催乳素通过酪氨酸激酶2/信号传导及转录激活蛋白5信号通路促进奶牛乳腺上皮细胞乳铁蛋白合成和分泌
2024-04-02杨跃杰王建发
杨跃杰 连 帅,2 武 瑞,2 王建发,2*
(1.黑龙江八一农垦大学动物科技学院,大庆 163319;2.黑龙江省牛病防制重点实验室,大庆 163319)
乳铁蛋白(LF)是一种天然的属于转铁蛋白家族的阳离子糖基化球状蛋白,存在于各种哺乳动物分泌物中,包括初乳、牛奶、唾液、眼泪和黏液,在先天免疫防御系统中起着至关重要的作用[1]。近年来,LF因在治疗病理方面和调节生理方面的潜在应用而受到诸多学者的关注。除了螯合游离的二价铁离子(Fe2+)和三价铁离子(Fe3+),LF的生物学功能也涉及抗菌、抗炎、抗病毒、参与铁代谢、抗氧化、抗肿瘤和提高自身抗体等多领域[2]。我国乳制品工业所需LF原料98%依靠进口,如何提高牛乳中LF的含量也是目前的难题,能否保障LF的自主可控供应直接决定了我国乳品产业的安全水平和核心竞争力[3]。
目前为止,LF的基因表达和合成分泌受到多维性和整体性的不同影响。根据体内和体外研究,LF的表达和分泌受核受体、转录因子、生长因子、刺激和炎症的调控[2]。已有研究表明,频繁刺激乳头挤奶可显著降低牛奶中LF的含量,延长挤奶间隔可增加LF的产量[4-5]。另外,4 mmol/L缬氨酸处理山羊乳腺上皮细胞增加了细胞内LF的分泌[6]。在试验中给小鼠喂食铁(120 mg/kg),结果表明铁显著增加哺乳期第1天和第25天乳腺组织LFmRNA的表达[7]。报道显示,视黄醇和雌激素呈时间和剂量依赖性诱导小鼠胚胎干细胞中LF的表达[8]。与此同时,在人类和小鼠LF启动子上发现的雌激素受体结合元件以受体和激素依赖的方式上调乳腺上皮细胞LFmRNA的表达水平,同时甲氧氯通过GC-Ⅱ序列刺激雌激素受体无效小鼠子宫中LF基因的表达[9-10]。因此,明确LF基因表达调控规律对通过体外生物技术合成LF具有重要意义。
LF启动子区域存在多个转录结合位点,特异性蛋白-1(SP-1)和雌激素受体结合在鼠LF启动子上,电泳迁移率转移试验中证明激活蛋白-2(AP-2)和SP-1在调控猪LF的转录中发挥重要作用[11-12]。同时,牛LF具有信号传导及转录激活蛋白5(STAT5)的转录结合位点,而催乳素(PRL)调控STAT5在奶牛乳腺上皮细胞(BMECs)产奶过程中起到核心作用[13-14]。此外,PRL通过酪氨酸激酶2/STAT5(JAK2/STAT5)信号通路的活化,可以促进哺乳期乳腺中乳脂、乳糖和乳蛋白的合成与分泌[15-16]。不同浓度的PRL和赖氨酸组合升高体外BMECs中LF的基因表达[17]。因此,本试验检测了PRL对BMECs中LF合成和分泌的影响,分析了PRL调控LF合成的作用机制,旨在为提升牛奶中功能性蛋白含量提供理论依据。
1 材料与方法
1.1 试验材料
PRL(HEK293)、匹莫齐特(Pimozide,HY-12987)购自MedChemExpress公司,酶联免疫吸附试验(ELISA)检测试剂盒(SEA780Bo)购自武汉云克隆科技有限公司,Cy3标记山羊抗小鼠(H+L)(A0521)、Cy3标记驴抗山羊(H+L)(A0502)和Alexa Fluor 488标记的山羊抗兔 (H+L)抗体(A0423)均购自上海碧云天生物技术股份有限公司,HRP-山羊抗兔二抗(H+L)(SA00001-2)、HRP-兔抗山羊二抗(H+L)(SA00001-4)和HRP-山羊抗鼠二抗(H+L)(SA00001-1)均购自Proteintech Group公司,HRP Anti-LF抗体(ab112970)和Anti-角蛋白-18抗体(ab668)均购自Abcam公司,JAK2(bs-23003R)和STAT5(bs-1142R)抗体均购自BIOSS公司,磷酸化抗体JAK2(AF3022)购自AFFINIY公司,磷酸化STAT5抗体(4322S)购自CST公司。
1.2 BMECs的分离纯化与鉴定
本试验采集了黑龙江省某正规奶牛屠宰场1头处于泌乳高峰期健康的中国荷斯坦奶牛乳腺组织。用手术刀切碎乳腺组织(0.5~1.0 mm3),用胶原酶Ⅰ型消化,37 ℃轻摇。用无菌的100目细胞筛过滤消化液和组织块后,收集细胞,1 200×g离心3 min。细胞在37 ℃的5% CO2细胞培养箱中用添加10%胎牛血清、1%青霉素(100 U/mL)/链霉素(100 mg/mL)的DMEM/F12培养基培养。采用差异胰蛋白酶分离出BMECs,验证BMECs的来源和纯度。免疫荧光法检测BMECs的标志蛋白(角蛋白-18)的表达。最后使用荧光显微镜拍摄。
1.3 BMECs合成分泌LF功能的鉴定
将BMECs以2×105个/mL的浓度接种于6孔板中,待细胞生长至80%时,使用饥饿培养基(无血清、无激素)培养纯化后的BMECs 16 h后,添加不同浓度的PRL[0(对照)、10、100、1 000 ng/mL]处理BMECs 24 h,为了验证STAT5在PRL促进BMECs LF合成和分泌的关键作用,添加STAT5抑制剂Pimozide [未添加Pimozide(对照)、10 μmol/L Pimozide、100 ng/mL PRL、10 μmol/L Pimozide+100 ng/mL PRL]处理BMECs 24 h(本研究添加的PRL浓度参考国内外通过放射免疫分析法检测的奶牛体内PRL浓度设置,同时探索高浓度PRL对BMECs中LF的影响[18-23];添加PRL培养BMECs 24 h参考前人[24]的研究方法)。收集细胞上清液,用ELISA试剂盒检测BMECs中LF的含量;收集细胞检测LF基因相对表达量和蛋白表达量;细胞使用LF荧光抗体染色,免疫荧光检测细胞内LF的荧光强度。所有试验均独立重复至少3次,每组技术重复3次。
1.4 LF、PRLR和JAK2/STAT5信号通路相关基因相对表达量的检测
利用总RNA提取试剂提取不同条件处理的BMECs中总RNA,根据PrimeScriptTMRT reagent Kit with gDNA Eraser将RNA反转录为cDNA。按照NovoStart®SYBR qPCR SuperMix Plus试剂盒说明书,检测BMECs中LF、PRLR、JAK2和STAT5基因相对表达量。反应程序为:95 ℃、1 min预变性;95 ℃、20 s变性,60 ℃、1 min延伸;40个循环。所用引物由上海生物工程有限公司合成,引物序列如表1所示。以β-肌动蛋白(β-actin)作为内参基因,目的基因相对表达量用2-△△Ct法计算。

表1 实时荧光定量PCR引物序列
1.5 BMECs中LF和p-STAT5相对荧光强度检测
将BMECs接种于待检测培养皿中,细胞培养至融合,洗涤,BMECs用4%多聚甲醛固定,0.5%Triton X-100溶液处理,3%牛血清白蛋白溶液(BSA)封闭,用p-STAT5和LF分别抗体孵育,在4 ℃过夜。荧光二抗37 ℃染色2 h。用抗荧光淬灭封片液(含DAPI)细胞核染色。最后在荧光显微镜观察荧光强度和入核水平。
1.6 BMECs中LF蛋白和JAK2/STAT5信号通路相关蛋白表达量检测
将处理后的BMECs在含有蛋白磷酸酶抑制剂混合物的放射免疫沉淀法缓冲液中裂解。使用BCA蛋白检测试剂盒对其浓度进行定量后,等量的样品总蛋白(20 μg)用十二烷基硫酸钠-聚丙烯酰胺凝胶(SDS-PAGE)制备试剂盒制备10%的SDS-PAGE电泳并转移到甲醇活化的聚偏二氟乙烯膜上。一抗为LF、JAK2、p-JAK2、STAT5、p-STAT5 1∶1 000、甘油醛-3-磷酸脱氢酶(GAPDH)1∶10 000,二抗根据一抗的种属来源不同选择。免疫反应条带采用超敏增强化学发光试剂盒进行可视化,使用Image Lab软件(Bio-Rad)测定条带强度。蛋白表达水平归一化为GAPDH表达,将蛋白磷酸化水平归一化为相应蛋白的总水平。
1.7 数据分析和统计分析
使用Graphpad prism 9.0软件进行统计学分析,结果表示为平均值±标准差。采用one-way ANOVA进行单因素方差分析,P<0.05为差异显著,P<0.01为差异极显著。
2 结果与分析
2.1 BMECs体系分离、纯化、标志蛋白鉴定
由图1-A可见组织块经过72 h后开始生长大量细胞;由图1-B可见原代纯化培养后,BMECs多呈现卵圆形;由图1-C可知BMECs在25 m3细胞培养瓶中细胞计数为1.63×106个/mL,存活率达到97%。由图1-D、图1-E和图1-F可知经免疫荧光检测,BMECs中CK-18在细胞上明显表达,证明本试验已获得纯化的BMECs,可用于后续试验。

A:组织块接种72 h后生长出的上皮样细胞;B:纯化后BMECs细胞;C:25 m3细胞培养瓶中BMECs细胞计数;D: BMECs细胞核DAPI染色;E:BMECs细胞CK-18染色;F:D、E合图。
2.2 LF的ELISA、实时荧光定量PCR的检测结果
为了检测PRL对BMECs中LF含量的影响,用不同浓度的PRL处理BMECs 24 h后检测细胞培养上清液中LF的含量。由图2-A可知,BMECs细胞上清液中的LF含量随着PRL浓度的增加,呈现先增加后下降的趋势,与对照组相比,10、100、1 000 ng/mL PRL能够显著或极显著提高BMECs中LF的含量(P<0.05或P<0.01)。由图2-B可知,与对照组相比,10 ng/mL PRL处理使BMECs中LF基因相对表达量显著上调(P<0.05),100 ng/mL PRL处理使BMECs中LF基因相对表达量极显著上调(P<0.01),1 000 ng/mL PRL处理使BMECs中LF基因相对表达量高于对照组,但无显著差异(P>0.05)。
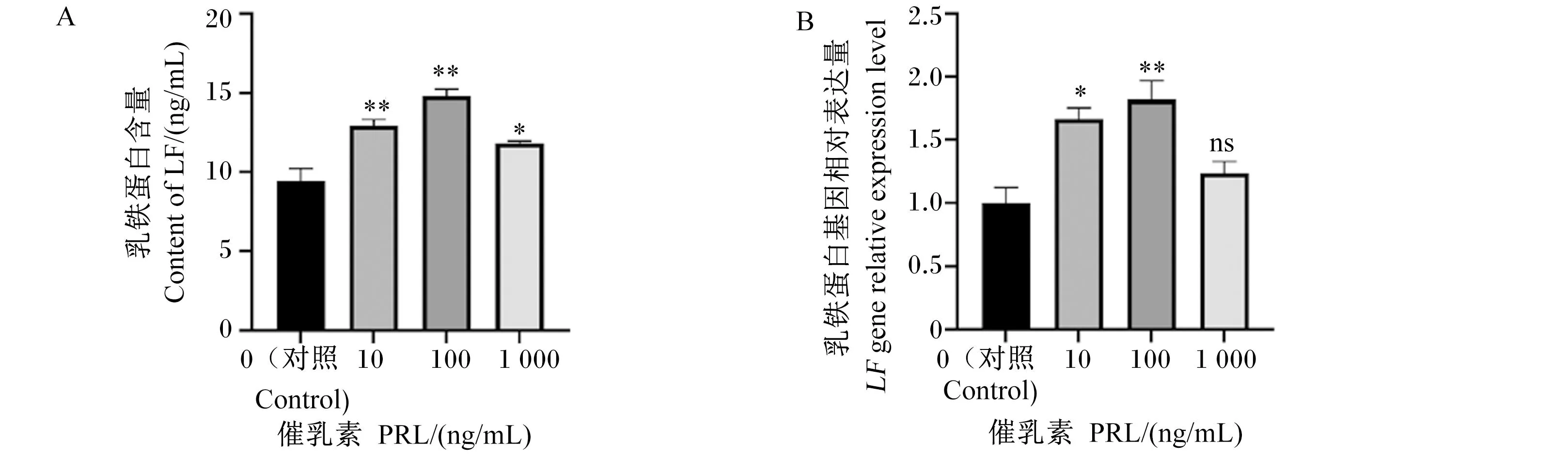
与对照组相比,*表示差异显著(P<0.05),**表示差异极显著(P<0.01),ns表示无统计学差异(P>0.05)。下图同。
2.3 PRL处理细胞可促进JAK2/STAT5相关基因转录
由图3可知,与对照组相比,10 ng/mL PRL处理使BMECs中JAK2、STAT5和PRLR基因相对表达显著上调(P<0.05),100 ng/mL PRL处理使BMECs中JAK2和PRLR基因相对表达量极显著提高(P<0.01),100 ng/mL PRL处理BMECs中STAT5基因相对表达显著上调 (P<0.05),1 000 ng/mL PRL处理使BMECs中PRLR基因相对表达量显著上调(P<0.05),1 000 ng/mL PRL处理使BMECs中STAT5基因相对表达量极显著上调(P<0.01),1 000 ng/mL PRL处理使BMECs中JAK2基因相对表达量提高,但无显著差异(P>0.05)。
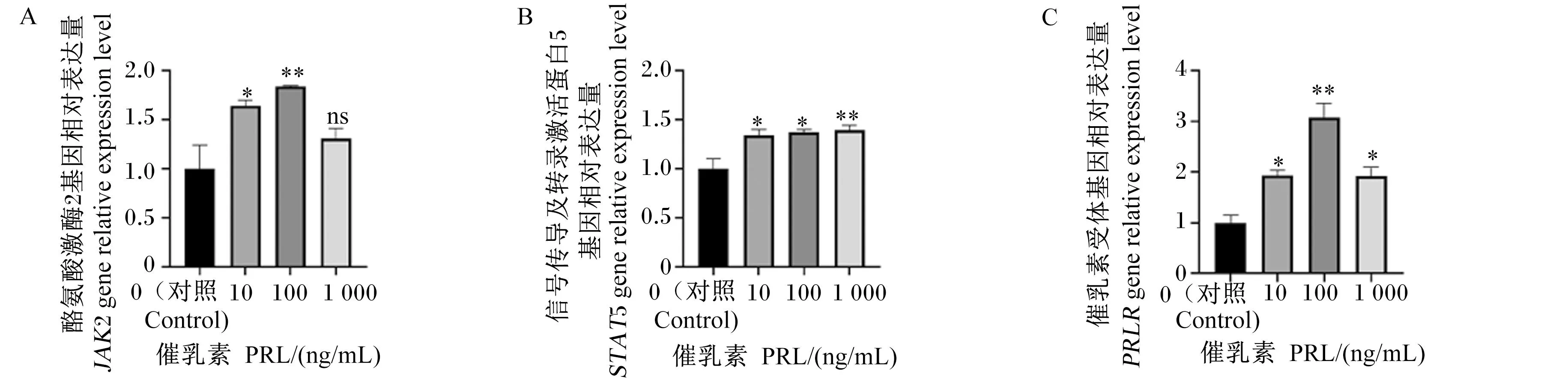
图3 不同浓度PRL处理BMECs中JAK2、STAT5和PRLR基因表达量的结果
2.4 PRL处理细胞可促进JAK2/STAT5信号通路蛋白表达
由图4可知,随着PRL浓度的升高,p-JAK2的蛋白表达量呈逐渐上升的趋势,与对照组相比,10、100、1 000 ng/mL的PRL处理的BMECs中p-JAK2的蛋白表达量极显著升高(P<0.01);p-STAT5的蛋白表达量随着PRL浓度增加呈先升高后下降的趋势,10和1 000 ng/mL的PRL处理的BMECs中p-STAT5的蛋白表达量与对照组相比显著升高(P<0.05),100 ng/mL的PRL处理的BMECs中p-STAT5的蛋白表达量与对照组相比极显著升高(P<0.01);随着PRL浓度的升高,LF的蛋白表达量呈逐渐上升的趋势,100和1 000 ng/mL的PRL处理的BMECs中LF的蛋白表达量与对照组相比极显著升高(P<0.01),10 ng/mL的PRL处理的BMECs中LF的蛋白表达量与对照组相比升高,但无显著差异(P>0.05)。

图4 不同浓度催乳素处理BMECs中LF、JAK2和STAT5蛋白表达的结果
2.5 PRL和Pimozide对BMECs中LF和p-STAT5相对荧光强度的影响
免疫荧光检测结果(图5)显示,与对照组相比,Pimozide组BMECs中p-STAT5的相对荧光强度极显著降低(P<0.01),PRL组BMECs中p-STAT5的相对荧光强度极显著升高(P<0.01),Pimozide+PRL组p-STAT5的相对荧光强度与对照组相比显著升高(P<0.05)。
由图6可知,Pimozide组LF的相对荧光强度与对照组相比显著降低(P<0.05),PRL组LF的相对荧光强度与对照组相比极显著升高(P<0.01),Pimozide+PRL组LF的相对荧光强度与对照组相比显著升高(P<0.05)。
2.6 Pimozide对BMECs中LF含量的影响
由图7-A可知,在用Pimozide处理上皮细胞24 h后,与对照组相比,Pimozide组细胞上清液中LF含量极显著下调(P<0.01),PRL组细胞上清液中LF含量极显著上调(P<0.01)。由图7-B可知,与对照组相比,Pimozide组LF基因相对表达量显著下调(P<0.05),PRL组LF基因相对表达量显著上调(P<0.05)。由图8-A和图8-B可知,Pimozide组LF的蛋白表达量与对照组相比极显著降低(P<0.01),PRL组LF的蛋白表达量与对照组相比极显著升高(P<0.01),Pimozide+PRL组LF的蛋白表达量与对照组相比极显著降低(P<0.01)。结果表明,STAT5抑制细胞模型构建成功,且抑制JAK2/STAT5信号通路部分抑制BMECs中LF的合成和分泌。

图7 PRL和匹莫齐特对BMECs中LF含量及基因表达的影响
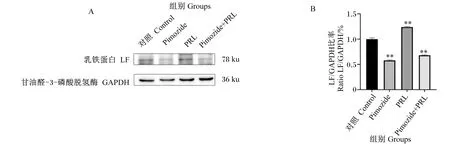
图8 PRL和匹莫齐特对BMECs中LF蛋白表达的影响
3 讨 论
BMECs合成和分泌乳蛋白是复杂的过程,受到多种因素的影响。早期通过隔绝细胞与基质的接触途径的方式,通过引起细胞形态变化而直接影响着细胞的生长和分化,这种调节细胞形状的方式也被证明可以调节其细胞功能[25-26]。乳腺上皮细胞的形态具有其他贴壁细胞类型相似的特性,都是极其容易受到周围细胞密度和细胞外基质变化的影响。与牛奶中的β-酪蛋白和乳清蛋白不同,LF在乳房发育过程中由乳腺上皮细胞大量合成和分泌,乳腺上皮细胞形状的改变与LF的表达息息相关[27]。此外,乳腺上皮细胞中的细胞形状和肌动蛋白细胞骨架的改变可以介导LF的表达[28]。本试验通过组织块贴壁法提取原代BMECs,经过纯化培养后进行LF的检测。通过与前人ELISA检测BMECs中LF分泌量相比,本试验纯化的细胞分泌LF的能力与其他人差异不大[29]。因此,本试验获得纯化细胞形态良好的BMECs用于后续检测调控LF合成和分泌的机制研究。
乳腺上皮细胞的生长分化和蛋白质的合成分泌受到细胞基质、细胞形态、多肽类和类固醇类激素等多种因素的调控[30]。JAK2/STAT5信号通路参与乳腺上皮细胞的增殖、分化和乳蛋白的合成分泌。有研究报道,STAT5促进乳腺上皮细胞的增殖,JAK2/STAT5途径的抑制会破坏乳腺形态发生和分化,抑制STAT5活化会减弱小鼠乳腺上皮细胞的增殖[31-32]。同时,STAT5除了对乳腺发育很重要,也被认为是参与调节和控制哺乳动物乳蛋白合成的主要因子[33]。前人的结果表明,炎症反应、细胞因子、氨基酸和激素等都可以通过STAT5的活化调控牛奶蛋白的合成和分泌[34-36]。PRL是一种由垂体、乳腺、肝脏和性腺等组织分泌的多肽类激素,参与多种生理功能,包括葡萄糖和脂质代谢作用、泌乳和性腺功能以及免疫调节作用等[37-38]。Lollivier等[39]试验结果显示,喹那啉内酯可以减少奶牛的产奶量并抑制奶牛乳腺上皮细胞中α-乳清蛋白和κ-酪蛋白的基因表达。不同浓度的PRL和赖氨酸组合升高体外BMECs中LF、β-酪蛋白和α-乳白蛋白的基因表达[17]。目前已知PRL、STAT5和乳蛋白之间存在交互网络,因此我们研究了PRL调控LF合成和分泌的机制,探究JAK2/STAT5信号通路与LF之间的作用关系。本试验结果表明,不同浓度的PRL处理BMECs后可以提高LF的含量,且10、100、1 000 ng/mL PRL可以促进LF、PRLR、JAK2和STAT5的基因表达,p-JAK2、p-STAT5和LF的蛋白表达呈现上升趋势。
同时,LF基因表达调控位点除了STAT5,还具有类固醇激素、感染应答转录因子和细胞分化转录因子等多个重复元件和结合位点[40]。其中牛LF基因启动子区域的类固醇激素结合位点包括雌激素受体结合位点和糖皮质激素受体结合位点。在鼠LF的启动子上发现的雌激素受体结合元件能够以受体和激素依赖的方式上调鼠乳铁蛋白的启动子转录过程[41]。此外,乳腺上皮细胞中LF的表达和产生受到LPS的刺激,牛乳铁蛋白的转录活性可被LPS通过牛乳铁蛋白启动子区域上的核因子-κB(NF-κB)结合位点诱导,且具有剂量依赖性[42]。因此,为了进一步对LF基因表达调控位点进行分析。本试验添加了Pimozide处理BMECs。通过ELISA、qRT-PCR、免疫荧光技术和Western blot检测结果显示,添加Pimozide抑制了LF的合成和分泌。
综上所述,PRL对于LF的转录和分泌过程发挥了重要的调节作用。结果表明,PRL通过JAK2/STAT5途径,促进LF的合成和分泌,从而导致奶牛产奶量的升高。通过对PRL调控LF机制的深入研究,有助于激素与LF之间调控网络的构建,为调控牛奶中LF的含量提供理论依据,丰富了体外调节LF基因表达和分泌的理论知识。通过明确了PRL促进LF合成和分泌的机制,在生产实践中可以使用多巴胺受体抑制剂提高奶牛PRL分泌量进而提高奶产量和LF产量,也可以使用合成生物技术在构建的基盘细胞或元件中过表达STAT5结合位点序列,在PRL环境下大量分泌LF。因此,通过调节体内激素水平或体外构建过表达STAT5的方式,发挥激素调控LF合成和分泌的作用,将对我国牛奶功能性蛋白含量的提高和我国奶业持续健康的发展具有深远意义。
4 结 论
添加10、100、1 000 ng/mL PRL处理BMECs可以促进LF的合成和分泌,其中100 ng/mL PRL处理结果最佳,并且STAT5抑制后,试验结果表明细胞中LF的表达下降。因此,PRL通过JAK2/STAT5信号通路促进BMECs中LF合成和分泌。
