蔷薇科果树花色苷的研究进展
2024-04-24邢昱肖坤苏凯张晨光刘春生李刚李晓颍
邢 昱 肖坤 苏凯 张晨光 刘春生 李刚 李晓颍


摘 要:花色苷是一种存在于植物组织和器官中,赋予它们丰富色彩并具有多种生物活性的色素,在食品、医疗及保健品行业具有广泛的应用前景。果实中的花色苷是影响果实品质形成的重要因素。本研究通过综合国内外蔷薇科果树花色苷的研究进展,阐述了花色苷的生物合成途径、转录调控以及影响花色苷生物合成的因素,使人们更深入地了解花色苷在果实生长发育过程中的代谢方式与途径,同时对今后高效利用花色苷提供了前瞻性的建议。
关键词:蔷薇科果树;花色苷;果实
中图分类号:S661.1文献标识码:ADOI编码:10.19440/j.cnki.1006-9402.2024.01.001
蔷薇科果树往往具有较高的经济价值,如苹果亚科中的苹果、梨、山楂,李亚科的桃、李、杏、樱桃,还有蔷薇亚科的草莓、树莓等果树。蔷薇科果实大多色彩艳丽,花色苷含量丰富。花色苷(Anthocyanin)是广泛存在于植物的果实、花、叶、茎、根等组织液泡中的一种水溶性植物色素,也是果实品质重要的组成成分之一[1-2]。花色苷在不同酸碱度或金属离子作用下可呈现红色、蓝色等颜色,因而可赋予植物组织和器官五彩缤纷的色彩[3]。花色苷主要影响高等植物根、叶、茎、花和果等器官,研究表明大多蔷薇科果树不同组织均有花色苷物质的存在。在苹果[4]果皮中,桃[5]、李[6]和樱桃[7]果肉中,颜色的形成都与花色苷的存在密切相关。
花色苷的抗氧化和自由基清除的生物活性可以预防多种疾病,对维持眼睛健康、调节代谢等方面均可发挥一定作用[8]。目前花色苷已成为育种研究和果实评价的重要指标。开展果实花色苷的研究不仅有助于理解花色苷合成与代谢途径,还对选育新品种时高效利用花色苷,提高果实品质具有重要的意义。
1 花色苷的生物合成与转录调控
1.1 花色苷的结构 花色苷由花色素和糖经糖苷键缩合而成[9]。花色苷的结构如图1所示,其基本碳骨架为C6-C3-C6[10],两个芳香环A和B由一个含氧六元杂环C隔开,是2-苯基苯并吡喃阳离子的多羟基和多甲氧基衍生物[11]。花色苷结构上的差异决定了其生物学性状和功能的多样性[12]。
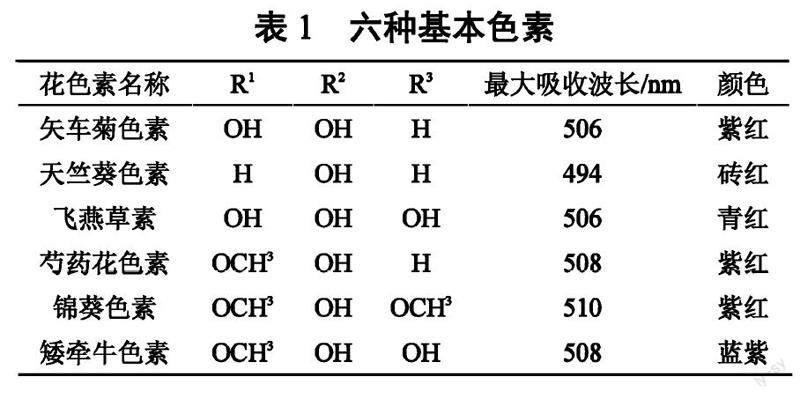
6种花色苷取代基位置及颜色见表1,植物中花色苷主要以矢车菊素(Cyanidin)、天竺葵素(Pelargonidin)、飞燕草素(Delphenidin)、芍药素(Peonidin)、锦葵色素(Malvidin)、矮牵牛素(Petunidin)这六种形态存在[4]。六种色素在A环和C环上具有相同的结构,但B环上有不同数量的羟基和甲氧基[13]。矢车菊素是自然界中最常见的花色苷,在B环的C3和C4处有两个羟基[14],在果实中表现橙色或红色。天竺葵素是主要的花色苷组成成分之一,在B环的C4位发生羟基化,主要表现为红色或洋红色。飞燕草素在B环的C3、C4和C5有三个羟基,并表现为紫色或蓝色。芍药素是通过C3位置的单一甲基化产生的,锦葵色素则由C3和C5位置的双氧甲基化而来。矮牵牛素是由飞燕草素在C3上再进行一次甲基化而来的[15]。
花色苷的共轭结构使其在紫外光区和可见光区都有显著吸收,吸收波长范围在494~510 nm。花色苷的吸光度受到羟基或甲氧基在B环上的取代影响,B环上的羟基取代越多,花色苷在可见光范围内的最大吸收就越大。B环上邻位和对位羟基的存在也会促进更长波长的光吸收和红移[16]。因此,花色苷使植物器官呈现出众多的色彩,也在果实上表现出颜色特有的特征。
1.2 花色苷的生物合成途径 花色苷生物合成途径是类黄酮类生物合成途径的分支之一[17]。花色苷合成是由苯丙氨酸通过苯丙烷代谢途径合成香豆酰辅酶A,然后参与类黄酮的合成,接着与丙二酰辅酶A反应生成查尔酮,并在内环生成二氢黄酮醇后分配到不同的途径,最后分别合成各类型花色苷,其中涉及到多种酶的配合[18]。图2是花色苷的生物合成途径[19]。
PAL:苯丙氨酸解氨酶;C4H:肉桂酸4-羟化酶;4CL:4-香豆酸辅酶A连接酶;CHS:查尔酮合酶;CHI:查尔酮异构酶;F3H:黄烷酮?-羟化酶;F3′5′H:类黄酮?′5′羟化酶;F3′H:类黄酮?′羟化酶;DFR:二氢黄酮醇4-還原酶;ANS:花色苷合酶;UFGT:黄酮-3-葡糖基转移酶;GST:谷胱肽S转移酶;MATE:多药毒性排除转运体;ABC:ATP结合盒。
在第一阶段以苯丙氨酸和乙酸作为黄酮类物质的直接合成前体,在苯丙氨酸解氨酶(PAL)、肉桂酸-4-羟化酶(CH4)、4-香豆酸辅酶A连接酶(4CL)和乙酸-CoA羧化酶的催化作用下合成4-香豆酰CoA和丙二酰CoA这两个底物[20]。上述两个底物在第二阶段进行反应,首先是在查尔酮合酶(CHS)的催化下发生缩合反应,从而合成四羟基查尔酮,紧接着在查尔酮异构酶(CHI)催化作用下生成类黄酮的直接前体物质,即柚皮素,随后在黄烷酮3-羟化酶(F3H)催化作用下,生成羟基化产物二氢黄酮醇。在类黄酮3'5'羟化酶(F3'5'H)、类黄酮5'羟化酶(F5'H)的催化下二氢黄酮醇会发生不同程度的羟基化反应,分别合成二氢杨梅黄酮和二氢栎皮酮。在第三阶段中利用二氢黄酮醇4-还原酶(DFR)的选择性催化特性将上述3种二氢黄酮醇还原为无色且不稳定的花色素,再利用花色苷合酶(ANS)脱水氧化生成有色不稳定的飞燕草素、天竺葵素和矢车菊素这三类花色素[21]。最终通过类黄酮-3-葡糖基转移酶(UFGT)将葡萄糖转移到花色素分子的羟基上生成糖苷键,在对应的糖基转移酶作用下形成花色苷[20]。三类花色苷通过甲基化、酰基化等修饰生成蓝紫色的锦葵素、砖红色的花葵素和紫红色的芍药素。花色苷在内质网中合成后通过囊泡与谷胱甘肽S-转移酶(GST)、多药毒性排除转运体(MATE)、ATP-结合盒(ABC)等转运蛋白运输到液泡中保存[22]。
1.3 花色苷的转录调控 有研究表明,MYB转录因子可以影响花色苷的合成。MdMYB10基因通过诱导下游基因DFR的表达进而正调控果实中花青素的合成,推测MYB10是蔷薇科植物果实呈色的关键调控因子[23]。比如梨(Pyrus pyrifolia)中分离出2个MYB转录因子(PbMYB9和PbMYB10b)并进行功能验证,发现其参与花色苷合成[24]。草莓中发现MYB10基因能促进花色苷含量的积累,并起到正向调控的作用[25]。
在MYB转录因子中,除了MYB相关的转录因子外,还有一段会影响花色苷合成的保守DNA结合区域(MYB结构域)。其中有一个被称为R3-MYB的MYB结构域,R3-MYB一方面通过抑制MBW(MYB-BHLH-WD40复合体)调控花色苷结构基因来阻碍其合成[26],另一方面,对花色苷生成有直接的抑制作用。R3-MYB在秋姬李中会抑制正调控转录因子PSMYB10.1和PSBHLH3对合成花色苷基因PSDFR和PSANS的促进作用,从而阻碍其合成[27]。但是在桃、苹果中发现R2R3-MYB可以直接与MdGSTF6的启动子结合促进花色苷合成[28-29]。
相关研究表明参与花和种皮色素沉着的遗传位点中,有一个基因座编码bHLH转录因子[30],该基因位点对花色苷生物合成的调控作用也十分显著。近几年的研究中,植物中的BHLH33转录因子常与MYB互作,调控结构基因的表达以影响花色苷合成。例如,苹果中的MDBHLH33可与转录抑制基因MDMYB16互作形成异源二聚体,从而减弱MDMYB16对花色苷的抑制作用来保证其正常合成[31]。在桃中,MYB10.1和BHLH3转录因子对合成花色苷也有促进作用[32-33]。
2 花色苷生物合成的影响因素
花色苷由于本身非常不稳定,容易受到温度、光照等外部环境因素的影响[34]。研究表明,温度可以通过影响花色苷合成相关酶的活性间接影响花色苷的合成[35]。在高温环境下,相关酶的活性下降导致花色苷的合成受到抑制,同时花色苷的稳定性遭到破坏,导致花色苷含量大幅下降[36]。
花色苷的合成中光照也具有重要影响,光质、光照强度和照射时间均会影响花色苷的生物合成[37]。研究发现,苹果果实在强光照条件下,花色苷合成相关基因的表达水平和遮光果实相比显著上升,其中MdMYB1基因的表达量增加了20~25倍[38]。此外,在草莓果实中,花色苷合成相关基因的表达量与光照强度呈正相关,强光照条件下果实中花色苷含量显著增加[39]。另外的研究指出,红光和绿光对桃果皮的颜色并无明显影响,白光有微弱促进作用,而蓝光能显著促进桃果皮颜色的提升[40]。
花色苷的合成除了受外部影响以外,还受内部因素的影响。比如糖就是一个影响花色苷合成和色泽的重要因素。田梦瑶等[41]采用了200 mmol/L蔗糖处理采收后的桃果实,发现这种处理能够促进果皮中花色苷含量的积累,不仅能调节相关酶的活性,还促进花色苷相关基因的上调表达。另一方面,贾海峰等[42]研究发现用蔗糖处理成熟期草莓也能提高果实中的可溶性糖和花色苷含量。
3 展望
一些国家已将花色苷类色素开发成为功能性食品和药品,在化妆品、医药等领域具有巨大的应用潜力。不仅如此,花色苷在食品方面的作用也不容忽视。花色苷含量不仅影响果实颜色的变化,还与果实的内在品质密切相关。因此,了解花色苷对果实生長发育的影响,研究果实中花色苷的含量和分布,不仅能够为果实的生产和杂交育种提供理论依据,对提高果实的利用率、产量和品质也具有重要意义。
参考文献
[1]彭祖茂,邓梦雅,严虞虞,等.植物中花青素含量测定及种类分布研究[J].食品研究与开发,2018,39(17):107-111.
[2]Liu S,Zang D,Sun Y,et al.Research advances on biosynthesis pathway of anthocyanins and relevant key enzymes[J].J.Soils Crops,2022,11:336-346.
[3]单建伟,刘晓津,索海翠,等.植物花色苷的研究进展[J].南方农业学报,2019,50(02):278-285.
[4]韦 婷,杨艳宇,李茓艳,等.pH示差法测定苹果皮花色苷含量[J].食品与发酵科技,2022,58(02):130-136.
[5]张 颖,刘璐琪,殷亚蕊,等.桃果肉颜色形成机制研究进展[J].河北果树,2018(06):1-4+10.
[6]李晓娇,刘 超,何春燕,等.粗梗稠李中花色苷的提取及抗氧化和抑菌活性研究[J].北方园艺,2021(23):89-97.
[7]刘馨娜,贾丽娜,苑 宁,等.樱桃的研究进展[J].粮食与油脂,2020,33(04):17-19.
[8]Cappellini F,Marinelli A,Toccaceli M,et al.Anthocyanins:from mechanisms of regulation in plants to health benefits in foods[J].Frontiers in plant science,2021,12.
[9]崔云斌,李嘉钰,刘 钰,等.植物花色苷及其生物合成与药理的研究进展[J].青海草业,2022,31(03):71-75.
[10]江玉婷,秦 昉,陈 洁,等.天然花色苷色素稳定化研究进展[J].食品与机械,2019,35(5):213-218,236.
[11]Ongkowijoyo P,Luna-Vital D A,de Mejia E G.Extraction techniques and analysis of anthocyanins from food sources by mass spectrometry:An update[J].Food chemistry,2018,250:113-126.
[12]聂泽坤.香樟果花色苷的高效提取及其稳态化性质研究[D].淮安:淮阴工学院,2021.
[13]Liu Y,Tikunov Y,Schouten R E,et al.Anthocyanin biosynthesis and degradation mechanisms in Solanaceous vegetables:A review[J].Frontiers in chemistry,2018,6:52.
[14]Gu K D,Wang C K,Hu D G,et al.How do anthocyanins paint our horticultural products[J].Scientia Horticulturae,2019,249:257-262.
[15]Liu H N,Shu Q,Lin-Wang K,et al.DNA methylation reprogramming provides insights into light-induced anthocyanin biosynthesis in red pear[J].Plant Science,2023,326:111 499.
[16]Houghton A,Appelhagen I,Martin C.Natural blues:Structure meets function in anthocyanins[J].Plants,2021,10(4)):726.
[17]邢夢云.杨 梅.FLSs和F3'5'H调控杨梅素生物合成的机制研究[D].杭州:浙江大学,2021.
[18]赵莹,杨欣宇,赵晓丹,等.植物类黄酮化合物生物合成调控研究进展[J].食品工业科技,2021,42(21)):454~463.
[19]Sun L,Huo J,Liu J,et al.Anthocyanins distribution,transcriptional regulation,epigenetic and posttranslational modification in fruits[J].Food Chemistry,2023:135 540.
[20]Colanero S,Perata P,Gonzali S.Whats behind purple tomatoes? Insight into the mechanisms of anthocyanin synthesis in tomato fruits[J].Plant physiology,2020,182(4):1 841-1 853.
[21]Dong T,Han R,Yu J,et al.Anthocyanins accumulation and molecular analysis of correlated genes by metabolome and transcriptome in green and purple asparaguses (Asparagus officinalis, L.)[J].Food chemistry,2019,271:18-28.
[22]李 栋,李 莉,徐艳群,等.植物中花色(下转第7页)(上接第3页)苷转运蛋白研究进展[J].食品安全质量检测学报,2020,11(3):15-20.
[23]梁玉镯,陈新娜,陈东亮,等.MYB转录因子调控植物花青素生物合成研究进展[J].江苏农业科学,2022,50(22):55-64.
[24]Zhai R,Wang Z,Zhang S,et al.Two MYB transcription factors regulate flavonoid biosynthesis in pear fruit (Pyrus bretschneideri Rehd.)[J].Journal of Experimental Botany,2016,67(5):1 275-1 284.
[25]Zhang Z,Shi Y,Ma Y,et al.The strawberry transcription factor FaRAV1 positively regulates anthocyanin accumulation by activation of FaMYB10 and anthocyanin pathway genes[J].Plant biotechnology journal,2020,18(11):2 267-2 279.
[26]Colanero S,Perata P,Gonzali S.The atroviolacea gene encodes an R3-MYB protein repressing anthocyanin synthesis in tomato plants[J].Frontiers in Plant Science,2018,9:830.
[27]方智振,姜翠翠,周丹蓉,等.‘秋姬李PsMYB18基因克隆与功能分析[J].果树学报,2019,36(7):15-23.
[28]牛铁泉,董燕梅,刘海霞,等.葡萄果实myba1与ufgt dfr的作用机制[J].中国农业科学,2018,51(12):149-158.
[29]Cao Y,Xie L,Ma Y,et al.PpMYB15 and PpMYBF1 transcription factors are involved in regulating flavonol biosynthesis in peach fruit[J].Journal of agricultural and food chemistry,2018,67(2): 644-652.
[30]Pal L,Dwivedi V,Gupta S K,et al.Biochemical analysis of anthocyanin and proanthocyanidin and their regulation in determining chickpea flower and seed coat colour[J].Journal of Experimental Botany, 2023,74(1):130-148.
[31]许海峰.bHLH33与MYB抑制子参与苹果花青苷生物合成的分子机理[D].山东:山东农业大学,2020.
[32]Tang T,Zhou H,Wang L, et al.Post-harvest application of methyl jasmonate or prohydrojasmon affects color development and anthocyanins biosynthesis in peach by regulation of sucrose metabolism[J].Frontiers in Nutrition,2022,9:871 467.
[33]Zhang Y,Ling J,Zhou H,et al.1-Methylcyclopropene counteracts ethylene inhibition of anthocyanin accumulation in peach skin after harvest[J].Postharvest Biology and Technology,2022,183:111 737.
[34]Li X,Sheng J,Li Z,et al.Enhanced UV-B radiation induced the proanthocyanidins accumulation in red rice grain of traditional rice cultivars and increased antioxidant capacity in aging mice[J].International Journal of Molecular Sciences,2023,24(4):3 397.
[35]Vukoja J,Pichler A,Kopjar M.Stability of anthocyanins, phenolics and color of tart cherry jams[J].Foods,2019,8(7):255.
[36]高 磊,李 慧,鄭 焕,等.果树中花色苷的生物合成及其调控机制研究进展[J].江苏农业学报,2022,38(1):258-267.
[37]李天来,王 峰,王秀杰,等.光对园艺植物花青素生物合成的调控作用[J].中国农业科学,2020,53(23):188-201.
[38]Takosam,FelixwJ,JacobsR,etal.Light-in ducedex-pressiono faMYB generegulate santhocy an in biosynthesis in red apples[J].PlantPhysiology,2018,142(3):1 216-1 232.
[39] 邵婉璐,李月灵,高 松,等.光照度对成熟红颜草莓果实着色和花青素生物合成的影响及可能的分子机制[J].植物研究,2018(5):661-668.
[40] 池 铭,孙丽娟,马立杰,等.不同光质处理对采后桃果皮色泽及花色苷代谢的影响[J].食品科学,2023,44(3):209-217.
[41] 田梦瑶,周宏胜,唐婷婷,等.外源蔗糖处理对采后桃果皮色泽形成的影响[j].食品科学,2022,43(1):177-183.
[42]Jia H,Wang Y,Sun M,et al.Sucrose functions as a signal involved in the regulation of strawberry fruit development and ripening[J].New Phytologist,2013,198(2):453-465.
