miR-27b对心力衰竭大鼠心肌细胞自噬、心肌能量代谢及PPARα信号的作用机制
2024-04-08陈运起吴钟伟赵圣吉李堪董王敢钟江华
陈运起 吴钟伟 赵圣吉 李堪董 王敢 钟江华
(1海南西部中心医院心血管内科,海南 儋州 571700;2中南大学湘雅医学院附属海口医院心血管内科)
相关报道显示,心力衰竭患者5年死亡率约高达50%〔1〕。心肌细胞能量代谢紊乱在该病中发挥着重要作用。正常情况下,心脏活动所需能量几乎完全靠有氧代谢提供,葡萄糖和脂肪酸等物质是心肌细胞代谢的重要能量底物,其中60%~80%的心肌能量供应来自脂肪代谢。心力衰竭过程中,微循环障碍导致心肌缺血、缺氧,脂肪酸氧化受损,能量代谢从优先利用脂肪酸为主转变为优先利用葡萄糖的无氧酵解,从而导致三磷酸腺苷(ATP)产生减少,以糖酵解为主的供能模式明显增加乳酸产生,使脂肪酸利用减少,造成脂质在心脏沉积,加重心肌损害,同时线粒体生物合成减少,ATP生成下降,导致心肌细胞自噬紊乱,从而增加心肌细胞凋亡,加之心力衰竭时线粒体生物合成减少,能量生成进一步下降,心功能继之恶化,因此又加速了心力衰竭进程〔2~4〕。磷酸腺苷活化蛋白激酶(AMPK)具有调控细胞能量代谢作用,主要通过调控其下游因子过氧化物酶体增殖物激活受体(PPAR)α促进能量产生,维持细胞能量平衡〔5〕。心肌细胞中的PPARα呈现高表达,其与多种心血管疾病联系密切〔6~7〕。另外研究发现〔8〕,AMPK/PPARα通路参与心肌代谢过程,并对心力衰竭具有一定保护作用。目前,关于心力衰竭如何有效治疗已成为心血管疾病研究者的研究重点,因此,寻找一种安全有效的治疗方法至关重要。miR-27b是miRNAs家族成员之一,与多种肿瘤及心脏相关疾病的产生及发展联系密切。研究发现〔9〕,在心肌炎中,沉默miR-27b表达后可抑制心肌细胞凋亡。但目前miR-27b与心力衰竭的相关研究较少,具体机制尚不明确,因此本实验研究miR-27b对心力衰竭大鼠心肌细胞自噬、心肌能量代谢及PPARα信号的作用机制。
1 材料和方法
1.1实验动物 SPF级SD健康雄性大鼠40只,体质量200~300 g,购于北京维通达生物技术有限公司〔许可证号:SYXK(京)2019-0428〕,于医院实验室适应饲养1 w。本实验已经过医院伦理委员会审批(批号:Y2019-009-14)。
1.2主要实验试剂与仪器 miR-27b拮抗剂及阴性对照(上海吉玛制药技术公司),苏木素-伊红(HE)和Masson染色液(南昌雨露实验器材有限公司),miR-27b、AMPK、PPARα引物合成(北京雷根生物技术公司),TUNEL工作液(北京嘉美诺斯生物科技公司),微管相关轻链蛋白(LC)3Ⅱ、LC3Ⅰ、自噬相关蛋白(Beclin)-1、组织蛋白酶(Cath)D一抗(上海信裕生物工程有限公司),羊抗兔二抗(北京百奥莱博科技公司)。
1.3分组、模型建立及干预 40只大鼠随机分为健康组、模型组、miR-27b-NC组及miR-27b组,每组10只。除健康组外,其余3组建立心力衰竭模型:戊巴比妥钠麻醉后备皮,暴露腹腔并游离出主动脉,在主动脉旁置入7号针后,用丝线分离出主动脉与7号针,结扎后拔出针,缩窄65%左右的腹主动脉,缝合。术后予青霉素腹腔注射5 d,预防感染。造模期间意外死亡3只大鼠,模型组、miR-27b-NC组及miR-27b组为每组9只大鼠。建模成功标准见病理检查结果。大鼠建模成功24 h后,健康组、模型组大鼠均于腹主动脉注射5 μl生理盐水,miR-27b-NC组及miR-27b组于腹主动脉分别注射5 μl(20 nmol/L)拮抗剂阴性对照、5 μl(20 nmol/L)miR-27b拮抗剂,1次/d,共4 w。
1.4取材 建模2 w后,每组取3只大鼠,采用磷酸盐缓冲液(PBS)经腹主动脉灌流,取大鼠心脏组织标本,纵向切取左室0.3~0.5 cm的心肌组织,于液氮中保存。将剩余部分心肌组织于多聚甲醛中固定,后行HE染色及Masson染色观察病理学形态。治疗结束后,麻醉大鼠,暴露心脏,采集心尖部血液5 ml,置于-80 ℃冰箱中待检,后按照上述方法取大鼠心肌组织备用。
1.5HE及Masson染色观察心肌组织病理形态及纤维化 取各组于多聚甲醛中固定的心肌组织,石蜡包埋、切片,行HE和Masson染色,显微镜下观察心肌组织病理学改变和胶原纤维化情况。
1.6实时荧光定量-聚合酶链反应(RT-PCR)检测miR-27b、AMPK、PPARα mRNA指标水平 取各组心肌组织,剪碎、研磨、离心后,Trizol提取总RNA。根据cDNA 第一链合成试剂盒说明,进行逆转录合成 cDNA。PCR条件:预变性95 ℃,1 min;94 ℃,30 s;54 ℃,30 s;72 ℃,1 min。共40个循,72 ℃ 5 min,以β-actin作为内参。基因相对表达量=2-ΔΔCt。引物序列:miR-27b正向:5′-GGGGTTCACAGTGGCTAAG-3′,反向:5′-CAGTGCG TGTCGTGGAGT-3′(118 bp);AMPK正向:5′-TCGACAGAAGA TTCGGAGCC-3′,反向:5′-CTTGGAGTTCGAAAAG TCCGT-3′(166 bp);PPARα正向:5′-AAGCCATCTTCACGATGCTG-3′,反向:5′-TCAGAGGTCCCTGAACAGTG-3′(117 bp),β-actin正向:5′-GCGAGAA GATGACCCAGAT-3′,反向:5′-CCAGTGGTACGGCCAGAGG-3′(528 bp)。
1.7ELISA法检测心肌细胞能量代谢指标 取各组大鼠心尖部血液2 ml,2 000 r/min离心15 min,取上清,于-80 ℃冰箱中待检。采用ELISA检测血清N末端B型脑钠肽前体(NT-proBNP)和游离脂肪酸 (FFA)水平。实验严格按照试剂盒说明书。
1.8TUNEL检测心肌细胞凋亡 取各组心肌组织石蜡切片,根据TUNEL试剂盒说明处理,镜下观察并拍摄图片,二氨基联苯胺(DAB)显色,镜下观察并计数阳性细胞。
1.9免疫印迹检测自噬相关蛋白LC3Ⅱ、LC3Ⅰ、Beclin-1、CathD表达 取各组心肌组织剪碎后研磨,裂解液裂解,匀浆机匀浆,离心5 min,3 000 r/min,离心半径为2 cm,取上清,辛可宁酸(BCA)法测定蛋白总浓度,将蛋白煮沸后取20 μl蛋白进行电泳后转移至聚偏氟乙烯(PVDF)膜上,脱脂奶粉封闭2 h,加入LC3Ⅱ、LC3Ⅰ、Beclin-1、CathD一抗(1∶500) 孵育过夜,PBS清洗,加入羊抗兔二抗(1∶1 000)孵育2 h,PBS清洗,增强化学发光法显色,凝胶成像仪中观察并摄像,以 ImageJ 分析各组蛋白相对表达。
1.10统计学分析 采用SPSS22.0软件进行单因素方差分析、SNK-q法或t检验。
2 结 果
2.1HE染色及Masson染色观察结果 HE染色:健康组心肌细胞形态正常,结构清晰;模型组与miR-27b-NC组心肌细胞水肿并有大量炎性细胞浸润,此外产生大量成纤维细胞;miR-27b组与模型组相比,成纤维细胞增生及炎性细胞浸润减少。Masson染色:胶原纤维呈现蓝紫色,心肌细胞及肌纤维呈现红色。健康组心肌组织结构正常,心腔附近有少量胶原纤维;模型组与miR-27b-NC组胶原纤维显著增加;与模型组相比,miR-27b组胶原纤维增生有所减少,见图1。上述结果提示建模成功。

图1 各组心肌组织病理
2.2各组心肌细胞凋亡比较 健康组细胞核呈现蓝色染色,边界清晰。模型组与miR-27b-NC组可见大量棕色染色,细胞核固缩;与模型组相比,miR-27b组深染棕色细胞核减少,见图2。
2.3各组心肌组织中miR-27b、AMPK、PPARα mRNA表达比较 与健康组相比,其余3组心肌组织中miR-27b表达显著升高,AMPK mRNA、PPARα mRNA表达显著降低(P<0.05);模型组与miR-27b-NC组心肌组织中miR-27b、AMPK mRNA、PPARα mRNA表达对比无统计学差异(P>0.05);与模型组和miR-27b-NC组相比,miR-27b组心肌组织中miR-27b表达显著降低,AMPK、PPARα mRNA表达显著升高(P<0.05),见表1。

表1 各组miR-27b、AMPK、PPARα mRNA表达及NT-proBNP、FFA水平对比
2.4各组细胞能量代谢指标比较 与健康组相比,其余3组血清中NT-proBNP、FFA水平显著升高(P<0.05);模型组与miR-27b-NC组血清中NT-proBNP、FFA水平对比无统计学差异(P>0.05);与模型组和miR-27b-NC组相比,miR-27b组NT-proBNP、FFA水平显著降低(P<0.05),见表1。
2.5各组心肌组织中LC3Ⅱ、LC3Ⅰ、Beclin-1、CathD蛋白表达比较 与健康组相比,其余3组心肌组织中LC3Ⅰ水平显著降低,LC3Ⅱ、Beclin-1、CathD水平显著升高(P<0.05);模型组与miR-27b-NC组心肌组织中LC3Ⅱ、LC3Ⅰ、Beclin-1、CathD水平对比无统计学差异(P>0.05);与模型组和miR-27b-NC组相比,miR-27b组心肌组织中LC3Ⅰ水平显著升高,LC3Ⅱ、Beclin-1、CathD水平显著降低(P<0.05),见表2、图3。
表2 各组LC3Ⅱ、LC3Ⅰ、Beclin-1、CathD蛋白表达比较
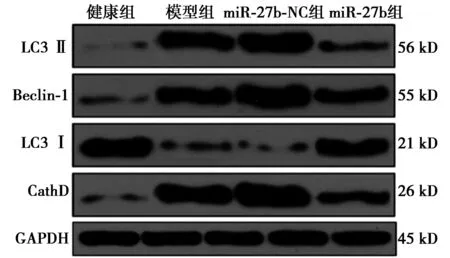
图3 各组LC3Ⅱ、Ⅰ、Beclin-1、CathD蛋白表达
3 讨 论
心肌细胞自噬在心力衰竭中发挥重要作用,心肌细胞自噬的形成与降解发生异常,使自噬体异常堆积,导致细胞凋亡〔10〕。自噬主要由细胞内缺氧、应激等因素诱发。自噬本是机体的正常生理应激反应,但在多种疾病发展中,细胞异常自噬发挥了重要作用。研究显示〔11〕,自噬会减少心肌细胞数量,促进心力衰竭发展。LC3Ⅰ、Ⅱ、Beclin-1 和CathD是与细胞自噬密切相关的蛋白。Beclin-1 和 CathD在正常细胞中表达量极低,而在发生自噬的细胞中表达量明显增高;LC3Ⅰ、Ⅱ是LC3的两种类型,在自噬发生后,LC3Ⅰ与相关蛋白结合后形成LC3Ⅱ,可反应自噬的活性。miR-27b与自噬具有密切的联系。研究表明〔12〕,抑制miR-27b可降低心肌细胞凋亡率。另外〔13〕,miR-27b与炎症反应、细胞凋亡过程有关。且在张韩等〔14〕研究中已经证明,沉默miR-27b表达后,通过抑制B细胞淋巴瘤基因(Bcl)-2腺病毒E/B19 kD相互作用蛋白(BNIP3)水平,激活Wnt/β-连环蛋白(catenin)通路,抑制自噬过程。本实验结果提示低表达的miR-27b具有抑制心肌细胞凋亡及自噬的作用。
心肌正常的能量代谢是心脏功能稳定的基础。在心力衰竭产生后,心肌组织能量代谢发生紊乱,导致心肌的能量供应不足。正常心肌活动是由FA和葡萄糖氧化代谢产生的ATP供能,其中FA是心肌能量代谢的主要来源;心力衰竭时FA氧化降低,血清FFA水平明显升高,FFA能损伤线粒体功能和呼吸链,抑制ATP的生成,使心肌细胞的能量产生和利用均出现障碍,加剧了心功能的恶化〔15〕。NT-proBNP参与心室重构过程,是心力衰竭的诊断指标。研究提示〔16〕,异常高表达的NT-proBNP水平提示心肌损伤严重,心肌能量代谢差。PPARα是AMPK通路的下游靶分子,对维持心肌能量代谢平衡具有一定作用。AMPK通过上调PPARα表达增加心肌线粒体脂肪酶活性,促进ATP产生,加强FFA氧化代谢及利用率,改善心力衰竭〔17〕。心肌能量需求中FFA占据70%左右,葡萄糖占据近30%,心力衰竭产生后会导致葡萄糖代谢增加,FFA的利用减少,影响线粒体功能,抑制糖酵解,导致心肌能量不足发生重构,促进心功能的恶化〔18〕。何远利等〔19〕研究提示,心力衰竭宁合剂通过调控AMPK/PPARα通路,抑制FFA水平表达,改善心力衰竭大鼠心肌能量代谢。沈林園〔20〕研究结果提示,抑制miR-27b水平后,能量代谢水平标记基因AMPK的表达量也显著降低,表明miR-23a和miR-27b可以显著地降低细胞的能量代谢水平。本实验结果与上述结论相似,提示抑制miR-27b水平后,通过调控AMPK/PPARα通路改善心肌能量代谢水平,从而起到保护心功能的作用。
综上所述,抑制miR-27b表达后可抑制心力衰竭大鼠心肌细胞自噬,减少心肌细胞凋亡,同时通过调控AMPK/PPARα通路改善心肌能量代谢水平,从而起到心保护的作用。
