牛支原体兔体攻毒模型的建立
2024-04-02武文英胡萌婕赵逸轩张宇豪郝成武郭爱珍陈建国陈颖钰
武文英,夏 青,胡萌婕,赵逸轩,王 琛,张宇豪,郝成武,贺 笋,郭爱珍,陈建国,2,陈颖钰*
(1.华中农业大学动物科学技术学院动物医学院,武汉 430070;2.华中农业大学生猪健康养殖协同创新中心,武汉 430070;3.华中农业大学农业微生物资源发掘与利用国家重点实验室,武汉 430070;4.天康生物制药有限公司,乌鲁木齐 830011)
牛支原体是牛呼吸道疾病综合征(bovine respiratory disease,BRD)的主要病原之一[1],能够引起牛的肺炎、中耳炎、关节炎及乳房炎等[2-3],发病率最高可达100%,死亡率可达50%[4],给养牛业造成了巨大的经济损失。自1961年首次被发现以来[5],牛支原体病已在世界范围内广泛传播;我国自2008年首次在湖北随州的牛肺组织中分离到该病原[6],之后在多地出现了该病的报道[7]。牛支原体可感染各个年龄段甚至各种品种的牛[8],其中肉牛和奶牛最为易感,犊牛病情相对严重,病死率较高[9]。牛支原体可在牛群中水平及垂直传播[10]。
目前,牛支原体尚无特效治疗药物。一方面由于缺乏细胞壁[11],青霉素类的抗生素对其敏感性较差。另一方面,抗生素的滥用也导致了新型耐药菌株的不断出现[12]。因此,疫苗免疫无疑是一个最佳的选择。我国目前尚无牛支原体的疫苗问世[13],疫苗评估是疫苗研制过程中的重要环节之一,宿主动物免疫攻毒保护试验是必不可缺的,因此制备疫苗可再现的攻毒模型是必要的。本体动物牛的成本较高,寻求合适的小动物替代攻毒模型是牛支原体疫苗中必不可缺的步骤之一[14]。
目前,虽然有部分学者在进行小动物模型的研究[15-17],但操作上存在有一定的差别。大多数仅用牛支原体对动物进行直接攻毒。而事实上牛支原体条件致病,在不同免疫条件下动物的临床发病会存在有差别。因此,为系统研究牛支原体小动物感染模型,本研究以日本大耳白兔为研究对象,不仅采用了牛支原体的单独感染,同时模拟了免疫抑制及刺激免疫状态,即用地塞米松降低兔免疫力;利用免疫刺激物巯基乙酸盐[18]和孔蓝蛋白(KLH)[19],刺激免疫,因此能够更好地模拟临床上的真实情况。为牛支原体疫苗的研发提供了重要的技术储备。
1 材料与方法
1.1 材料
1.1.1 主要试剂 血蓝蛋白(Keyhole Limpet Hemocyanin,KLH)、最低基本培养基(minimum essential medium,MEM)、丙酮酸钠购自美国Sigma公司;琼脂糖、PPLO肉汤粉、酵母粉购自美国BD公司;酚红购自上海沪试有限公司;青霉素钠购自红宝兽药有限责任公司;马血清购自hyclone公司。
1.1.2 菌种、蛋白及实验动物 牛支原体HB0801株由本实验室分离保存。抗体检测用P579蛋白为本实验室制备并保存。
健康雄性2月龄日本大耳白兔(约1.5 kg)购于华中农业大学实验动物中心。通过PCR方法检测确认无支气管败血波氏杆菌、巴氏杆菌、肺炎克雷伯菌以及牛支原体。于试验前进行7天的安静饲养,缓解应激。动物试验经由华中农业大学科学伦理委员会审批符合动物实验伦理审查要求(伦理编号:HZAURAB-2022-0003)。
1.1.3 PPLO培养基配制 液体培养基:PPLO10.5 g,酵母粉2.5 g,丙酮酸钠0.5 g,加单蒸水定容至440 mL,121 ℃灭菌15 min,待培养基冷却后,加入马血清50 mL,灭菌10×MEM 5 mL,无菌4万单位·mL-1青霉素钠溶液5 mL。
固体培养基:PPLO10.5 g,酵母粉2.5 g,丙酮酸钠0.5 g,琼脂粉7.5 g,加单蒸水定容至440 mL,121 ℃灭菌15 min,待培养基温度降至45 ℃左右,加入马血清50 mL,灭菌10×MEM 5 mL,无菌8万单位·mL-1青霉素钠溶液5 mL,然后倾倒平板。
1.2 方法
1.2.1 HB0801攻毒菌株的复苏及计数 将原代HB0801菌株冻干粉接种于液体PPLO培养基中培养36 h后再以 1∶100 的比例转接至PPLO液体培养基中。37 ℃静置培养48 h后以1∶1 000比例再次转接至PPLO液体培养基中。自24 h起每隔12 h进行倍比稀释至109,用颜色改变单位(CCU)及平板菌落计数法CFU进行计数[20]。
将培养好的牛支原体高速离心浓缩至1010后置于4 ℃保存,以备攻毒使用。
1.2.2 常见呼吸道病原菌的检测 利用PCR方法对多杀性巴氏杆菌[21]、肺炎克雷伯菌[22]以及支气管败血波氏杆菌[23]进行检测。
用牛支原体Uvrc(GenBank登录号:AF003 959.1)特异性片段对牛支原体进行普通PCR检测[24]。用设计的引物和探针进行荧光定量PCR检测[25],引物见表1,反应条件及体系见表2。

表1 PCR引物

表2 PCR反应的体系及条件
1.2.3 分组及攻毒 将11只日本大耳白兔分为4组,除对照组2只兔外,攻毒组均为3只。
组1为仅攻击HB0801组:气管注射1010CFU·mL-1HB0801株,1 mL·只-1,连续攻击3 d。
组2为注射地塞米松后攻击HB0801组:连续5 d肌肉注射地塞米松0.5 mg·kg-1后,第6天开始气管注射1010CFU·mL-1HB0801株1 mL·只-1,连续攻击3 d。
组3为攻击HB0801后注射免疫刺激物KLH和巯基乙酸盐培养基组:气管注射1010CFU·mL-1HB0801株,1 mL·只-1,连续攻击3 d后,再连续3 d肌肉注射KLH 0.1 mg·只-1,同时腹腔注射巯基乙酸盐培养基1 mL·只-1。
组4为空白对照组:气管注射PPLO液体培养基连续3 d。
1.2.4 观察并记录临床症状 每日观察各组日本大耳白兔精神状态、呼吸状况,口鼻分泌物等临床症状,记录体温。
1.2.5 鼻拭子排菌情况检测 隔天采集兔鼻拭子至无菌PPLO液体培养基中,充分混匀后用0.45 μm[26]滤器过滤,取滤液500 μL接种至等体积PPLO液体培养基中,于37 ℃ CO2培养箱培养2~3 d。将菌液离心后取上清煮沸作为模板PCR扩增Uvrc片段检测牛支原体。
1.2.6 血清中抗体的检测 采用本实验室前期所建立的ELISA抗体检测法对完成安静饲养后,进行试验操作前1天的血清及攻毒后第7、14、21天以及第31天的血清进行抗体检测[27]。
具体步骤如下:包被P579蛋白0.1 μg·孔-1,4 ℃过夜后充分洗涤,加入50 μL 待检血清/阴阳性对照以及50 μL酶标单抗,充分混匀后置37 ℃温育60 min。充分洗涤后加入底物进行显色,10 min后进行终止。OD450nm读数。按照以下计算公式,计算阻断率(PI值)。
PI值=(1-S/N)×100%,其中,S=样品OD450nm值;N=阴性对照血清OD450nm平均值。
判定:试验成立的条件阴性对照血清OD450nm平均值>0.65,且<2.0,阳性对照血清PI值均>60%,则判为试验成立。
结果判定:待检样品PI值≥40%时,判为阳性;待检样品PI值<40%时,判为阴性。
1.2.7 肺部病理变化的评分及病理切片的制作 于攻毒后第31天将所有日本大耳白兔进行剖杀。观察并记录大体病变情况,并依据实变型肺部评分标准(LLS)对肺组织进行评分[28]。
评分标准:在LLS方案中,根据肺叶的大小,每个肺叶被分成几个三角形(每个尖叶和中叶7个,每个膈叶19个,附属叶8个)。每叶受影响的三角形数乘以5,再除以每叶的三角形数(完全受这些病变影响的叶的得分为5)。LLS的最高分为30分(每叶5分)。兔肺包括左尖叶、左膈叶、右尖叶、右中叶、右膈叶和心叶。
在眼观变化明显的组织交界处无菌采集约为1 cm3的肺部组织4%多聚甲醛-PBS 缓冲液中进行固定后制作HE染色切片。
1.2.8 肺组织中牛支原体的检测 无菌条件下,取病变肺组织 0.5 g 进行充分匀浆,离心后取上清过滤,接种至PPLO液体培养基中,置于37 ℃含有5% CO2培养箱中培养 72 h后观察培养基颜色变化。
2 结 果
2.1 HB0801攻毒菌株的复苏及计数
复苏后的牛支原体于36 h时可达到109CCU·mL-1及3.67×109CFU·mL-1。符合攻毒浓度要求。
2.2 临床症状的观察
各攻毒组兔均表现为精神状态变差,食欲下降,体温无显著变化。对照组兔均表现正常。
攻毒组中,组2临床变化最为显著,一只明显流涕(图1A),另一只鼻腔出现肉眼可见白色分泌物(图1B)。

A.流涕;B.鼻腔有白色分泌物A. Runny nose; B. White nasal discharge
2.3 鼻拭子中排菌情况检测
隔天对鼻拭子中牛支原体的排菌情况进行检测。检测结果如表3所示。PCR结果显示,各攻毒组于攻毒后第1天均可以检测到牛支原体,组1表现为间歇性排菌,虽然第1天有2/3的兔检测为阳性,但第3~5天均为阴性,第7~11天也仅有1/3的兔检测为阳性。表明该模型不能有效诱导稳定的排菌。
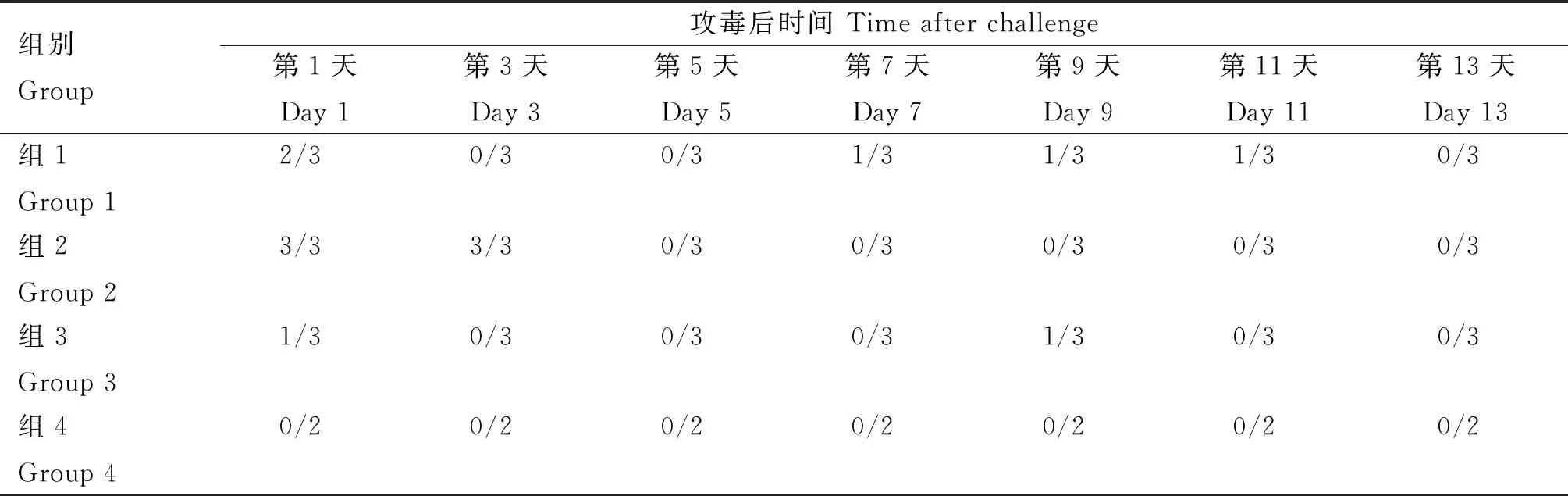
表3 PCR检测鼻拭子中牛支原体
组2虽然仅在攻毒后第1和3天检测到阳性,但所有的兔均表现为排菌,表明该模型稳定性较好。
组3仅第1天和第9天有1/3的兔检测到了牛支原体。
空白组均未检测到牛支原体。
2.4 血清中抗体的检测
对攻毒前及攻毒后第7、14、21、31天的血清进行抗体检测,结果显示(表4),组1自攻毒后第7天有1只兔开始检测到抗体,至第21天所有兔均为抗体阳性。
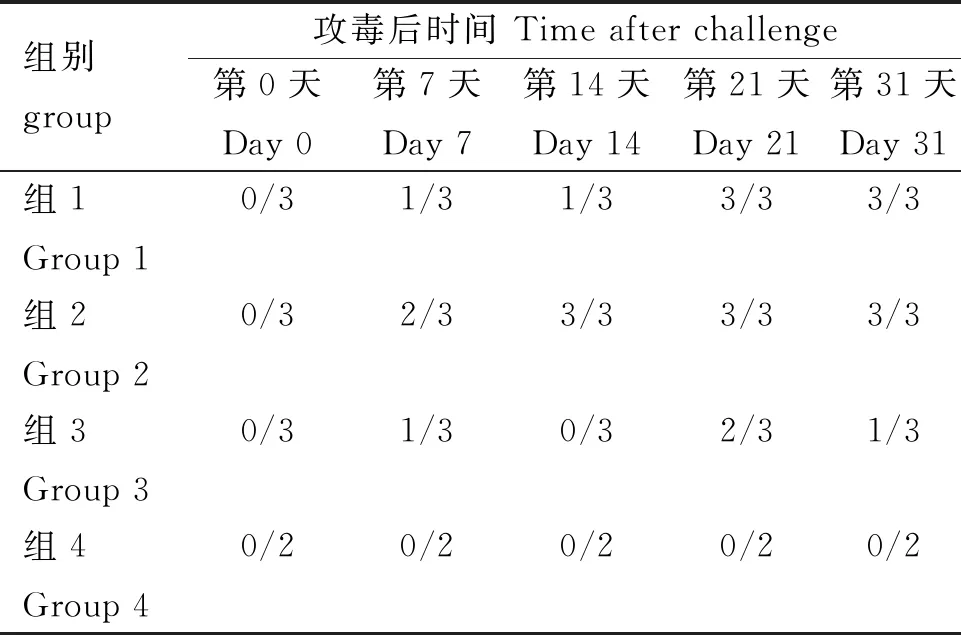
表4 血清ELISA检测结果
组2自攻毒后第7天开始,有2只兔检测为抗体阳性,随后所有兔转阳。表明该模型能够诱导较好的抗体水平。
组3抗体水平呈现波动,仅第7、21和31天能够检测到部分兔的抗体。
空白对照组均为抗体检测阴性。
2.5 肺部病理变化
于攻毒后第31天对所有兔进行剖杀。发现各攻毒组兔肺均出现不同程度的病变。组1中各组兔均有不同程度的淤血,组2除有明显出血外,有2只兔有明显肉变(图2)。组3除1只兔有肉样变化外,其它仅有淤血。组4肺部无显著病理变化。依据评分标准对各肺脏进行评分,结果显示(表5),在各攻毒组中,组2评分最高,为2.82分。
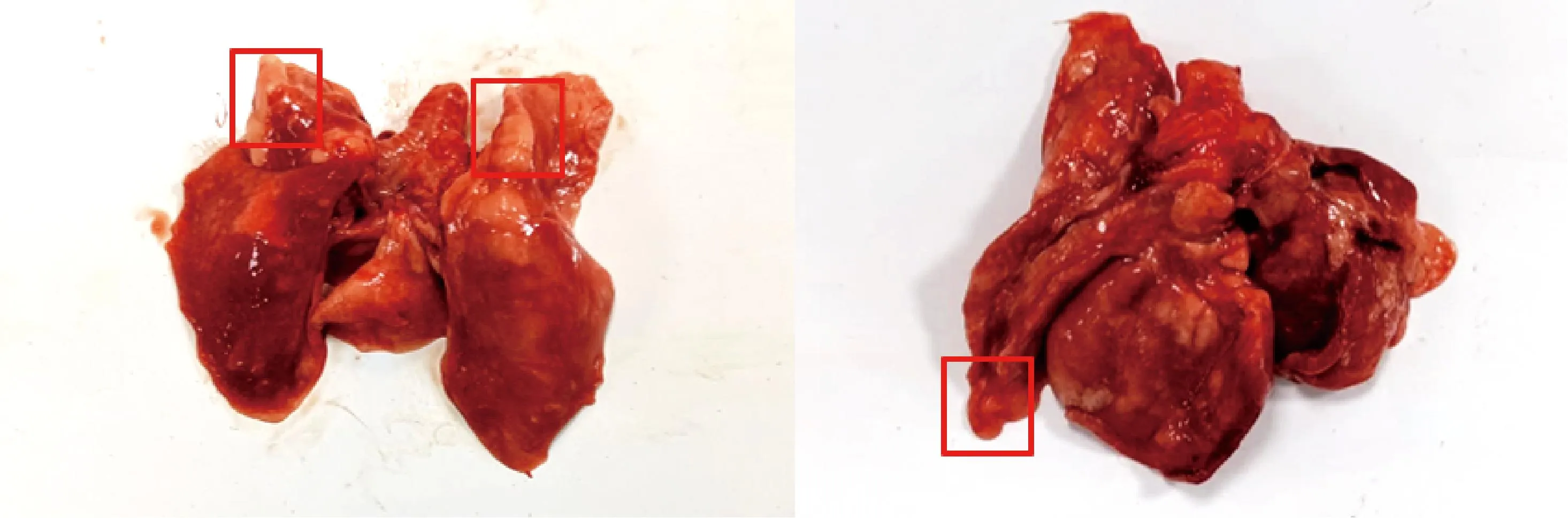
图2 组2兔肺部多处出现肉样病变Fig.2 Multiple fleshy lesions in the lungs of group 2 rabbits

表5 各组兔肺脏得分
进一步的病理切片结果(图3)显示,各攻毒组均表现为肺泡壁增厚,肺泡Ⅱ型上皮细胞增生,大量炎性细胞聚集,巨噬细胞浸润,有炎性渗出物,符合牛支原体造成的间质性肺炎的病变特征[29]。
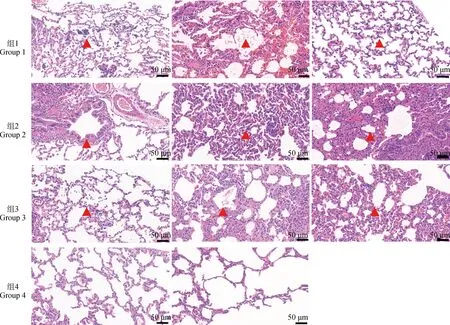
图3 各组兔肺病理切片Fig.3 Pathological sections of the lungs from rabbits of each group
该部分研究结果显示,用地塞米松诱导后再进行牛支原体的攻毒能够引起最显著的病理变化。
2.6 肺组织中牛支原体的检测
在肺组织中,各组兔均未检测到多杀性巴氏杆菌、肺炎克雷伯菌以及支气管败血波氏杆菌。仅组2的一只兔用PCR及荧光定量PCR检测均为阳性。其他组均未检测到阳性。详见图4。
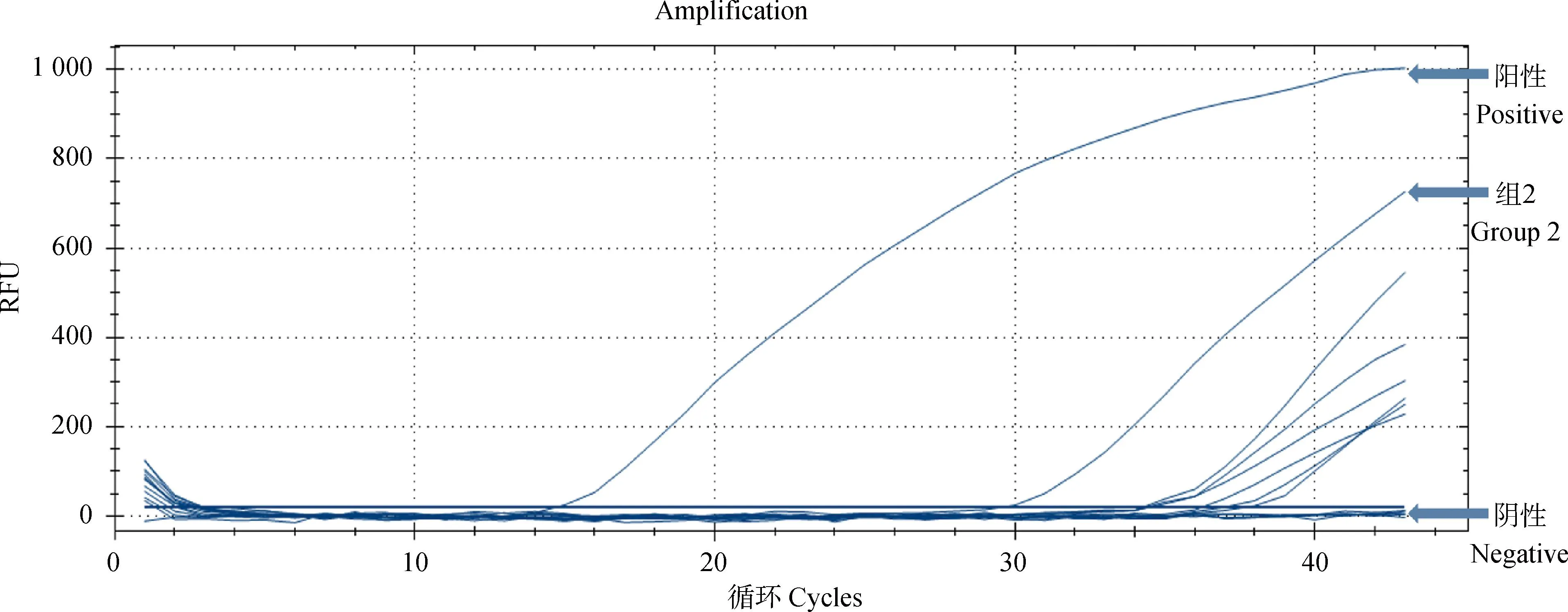
图4 荧光定量PCR结果Fig.4 The result of qPCR
3 讨 论
牛支原体对于宿主的特异性要求较高,动物模型复制成为制约牛支原体疫苗研究的瓶颈问题。前期的研究显示,用鸡胚作为替代动物,虽有较好的致死性,但是缺乏较好的评价指标,临床症状、抗体水平等不明显。小鼠作为动物模型既不能引起明显的临床症状,也无显著病理变化[30]。选用兔作为牛支原体的代替模型,相较于其他动物有较好的易感性和临床意义[16]。
本试验采取气管注射的方法攻毒兔,设置地塞米松免疫抑制组和KLH和巯基乙酸盐培养基作为免疫刺激物的组别,模拟在实际情况中,牛支原体作为条件致病菌,常在应激条件下由于免疫力下降时造成的继发感染[31]。利用药物降低动物的免疫力达到一定的攻毒效果,是之前试验均没有尝试过的,这样做是为了尽可能地贴合实际情况,诱发更为典型的病理变化。
综合临床症状、排菌、抗体及大体病变等各项指标,用地塞米松诱导后再进行HB0801攻毒确实表现出了稳定的攻毒效果,能够诱导出流涕、鼻腔出现白色分泌物等临床症状,能够导致尖叶和心叶的肉样变,使得肺泡壁增厚,Ⅱ型肺泡细胞增生,出现纤维素性渗出以及巨噬细胞浸润等病理现象;病原和免疫反应方面,能够出现稳定的排菌和较长的抗体持续时间。但与自然感染患牛相比,体温未出现明显升高,肺部的实质性病变也不够明显[32]。
4 结 论
综合本研究结果,得出以下结论:1)日本大耳白兔感染牛支原体模型建立成功。2)地塞米松造成免疫抑制后连续3 d,每天每只兔气管攻毒1 mL浓度为1010CFU·mL-1的牛支原体HB0801株的感染模型最好。
