植物乳植杆菌类胡萝卜素生物合成关键基因LpispA的功能鉴定
2024-04-03李舒丽谢卓婷沈润泽郭丽琼林俊芳陈谋通叶志伟
李舒丽,谢卓婷,沈润泽,郭丽琼,2,林俊芳,2,陈谋通,叶志伟,2*
(1.华南农业大学食品学院,广东广州 510642)
(2.广东省微生态制剂工程技术研究中心,广东广州 510642)
(3.广东省科学院微生物研究所,华南应用微生物国家重点实验室,广东省微生物安全与健康重点实验室,农业农村部农业微生物组学与精准应用重点实验室,广东广州 510070)
类胡萝卜素(Carotenoids)是由高等植物、微生物和藻类等合成的一大类类异戊二烯代谢物[1-3]。迄今为止,已在自然资源中发现了1 200 多种类胡萝卜素,包括烃类的胡萝卜素和含氧的叶黄素[4,5]。类胡萝卜素具有抗衰老和抗肿瘤等多种活性功能,适量摄入该化合物可以有效防止机体细胞和组织受到氧化损伤,提高机体免疫系统能力[6-8]。目前,类胡萝卜素已被广泛应用于医药保健、食品着色、饲料营养补充和化妆品生产等多个领域[9]。据美国商务通讯公司(Business Communications Company,BCC)的市场调研报告称,商业应用类胡萝卜素的全球市场价值预计到2027 年将突破27 亿美元关口,2022 年至2027 年的复合年增长率为5.7%[10]。
随着生物功能技术的发展和人们食品安全意识的提高,相对于植物提取法和化学合成法,人们更青睐于通过合成生物学等手段构建微生物细胞工厂生产的新型和高安全性类胡萝卜素。研究显示,具有“公认安全”(Generally Regard As Safe,GRAS)地位的植物乳植杆菌(Lactiplantibacillus plantarum)在其代谢过程中能产生C30类胡萝卜素、细菌素、胞外多糖和叶酸等多种有益物质[11-13]。各种类胡萝卜素的生物合成途径是了解生物的次生代谢演变的一个典型系统[14]。尽管目前已鉴定出各种C30类胡萝卜素,但由于缺乏组学数据和胡萝卜素基因的功能鉴定从而限制了对植物乳植杆菌甚至乳酸菌的类胡萝卜素合成途径的研究。法尼基焦磷酸(Farnesyl Pyrophosphate,FPP)和牻牛儿基牻牛儿焦磷酸(Geranylgeranyl Pyrophosphate,GGPP)分别是C30和C40类胡萝卜素的直接前体,Matthews等[15]和Lois 等[16]研究发现,前体物质的合成是类胡萝卜素合成代谢途径中的限速步骤甚至是“瓶颈”。FPP 是由FPP 合成酶(FPP Synthase,ispA)催化异戊烯基焦磷酸(Isopentenyl Pyrophosphate,IPP)和二甲丙烯基焦磷酸(Dimethylallyl Pyrophosphate,DMAPP)缩合而成的,再经GGPP 合成酶(GGPP Synthase,crtE)进一步催化形成GGPP。因此,前体物质合成酶性质的研究对深入解析类胡萝卜素合成的分子机理和调控机制都至关重要。
项目组Ye 等[17]的前期转录组分析表明,海洋源植物乳植杆菌菌株Lp10 含有类胡萝卜素生物合成的基因簇。本研究对植物乳植杆菌FPP 合酶基因LpispA进行了克隆、绘制系统进化树和构建类胡萝卜素合成颜色功能互补的大肠杆菌表达系统pAC-BETA Δ E-ispA,利用紫外可见光谱(Ultraviolet-Visible Spectroscopy,UV-Vis)和反相高效液相色谱(Reversed-Phase High Performance Liquid Chromatography,RP-HPLC)检测产物成分来验证基因的功能,为探究植物乳植杆菌类胡萝卜素生物合成机制及为高效合成新的类胡萝卜素奠定理论基础,有望进一步推进应用植物乳植杆菌作为类胡萝卜素补充剂。
1 材料与方法
1.1 材料
1.1.1 菌株和质粒
植物乳植杆菌Lp10 菌株由所在研究组实验室从海带中分离纯化并保存;质粒pAC-BETA 由所在研究组保藏;大肠杆菌DH5α购买于大连TaKaRa公司。
1.1.2 主要的试剂和仪器
细菌基因组DNA 提取试剂盒购买于TIANGEN公司;ClonExpress II 一步克隆试剂盒和Phanta Max超保真DNA 聚合酶购买于Vazyme 公司;限制性内切酶SpeI、NdeI 购买于大连Takara 公司。
高效液相色谱仪LC-2030(日本,株式会社岛津制作所);紫外可见分光光度计UV-2802PC(上海,尤尼科);高速冷冻离心机5804R(德国,Eppendorf);MJ Mini PCR 仪(美国,BIO-RAD);GelDoCTMXR +凝胶快速成像系统(美国,BIORAD)。
1.1.3 引物
本文所用引物如表1 所示。
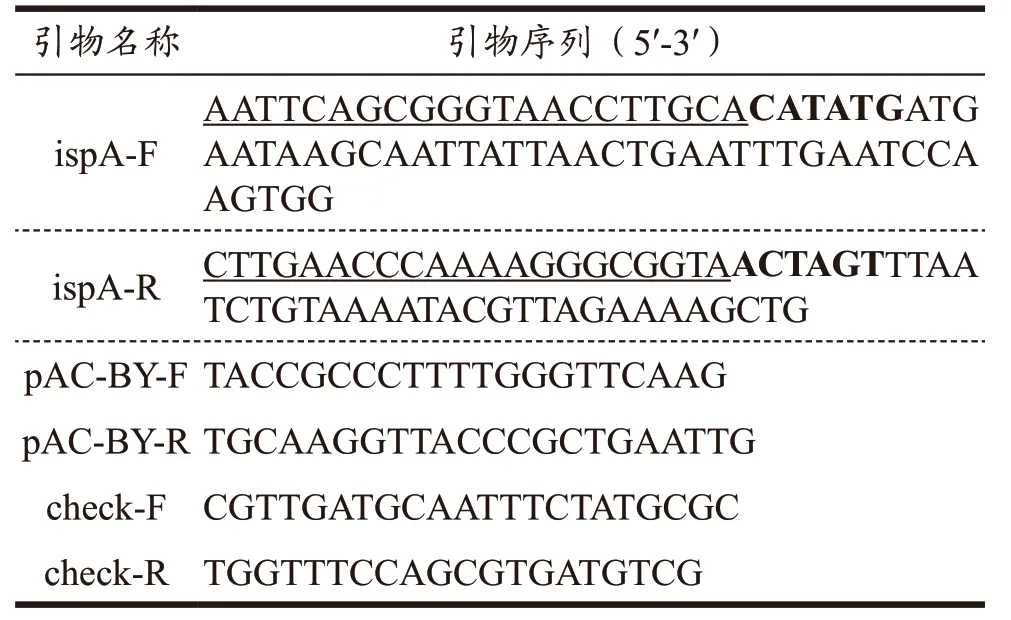
表1 基因扩增的引物序列Table 1 Primer sequences for gene amplification
1.2 实验方法
1.2.1 目的基因扩增
以试剂盒提取的植物乳植杆菌Lp10 菌株的基因组DNA 为模板,设计引物ispA-F&R(表1),扩增LpispA基因的开放阅读框(Open Reading Frame,ORF)序列。反应体系(50 μL):模板200 ng,上下游引物各2 μL(0.4 μmol/L),Phanta Max 1 μL,dNTPs 1 μL,2×PCR buffer 25 μL,加超纯水至50 μL。扩增条件:95 ℃ 3 min;95 ℃ 15 s,60 ℃ 15 s,72 ℃30 s,30 个循环;4 ℃保存。以质粒pAC-BETA 为模板,设计引物pAC-IBY-F&R,反向扩增线性化片段pAC-IBY,反应体系与上述LpispA基因扩增相同;扩增条件:95 ℃ 30 s;95 ℃ 15 s,59 ℃ 15 s,72 ℃5 min,30 个循环;4 ℃保存。PCR 产物用1%(m/V)琼脂糖凝胶电泳检测,切下目的条带并使用琼脂糖凝胶DNA 纯化回收试剂盒(MAGEN)回收目的片段并进行测序比对。
1.2.2 ispA系统进化树的构建
用MEGA 软件(7.0)对本研究中获得的植物乳植杆菌LpispA基因推断的氨基酸序列以及从GenBank 检索的相关物种(包括植物、藻类、细菌和蓝细菌)的氨基酸序列进行比对,采取邻接法(Neighbor-Joining,NJ)构建系统进化树。
1.2.3LpispA基因功能互补表达载体的构建
植物乳植杆菌LpispA基因的功能互补表达质粒pAC-BETAΔE-ispA 的构建路线如图1 所示。用ClonExpress II 一步克隆试剂盒将目的片段LpispA和pAC-IBY 连接,然后转化至大肠杆菌DH5α感受态细胞中,于含氯霉素 (30 μg/mL)的LB(10 g 胰蛋白胨,5 g 酵母提取物,10 g 氯化钠,1 L 超纯水,pH 值7.0)平板中37 ℃培养过夜,用引物check-F&R进行PCR 验证。挑选拟转化子并于含30 μg/mL 氯霉素的LB 液体培养基中37 ℃、200 r/min 下扩大培养,提取质粒进行双酶切(SpeI 和NdeI)检验,并送至公司测序分析。将正确的重组质粒命名为pACBETAΔE-ispA。
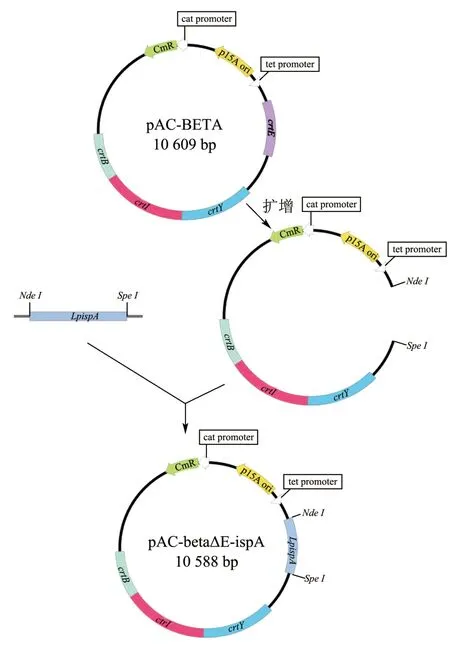
图1 重组载体pAC-BETAΔE-ispA 的构建路线Fig.1 Structure route of recombinant vector pAC-BETAΔE-ispA
1.2.4 转化子色素的提取
参照叶志伟[18]的方法,将含有质粒pAC-BETA、pAC-BETA Δ E-ispA 和空载体的大肠杆菌工程菌株分别接种于LB 液体培养基,37 ℃、200 r/min 黑暗培养36 h。每个样品取两份培养物各50 mL,一份用于转化子色素的提取,具体操作如下:a)将50 mL发酵液于4 ℃,10 000 r/min 离心5 min,弃上清液;b)用超纯水重新悬浮沉淀的大肠杆菌细胞,短暂离心弃上清液;c)加入2 mL 丙酮,涡旋振荡分散沉淀物;d)在55 ℃水浴中处理15 min,每隔5 min 振荡一次;e)4 ℃,10 000 r/min 离心20 min 后,收集含有色素的上清液;f)离心所得的细胞沉淀物再按步骤c~e 处理一次;g)收集所有上清液并用丙酮定容至5 mL,-80 ℃保存(类胡萝卜素易光解,色素提取过程中应避光处理)。另一份用于测定细胞干重,具体操作如下:①重复以上步骤a~b;②将细胞在65 ℃下避光干燥处理至恒重(约24 h),称重。
1.2.5 色素提取物的UV-Vis和RP-HPLC分析
使用UV-Vis 波长扫描提取液以获得350~800 nm光谱范围内的光吸收情况。确定最大吸收波长后,参照Inbaraj 等[19]的方法,通过RP-HPLC 对各色素溶液进行定性和定量。色谱条件为:色谱柱,J&K Scientific C18反相柱(250 mm×4.6 mm,5.0 μm);流动相,甲醇:乙腈: CH2Cl2=85:12:3(V/V/V)(含0.4 g/L抗坏血酸);流速,1 mL/min;柱温,30 ℃;进样量,20 μL。分析过程中使用标准加入法,即在各样品中加入一定量的β-胡萝卜素标准品(Solarbio)对提取液进行目的物质β-胡萝卜素的定性,利用β-胡萝卜素标准品试验绘制的校准曲线进行定量分析。
1.2.6 数据分析
根据样品中类胡萝卜素的质量浓度和细胞干重,计算每克细胞干重的β-胡萝卜素含量(mg/g DCW)。每个样品设置3 个平行,实验数据采用平均数±标准差表示。使用Graphpad Prism 8.0.2 软件构建图形,当P<0.05 时为显著性差异。
2 结果与讨论
2.1 LpispA基因和pAC-IBY片段克隆及纯化
对植物乳植杆菌Lp10 的LpispA基因和质粒pAC-BETA 中的pAC-IBY 片段进行克隆及纯化,琼脂糖凝胶电泳结果如图2 所示,可发现前者在800~1 200 bp 处有显著的单一条带,与LpispA基因ORF 序列891 bp 预期大小一致(图2a),后者在10 000 bp 左右处有明显的单一条带,与pACIBY 片段9 685 bp 预期大小一致(图2b)。扩增产物的测序结果表明,LpispA基因ORF 序列和pACIBY 片段均已被成功克隆。
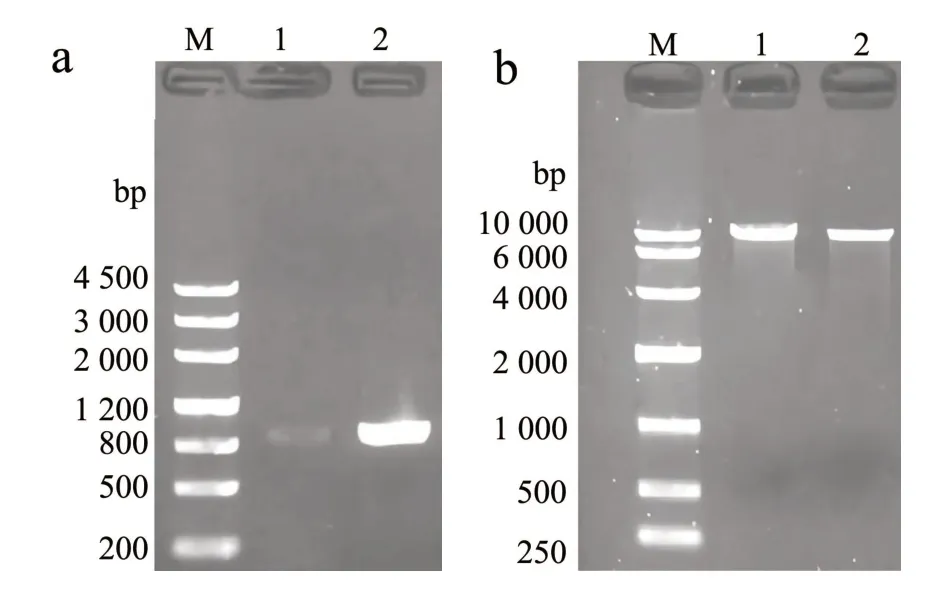
图2 LpispA 和pAC-IBY 的PCR 扩增Fig.2 PCR amplification for LpispA and pAC-IBY
2.2 LpispA的系统发育分析
利用植物、藻类、细菌和蓝细菌等4 个类群的氨基酸序列构建LpispA 系统进化树(图3)。系统进化树中,红色三角形标记显示了植物乳植杆菌LpispA 在系统树上的位置,其它物种氨基酸序列的GenBank 登陆号显示在相应每个物种名称后面。系统进化树表明植物乳植杆菌与桃色欧文氏菌(Erwinia persicina)遗传距离最近,经过逐层汇聚,相对与藻类和植物等遗传距离较远。表明在所选的物种中,植物乳植杆菌与细菌分类中的桃色欧文氏菌亲缘关系最近,与蓝细菌次之,与藻类和植物的亲缘关系最远。

图3 基于FPP 合成酶(ispA)氨基酸序列NJ 法构建的系统进化树Fig.3 Phylogenetic tree analysis of farnesyl pyrophosphate synthase (ispA) based on the amino acid sequences by the neighbor-joining method
2.3 pAC-BETAΔE-ispA重组载体的构建
对构建的重组载体pAC-BETAΔE-ispA 进行双酶切鉴定,结果如图4 所示,酶切出两条条带,亮度清晰,大小分别与线性化载体pAC-IBY 和目的基因LpispA一致。对重组质粒进行测序分析鉴定,结果显示重组载体pAC-BETAΔE-ispA被成功构建。
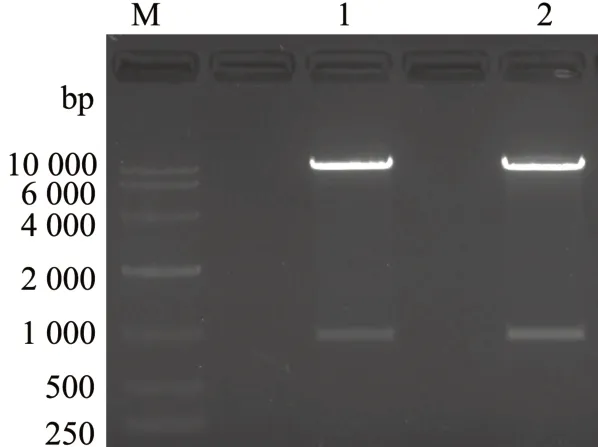
图4 pAC-BETAΔE-ispA 重组载体双酶切验证Fig.4 Identification of recombinant vector pAC-BETAΔEispA by double enzyme digestion
2.4 转化子类胡萝卜素含量的测定
低温离心收集含质粒pAC-BETA(阳性对照)、pAC-BETAΔE-ispA 和空载体(阴性对照)构建的大肠杆菌工程菌株的细胞(图5a),再用丙酮提取菌体中的色素(图5b)。其中,阳性对照含有源自细菌的β-胡萝卜素生物合成基因簇的质粒,菌体β-胡萝卜素沉着,细胞呈现橙黄色。结果显示,构建的LpispA基因功能互补表达工程菌株及其提取的色素颜色皆显浅橙黄色,对比阳性对照颜色稍浅,但深于阴性对照(图5)。对提取到的色素进行UV-Vis 波长扫描测量,转化子与β- 胡萝卜素标准品的色素样品皆呈现类胡萝卜素紫外可见光吸收图谱的典型三指吸收峰结构,其最大吸光度在450 nm(图6a)。利用RP-HPLC 进行定性定量分析,结果显示,pAC-BETAΔE-ispA 组、β-胡萝卜素标准品组以及pAC-BETAΔE-ispA+组(添加了β-胡萝卜素标准品)均在保留时间43.90 min 下有相似的色谱峰出现,说明pAC-BETAΔE-ispA 组细胞合成了β-胡萝卜素(图6b)。由β-胡萝卜素标准品制备的校准曲线进行定量,结果显示转化子pACBETA,pAC-BETAΔE-ispA 的β- 胡萝卜素含量分别为0.80 mg/g DCW 和0.05 mg/g DCW,差异显著(P<0.05)(图7)。实验组的β-胡萝卜素含量较对照组低,推测可能的原因是对照组pAC-BETA质粒的类胡萝卜素功能酶基因来源于单一物种,而实验组pAC-BETAΔE-ispA 为重组质粒,其中来源于不同物种的类胡萝卜素功能基因组合在异源表达代谢通路中,容易出现基因表达和酶活性失衡[20],不平衡的碳通量易导致中间产物的积累,从而降低系统合成效率[21],而且可能产生对宿主生长不利的中间产物[22]。

图5 转化子和提取色素的颜色分析Fig.5 Colors analysis of transformants and extracted pigments

图6 色素提取物的UV-Vis 和RP-HPLC 分析图谱Fig.6 Chromatogram of pigment extracts by UV-Vis and RP-HPLC

图7 转化子的β-胡萝卜素含量Fig.7 β-carotene content of transformants
2.5 细菌类胡萝卜素的潜在生物合成途径
质粒pAC-BETA 含有来自草生欧文氏杆菌的β-胡萝卜素生物合成基因簇,包括GGPP 合成酶基因crtE、番茄红素环化酶基因crtY、八氢番茄红素脱氢酶基因crtI和八氢番茄红素合成酶基因crtB。含有该质粒的大肠杆菌培养物可通过β-胡萝卜素的积累而显现黄色表型[23]。依据以上实验结果,植物乳植杆菌FPP 合成酶基因LpispA在构建的大肠杆菌功能互补表达系统中成功表达并使宿主细胞产生β-胡萝卜素,实现了对草生欧文氏杆菌GGPP 合成酶基因crtE的功能替换,为类胡萝卜素合成多功能酶,实现在C40类胡萝卜素合成路径中替换crtE的缺失。Armstrong 等[24]研究发现,ispA 与crtE 的氨基酸序列在系统发育分析上具有较高的同源性。Miguel 等[25]研究表明,玉米胚乳中的FPP 合成酶具有GGPP 合成酶的功能活性,既影响FPP 的产量,也对GGPP 形成产生作用。通过定位突变等处理改变微生物中酶的链长决定区域(CLDR)或其它区域的氨基酸序列,可实现FPP 合成酶与GGPP 合成酶的功能互相转化。Ohnuma 等[26]利用化学诱变法将热脂肪芽孢杆菌(Bacillusstearather mopbilus)的FPP 合成酶成功转化为GGPP 合成酶。Ohnuma等[27]通过定点突变的方式在古细菌中将GGPP 合成酶基因转变为了能表达FPP 合成酶的基因。Kawasaki等[28]的研究表明灰孢链霉菌(Streptomyces griseolosporeus)中突变的GGPP 合成酶和FPP 合成酶可分别产生FPP 和GGPP。细菌类胡萝卜素的合成一般是经由甲羟戊酸(Mevalonate,MVA)途径[29],结合本实验结果推测出细菌类胡萝卜素的潜在生物合成途径如图8 所示。

图8 细菌类胡萝卜素的潜在生物合成途径Fig.8 The potential biosynthetic pathway for carotenoids in bacteria
在MVA 途径中,2 分子乙酰辅酶A(Acetyl-Coenzyme A,Acetyl-CoA)经缩合形成3 羟基-3-甲基戊二酰辅酶A(3-Hydroxy-3-Methyl-Glutaryl-Coenzyme A,HMG-CoA),接着在HMG-CoA 还原酶(HMG Reductase,HMGR)的作用下生成甲羟戊酸。甲羟戊酸再经多个相关酶的催化下合成前体物质IPP(C5),及在IPP 异构酶idi(IPP Isomerase)的作用下生成其同分异构体DMAPP。在C30类胡萝卜素合成途径中,反应起始于IPP 与DMAPP 在ispA 作用下的缩合生成前体物质FPP(C15);2 分子的FPP 在脱氢角鲨烯合成酶crtM 的作用下生成C30类胡萝卜素diapophytoene,再经脱氢酶crtN 催化生成diaponeurosporene(C30)。在C40类胡萝卜素合成途径中,FPP 经crtE/ispA 催化合成前体物质GGPP(C20);2 分子的GGPP 在八氢番茄红素合成酶crtB 的作用下生成无色的八氢番茄红素phytoene(C40),再由八氢番茄红素脱氢酶crtI 和番茄红素环化酶crtY 催化去饱和环化反应最终生成β-胡萝卜素。
3 结论
本论文利用植物乳植杆菌LpispA基因功能替换质粒pAC-BETA 的crtE基因,构建了基于植物乳植杆菌LpispA基因的大肠杆菌功能互补表达系统pAC-BETAΔE-ispA,通过UV-Vis 和RP-HPLC 法测定含重组质粒pAC-BETAΔE-ispA 大肠杆菌工程菌株的类胡萝卜素生物合成,验证了LpispA 的功能。结果表明,构建的大肠杆菌工程菌株pAC-BETAΔE-ispA具有β-胡萝卜素合成功能,推测LpispA功能性替代了crtE的作用而合成C40β-胡萝卜素的前体物质GGPP,LpispA 为首次在植物乳植杆菌合成的类胡萝卜素途径中发现底物特异性变宽的酶,为类胡萝卜素合成多功能酶。本研究构建的LpispA基因功能互补的大肠杆菌表达系统,为植物乳植杆菌类胡萝卜素的生物合成机制研究提供了实验基础,有利于推动植物乳植杆菌作为类胡萝卜素补充剂的应用,扩充了利用异源系统工程菌株生产更多新型的高价值的化合物所需的基因库。
