猪肉鲜味物质及其形成机理研究进展
2024-04-08秦凯鹏门小明徐子伟
秦凯鹏,门小明,徐子伟,*
(1.中国计量大学 生命科学学院,浙江 杭州 310018; 2.浙江省农业科学院 畜牧兽医研究所,浙江 杭州 310021)
1 食品鲜味及其感知过程
鲜味(umami)最初由日本学者Ikeda于1908年提出,当时被报道仅具有风味增强作用[1],直到2014年才被认定为基本味觉特征[2]。研究发现,鲜味物质主要通过与口腔味觉受体细胞(TRCs)结合,刺激神经传导至大脑皮层产生味觉感知。目前已经鉴定到6种与鲜味感知相关的特异性G蛋白偶联受体(GPCR)[3-6],其中T1R1/T1R3鲜味异二聚体被证实在识别鲜味感知中起主要作用[7]。如图1所示,当鲜味物质进入口腔后,通过配体-受体特异性结合激活TRCs上的T1R1/T1R3鲜味异二聚体,导致G蛋白亚基发生解离,后者与磷脂酶C-β2(PLC-β2)结合促使磷脂酰肌醇4,5-二磷酸(PIP2)裂解产生肌醇-1,4,5-三磷酸(IP3),从而刺激内质网将Ca2+释放、细胞膜去极化和三磷酸腺苷(ATP)释放[8-9]。ATP作为神经递质刺激传入神经纤维,经面神经和迷走神经等传送到孤束核(NST),进一步到达下丘脑的腹侧,连接到大脑味觉皮层(GC)和岛叶皮层(IC)等区域,最终形成鲜味感知[10-12]。
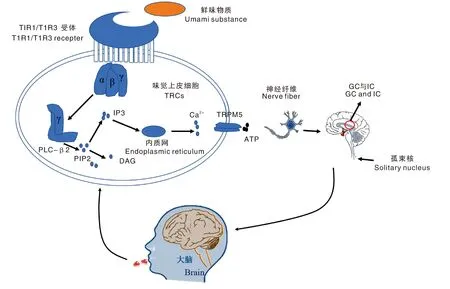
PLC-β2,磷脂酶C-β2;PIP2,磷脂酰肌醇4,5-二磷酸;DAG,三酰甘油;IP3,肌醇-1,4,5-三磷酸;GC,大脑味觉皮层;IC,大脑岛叶皮层;TRPM5,瞬时受体电位阳离子通道亚家族成员5;ATP,三磷酸腺苷。PLC-β2, Phospholipase C-β2; PIP2, Phosphatidylinositol 4, 5-diphosphate; DAG, Triacylglycerol; IP3, Inositol-1,4, 5-triphosphate; GC, Cerebral gustatory cortex; IC, Insular cortex of brain; TRPM5, Transient receptor potential cation channel subfamily M member 5; ATP, Adenosine triphosphate.图1 鲜味感知机制Fig.1 Umami perception mechanism
与众多食物一样,猪肉不仅可以提供丰富的蛋白质营养,还能为消费者带来丰富的味觉体验。其中鲜味是猪肉最明显的味觉特征,也被称为肉味或类肉味。通过提升鲜味特征,能够改善猪肉的综合口感与风味品质[13]。认识鲜味感知过程是人们认识猪肉鲜味特征,进行风味品质评价的重要基础。
2 决定猪肉鲜味特征的物质基础
2.1 游离氨基酸在猪肉鲜味形成中的作用
在可食用新鲜猪肉中,游离氨基酸含量通常在600 mg·kg-1以上[14-15],主要来自活体肌肉蛋白质代谢和宰后肌肉成熟分解过程。这些游离氨基酸通过与味蕾细胞受体结合,可以激活味觉感知传导过程,从而表现呈味活性。有研究发现,不同种类游离氨基酸因侧链基团差异而表现出不同呈味活性特征[16]。对猪肉鲜味形成而言,游离氨基酸的作用可以归纳为以下3种。
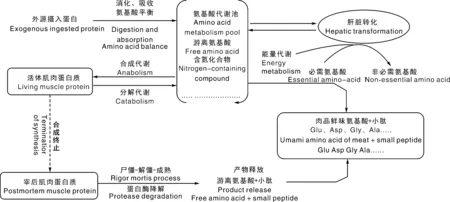
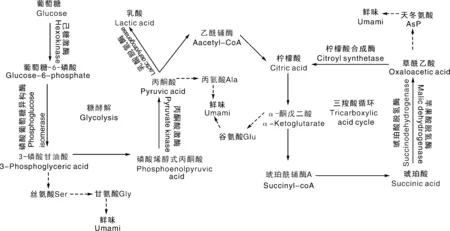
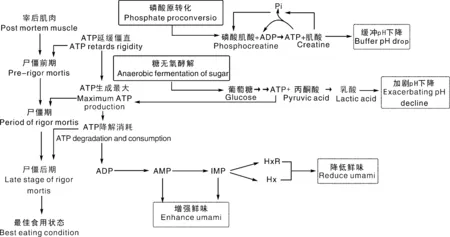
首先是谷氨酸(Glu)和天冬氨酸(Asp)的直接呈味作用。在分子结构上,两者均是具有酸性侧链的L型氨基酸,从而表现酸味和鲜味的组合酸鲜味感,是猪肉中最典型的鲜味氨基酸[17]。如图2所示,Glu与Asp具有相同的特征骨架-O-(C)n-O-,相对容易形成自身环状[18],这是其他编码氨基酸都不具备的。有报道指出,Glu中亲水性基团—COOH与—NH3在弱酸性条件(6 1~5是核糖碳原子位置编号;Glu,谷氨酸;Asp,天冬氨酸;Ala,丙氨酸;Gly,甘氨酸;AMP,腺苷酸;IMP,肌苷酸;GMP,鸟苷酸。1-5 are the position number of the ribocarbon atom; Glu, Glutamate; Asp, Aspartate; Ala, Alanine; Gly, glycine; AMP, Adenosine monophosphate; IMP, Inosine monophosphate; GMP, Guanylic acid.图2 猪肉主要滋味物质化学结构式Fig.2 Chemical structure of main flavor substances of pork 其次是甘氨酸(Gly)和丙氨酸(Ala)等甜味氨基酸的增鲜作用。Gly和Ala是典型的甜味氨基酸,但也具有相对较小的鲜味活性[22]。也有研究表明,猪肉滋味鲜美程度与Glu、Gly含量呈正相关,并且中国本土猪种的Glu和Gly含量高于杂交猪种[23]。在分子结构上,具有短侧链的苏氨酸(Thr)、丝氨酸(Ser)、Ala和Gly同时具有甜味和鲜味组合后的甜鲜味感[24]。但如图2所示,Ala和Gly并不能在分子内形成容易被鲜味受体结合的类似Glu的五元环结构,这可能是Ala和Gly虽具有鲜味活性,但远小于甜味活性的原因之一。此外,鲜味和甜味同样都给人以愉悦满足感,少量甜味对鲜味释放可能具有积极作用[25]。 最后是赖氨酸(Lys)和精氨酸(Arg)等苦味氨基酸的提鲜作用。在分子结构上,具有长、大侧链或碱性侧链的缬氨酸(Val)、亮氨酸(Leu)、异亮氨酸(Ile)、苯丙氨酸(Phe)、酪氨酸(Tyr)、色氨酸(Trp)、组氨酸(His)、Lys和Arg等通常具有苦味特征[24]。有研究指出,含量低于阈值浓度的苦味氨基酸具有增强其他氨基酸呈味特征的作用[26],可能由于苦味氨基酸作用主要取决于侧链基团的疏水作用,与甜味和鲜味氨基酸的亲水作用形成反差对比,导致后者作用得到进一步加强。 核苷酸广泛存在于肉类、食用菌和海鲜类等鲜味特征明显的食品中,目前已发现30余种具有呈味作用的核苷酸类及其衍生物[27-28]。对不同品种猪肉进行研究发现,肌苷酸(IMP)是引起猪肉鲜味最重要的呈味核苷酸,其含量能达到1 500 mg·kg-1以上,与Glu、Asp同为猪肉主体鲜味物质[29-31]。对单一滋味物质而言,刺激味蕾产生味觉特征的最小生理浓度称之为阈值,阈值越低表示该物质呈味活性越强[32]。鸟苷酸(GMP)与腺苷酸(AMP)的阈值分别为125、250 mg·kg—1[30],因此猪肉中GMP与AMP的增鲜作用也不容忽视[31]。综合分析,IMP、AMP和GMP含量可能与中国地方猪的滋味鲜美特征有密切关系,如莱芜猪呈味核苷酸含量高于长白猪、大约克猪[33];青海八眉猪呈味核苷酸含量高于甘肃黑猪和甘肃长白猪[34];东北民猪鲜味核苷酸含量也显著高于杜长大猪[31]。显然,IMP、AMP和GMP这3种核苷酸在猪肉鲜味形成中均发挥了重要作用。 鲜味核苷酸是一类由核糖或脱氧核糖、嘌呤碱基和磷酸基团组成的化合物,磷酸基团取代核糖或脱氧核糖上羟基的碳原子位置不同影响着化合物的呈味特征[29]。如图2所示,3种鲜味核苷酸的核糖结构中1号碳原子羟基均被嘌呤碱基取代,当5号碳原子羟基被磷酸基团取代,即形成5′-核苷酸时具有鲜味,而磷酸基团位于2号和3号碳原子时则不具有鲜味。此外,核苷酸嘌呤环6号碳原子上的亲水基团对鲜味活性有正向积极作用,如IMP和GMP的羰基、AMP的氨基等[35]。了解这些结构特征可以为我们进一步研究揭示猪肉鲜味物质提供参考依据。 肽类呈味作用与自身携带的氨基酸残基及其疏水性有关,通常鲜味肽会含有Asp、Glu或亲水性氨基酸残基,但也有研究表明,Asp与Glu残基会抑制多肽的呈鲜效果或增鲜效果[39-40]。Yu等[41]利用分子模拟手段研究T1R1/T1R3鲜味受体与不同结构的鲜味六肽分子的结合特征,发现鲜味肽产生鲜味的主体部分是其酸性和碱性部分,同时具备正电基团、负电基团与疏水基团,并且连接到对应的感受器上才能产生鲜味。随着相关技术的成熟,鲜味肽将是猪肉鲜味研究的一个重要方向。 不同物质间协同作用是猪肉鲜味产生的重要机制。席斌等[34]发现青海八眉猪、甘肃长白猪与甘肃黑猪的鲜味物质含量差异不大,但八眉猪口感风味表现相对更佳,推测可能是鲜味氨基酸与鲜味核苷酸之间协同作用的原因。其实,早在1971年Yamaguchi等[42]提出的味精当量(EUC)公式:EUC=∑aibi+1 218∑aibi∑ajbj,已经充分体现了鲜味氨基酸和核苷酸之间的协同效应。布丽君等[43]通过分析发现,Glu与Asp之间,氨基酸与核苷酸之间协同作用显著,换算成谷氨酸钠的呈味强度显著高于各鲜味物质呈味强度之和。还有研究发现,GMP是一种比味精更强的增味剂,与谷氨酸钠等量混合时鲜味增加30倍,而与IMP等量混合则增加8倍[44-45];AMP含量在50~100 mg·mL-1时也表现较强的鲜味增强效果[46]。 氨基酸之间的互作效应主要体现为猪肉中的部分氨基酸具增强Glu、Asp鲜味效果的作用,如前文所提到的Lys、Arg等。除此之外,从熟制猪肉中鉴定出的ED、EE、DES等富含谷氨酸的寡肽能在一定程度掩盖苦味[47];将呈鲜味的二肽KG与具有其他味道的4种多肽混合后,呈现出类似牛肉风味强化肽(BMP)的强烈鲜味[48];具有鲜味的EV、AED、SPE、EEN和0.2% IMP混合后鲜味提升显著[49]。研究证明,鲜味寡肽与谷氨酸钠具有协同作用,如鲜味肽FSGLDGSK,当其浓度为2‰时即可提高谷氨酸钠溶液的鲜味[50]。显然,不同物质间的协同效应是鲜味强烈的重要支撑,在考察猪肉鲜味时,应建立综合评价方法来全面分析鲜味物质含量。 蛋白质是构成动物肌肉组织的最主要成分之一。如图3所示,活体肌肉蛋白质代谢状态与宰后肌肉蛋白质降解程度共同决定着肉品游离氨基酸含量组成。对活体肌肉而言,游离态氨基酸和小肽分子主要来自3个方面:1)无论必需氨基酸,还是非必需氨基酸或者小肽分子都可通过消化吸收摄入;2)机体组织蛋白质合成与分解代谢的动态平衡决定着游离氨基酸含量;3)具有鲜味作用的Glu、Asp、Ala和Gly不仅可以通过外源吸收,还可以通过肝转化生成[51-52]。 图3 蛋白质营养代谢与宰后降解Fig.3 Nutrition metabolism and postmortem degradation of proteins 由于氨基酸吸收和利用受摄入氨基酸比例的影响,过多摄入某种氨基酸往往在吸收和转化环节发生改变,并不会直接导致其含量增加[53]。这也是通过添加氨基酸调控猪肉鲜味物质含量,通常并不会收到稳定理想效果的重要原因。有研究发现,一些非必需功能性氨基酸(精氨酸、亮氨酸等)可能通过其他途径(调控骨骼肌肌纤维组成比例)影响猪肉风味品质[54-55]。提高日粮蛋白质利用率、改变肠道微生物菌群等也可以提高肌肉组织的鲜味氨基酸含量[56]。 活体动物的蛋白质消化吸收和营养代谢过程等为宰后肉品游离氨基酸含量组成提供了物质基础。当动物被屠宰后,肌肉蛋白质合成代谢被迫中断,进入单向水解阶段。随着宰后肌肉ATP消耗和pH下降,蛋白酶系统被激活,进而开始分解肌原纤维蛋白,最终完成肌肉成熟嫩化过程,并伴随游离氨基酸、小肽等风味物质形成与释放[57]。宰后肌肉“僵直-解僵-成熟”过程成为影响猪肉中鲜味物质含量的最直接环节。 在活体肌肉中,一些能量代谢中间产物直接参与风味物质代谢过程,甚至本身就是重要的风味物质或其前体物。如图4所示:葡萄糖无氧酵解中间产物3-磷酸甘油酸、丙酮酸,三羧酸循环中的草酰乙酸均是鲜味氨基酸生成的重要前体物;3-磷酸甘油酸是Gly合成前体物;丙酮酸则会通过2个途径分别合成Ala与Glu;草酰乙酸则是Asp合成前体物[58]。因此,增强机体能量有氧代谢途径,对鲜味氨基酸的积累可能具有正向调控作用。 图4 糖酵解与三羧酸循环途径Fig.4 Glycolysis and tricarboxylic acid cycle pathways 不同类型肌纤维及其代谢特征对猪肉品质有重要影响。一致认为,Ⅰ型也被称为慢速氧化型肌纤维,具有较高肌红蛋白含量、较低ATP酶活力与糖酵解潜力,以糖脂有氧氧化代谢为特征,肌纤维直径表现细腻;Ⅱb型又被称为快速酵解型,与Ⅰ型肌纤维恰恰相反,表现出较高的ATP酶活力与糖酵解潜力,以无氧酵解为代谢特征,肌纤维直径相对粗大;Ⅱa与Ⅱx型介于Ⅰ型与Ⅱb型之间,但Ⅱa型氧化代谢能力高于Ⅱx型[59]。由此可见,通过提高猪肉氧化代谢特征的肌纤维类型比例,不仅可以改善猪肉色泽、pH、系水力和嫩度等表观性状,还能富集糖脂氧化代谢产物,关联氨基酸等滋味物质生成,从而提高猪肉综合风味品质。目前关于肌纤维类型与风味物质代谢关系的机理研究较少。 宰后肌肉需要经历尸僵前期、尸僵期和尸僵后期3个阶段,才能成熟为真正意义上的肉品。如图5所示,在尸僵前期,宰后肌肉最初启动磷酸原转化(ATP-CP)途径满足ATP需要,当70%的磷酸肌酸被消耗后启动糖无氧酵解过程[60]。两个过程对比发现,糖无氧酵解过程中丙酮酸向乳酸转化,导致宰后肌肉pH值急剧下降、系水力降低,水溶性滋味物质流失增加[61],而ATP-CP途径则表现相反。当ATP消耗殆尽,宰后肌肉达到最大程度僵直状态,此后进入僵直后期成熟阶段。在成熟过程中,不仅组织蛋白水解生成游离氨基酸,而且ATP降解生成鲜味物质IMP,从而使肉品风味得到改善[50]。 图5 屠宰后IMP生成途径Fig.5 Post-slaughter pathway of IMP generation 如图5所示,ATP在酶作用下被依次分解成二磷酸腺苷(ADP)和AMP,AMP在脱氨酶作用下分解生成IMP。当IMP生成速度大于降解速度时,猪肉IMP含量不断增加,通常在宰后24 h含量达到最高[62]。随着ATP消耗殆尽或IMP合成代谢相关酶失活,IMP将不再增加并开始降低,但IMP也会继续降解成具有苦味的肌苷(HxR)和次黄嘌呤(Hx)[25]。与IMP相比,HxR和Hx在一定程度会缓冲猪肉鲜味。因此在生产实践中,通过抑制IMP降解酶活性可有效降低宰后肉品IMP降解速率,为肉品保鲜起到积极作用[63]。 无论鲜味氨基酸还是鲜味核苷酸的生成途径,都是以能量代谢过程为枢纽。目前普遍接受的调控能量代谢的信号通路有:1)AMPK/SIRT1信号通路。高含量NAD+可以辅助激活SIRT1,从而增加AMPK活性,AMPK通过激活丙酮酸激酶来调节糖酵解[64]。丙酮酸作为Glu与Ala合成的重要前体物,其含量变化可能会影响鲜味氨基酸生成。2)PI3K/Akt信号通路。激活后的磷脂酰肌醇三磷酸酶(PI3K)通过级联反应激活丝氨酸/苏氨酸激酶(AKT)。活化的AKT可以调节GSK3β进而增强糖原合酶活性促进糖原生成;还可以通过磷酸化叉头框蛋白O1(FOXO1),致使其失活以抑制糖异生基因表达,从而降低非糖物质向葡萄糖转化的速率,抑制机体能量代谢[65]。 有研究发现,UCP基因的表达可以增强氧化磷酸化速率而ATP产量不变,UCP基因在骨骼肌中的高表达会导致能量消耗增多,但IMP合成前体物含量不变,进而影响IMP的积累[66]。AMPD1也可在骨骼肌中高水平表达,其产物腺苷单磷酸脱氨酶是IMP合成途径中的关键限速酶;鸡AMPD1基因与肌苷酸含量存在强相关性[67]。无论是对能量代谢的源头还是对中间产物的调控,都会影响鲜味物质合成的重要前体物含量,直接影响鲜味物质的积累。 此外,国内外研究发现了一系列在肌纤维类型转化中发挥调控作用的细胞信号通路与因子,如CaN、AMPK、Wnt、PGC1α、MEF2、FOXO1和MRFs等可能对猪肉风味物质形成具有调控作用[68]。上述研究报道为优质风味猪育种与分子营养调控奠定了基础,值得后续深入探索研究。 在了解猪肉鲜味感知过程基础上,通过综述猪肉鲜味特征形成物质基础的国内外研究进展,阐明了鲜味氨基酸、鲜味核苷酸与鲜味肽是猪肉鲜味呈现的重要物质基础,其中鲜味肽的作用越来越受到研究关注,不同鲜味物质间通过协同效应可以进一步增强鲜味活性。肌肉氨基酸代谢、能量代谢和肌纤维类型构成等是决定猪肉鲜味物质生成的重要因素,相关控制基因与分子通路还需要深入研究探索,以期为猪肉风味品质改良调控提供新思路和科学依据。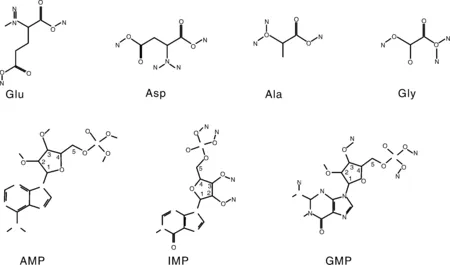
2.2 核苷酸在猪肉鲜味形成中的作用
2.3 呈味肽在猪肉鲜味形成中的作用

2.4 不同滋味物质的互作效应
3 影响猪肉鲜味物质生成途径
3.1 蛋白质营养代谢与宰后降解
3.2 活体肌肉能量代谢与肌纤维类型
3.3 宰后肌肉ATP生成转化与核苷酸降解
4 影响猪肉鲜味物质生成的分子机制探讨
5 小结与展望
