要结合分子生物学对Ael患者血型进行准确鉴定。 我们鉴定的1 例Ael亚型基因型AEL.02/O.01.02,等位基因A1.01 的7 号外显子上发生c.467C>T(p.Pro156Leu)、c.646T>A(p.Phe216Ile)和c.681G>A突变,已知c.467C>T 并不影响酶的活性,而c.681G>A 为同义突变,因而c.646T>A 突变是导致GTA活性减弱的主要原因。 相关文献用3D 模型预测c.646T>A 突变给GTA 带来的影响:未引入新的氢键,未改变酶的空间结构,但影响了酶的稳定性下降是造成酶活性减弱进而导致Ael亚型形成的原因[22]。 检出的另1 例AelB 亚型基因型AEL.05/B.01,等位基因A1.01 上发生c.467C>T 和c.767T>C(p.Ile256Thr)突变,高晶等[23]利用3D 结构模型分析发现,在野生型256Ile 和突变型256Thr 这2 个蛋白结构中,均是256 位氨基酸的主链氧和Arg248位的侧链氢之间形成氢键,突变部位局部结构变化不大,但突变后的热动力学稳定性改变值达到2.83 kcal/moL,因而很可能是突变导致酶的稳定性下降,产生Ael表型。
检出3 例B3亚型,B3是指红细胞上的B 抗原与抗-B 和抗-AB 试剂发生2+及以上的特征性混合视野,而血清中一般无抗-B,唾液中含有正常量的B 物质和H 物质[1]。 这类亚型与血液疾病导致的获得性血型变异及血型嵌合体外观上难以区分,均有混合型凝集现象,故难以进行血清学分类,基因水平上的进一步研究成为必要。 随后我们对此3例标本进行了基因型鉴定,鉴定均为等位基因B3.03,是发生在内含子3 上的c.155+5G>A 突变。 c.155+5G>A 最早由2012 年Chen 等[24]报道并阐述了其导致B3亚型的分子机制:c.155+5G>A 突变引起剪接错误,产生至少7 种可变剪接类型,但超过99%的剪接转录物不产生功能性蛋白,并且预测具有外显子3 缺失的剪接转录物中不到1%的能产生功能性GTB,而此GTB 糖基转移酶仅具有正常GTB 功能的一半。 这2 种相互作用机制共同解释了弱B 抗原表达的机理,并最终导致B3典型混合视野。
检出1 例B(A),B(A)血型如同cisAB 一般特殊,其血型表观学不符合孟德尔遗传规律,而是以顺式遗传为特点,红细胞上含有接近正常含量的B抗原和弱A 抗原[25]。BA与不同的A基因或者B基因存在时,由于等位基因的竞争作用,除了表现出经典的B(A)表型外,还会表现出不同的血清学特点,比如与A基因共表达时会出现ABx 表型甚至出现正常的AB 型,而与B基因共表达时会出现AxB 表型[14],加上不同实验室所用抗血清效价差异,使得复杂的B(A)表型与AxB、ABx 亚型很难在血清学上区分开来。 目前B(A)亚型等位基因共有6 种(BA.01—BA.06),具有基因型种类集中、明确的特性。 已知BA.02 是在等位基因B.01 基础上发生c.700C>G(p.Pro234Ala)突变。 纵观等位基因BA.01 至BA.06,我们发现,除了GTA 和GTB 酶自身区别的4 个氨基酸外,只有2 个位点氨基酸(214和234) 的突变会改变酶的特异性:BA.02(p.Pro234Ala)、BA.04(p.Met214Val) 和BA.05(p.Met214Thr),而且这2 个位点的突变均是在GTB上,使得B等位基因具有编码双功能酶能力,产生了弱的GTA 酶活性,从而合成弱A 抗原,产生B(A)表型。
3 例涉及Bw的表型,分别为:ABw1 例(A1.02/BW.07),Bw1 例(BW.31/O.01.02),A2Bw1 例(A2.05/BW.31)。 迄今为止国际上共发现34 种BW等位基因:BW.01—BW.34。BW.07 是在等位基因B.01 基础上发生c.1055G >A(p.Arg352Gln)基因突变,血清学呈ABw,这与国内相关报道一致[26]。BW.31 是母子两人,再次证明ABO 亚型突变等位基因呈遗传性继承,其突变位点是c.900G >C(p.Trp300Cys)。BW这些等位基因我们可以看出突变未造成GTB 酶的特异性改变,但却很可能改变了酶局部结构,降低了酶的稳定性,进而削弱了酶的活性,造成了红细胞上B 抗原合成的减弱,这也从客观上部分解释了抗原抗体在质和量上出现差异性的原因[27]。
本研究共检出7 例类孟买血型,类孟买血型具有复杂的基因多态性和遗传异质性特点,我们需要联合基因检测对其血清进行准确鉴定并探讨其发生的可能机制,从而为临床安全供血提供依据。 目前发现FUT1 有60 多种等位基因,大致分为3 类:H+、H+weak及H-基因型,类孟买血型属于H-(Null alleles)基因型,最常见的是:h1(c.547delAG)、h2(c.880delTT)、h3(c.658C >T)和h4(c.35C >T)型[28]。 本研究共检出1 例h1h1、4 例h1h2 和2 例h2h2。 其中h1 分子机制:碱基AG 缺失,阅读框移码突变,终止密码子前移,不仅影响了第184—267位氨基酸,而且在第268 位氨基酸处终止合成链,形成截短的酶,H-岩藻糖基转移酶失去活性,不能合成H 抗原,使得红细胞上的ABH 抗原缺失或极弱表达[29]。 同样h2 也是碱基的缺失发生框移突变,终止密码子前移,在第333 位氨基酸处终止合成链,合成后的氨基酸链长度只有正常的90%[30],突变使得酶的活性严重降低或者失活,不能合成H抗原,产生类孟买血型。 7 例类孟买血型患者的FUT2 均存在c.357C>T 突变,该突变在我们汉族人群中普遍存在,未改变编码氨基酸(天冬酰胺),属于同义突变,不影响SE-岩藻糖基转移酶活性,仍为分泌型。 另外FUT2 我们还检出含c.716G>A 杂合突变4 例,纯合突变2 例,虽然该点突变属于错义突变,氨基酸由p.Arg239Gln 置换,但该突变不会导致SE-岩藻糖基转移酶活性改变,只是增加了SE基因的多态性[31]。 故本研究的7 例类孟买血型均是由FUT1 基因缺陷导致,而FUT2 正常的H 抗原缺失的分泌型。
综上所述,我们共鉴定出9 例ABO 亚型、7 例类孟买血型,阐述了表型与基因型之间关联性,揭示了这2 种血型产生的可能分子机制,为ABO 血型的准确鉴定与预判提供精确的基因检测靶点和理论依据,从而为临床安全输血、器官移植、胎母免疫性溶血病的预测与防治提供参考。
