植物纤维的降解机制及菌酶协同发酵应用
2024-04-02易兰兰朱俊红程文杰赵素梅
易兰兰 朱俊红 程文杰 赵素梅
(云南农业大学动物科学技术学院,昆明 650201)
微生物是饲料原料发酵的主要驱动力,但单纯的微生物发酵对饲料原料中纤维物质或抗营养因子的降解有限,在此基础上添加纤维素酶、植酸酶或蛋白酶等能有效降解纤维物质和抗营养因子,提高饲料原料营养价值[1]。外源性酶与微生物间的协同作用主要涉及纤维素酶和木聚糖酶,它们分别降解纤维素和半纤维素。纤维素酶由至少三大类酶组成,即内切葡聚糖酶、外切葡聚糖酶和β-葡萄糖苷酶,同时木聚糖酶在半纤维素的降解中发挥作用,与纤维素酶协同降解纤维素[2]。微生物在发酵过程中能产生少量的酶,可构建发酵环境,提升发酵物品质,且部分益生菌进入畜禽消化道后仍发挥益生作用。Du等[3]报道,菌酶协同发酵桑叶改善桑叶青贮饲料的微生物群落结构、发酵品质,有效抑制有害微生物的生长,主导厌氧发酵过程,其作用机理是乳酸菌作为桑叶青贮过程的优势菌种,能通过糖酵解途径将葡聚糖酶、果胶酶和羧甲基纤维素酶分解纤维素和半纤维素产生的糖转化为乳酸。全株玉米接种枯草芽孢杆菌青贮,可降低粗纤维含量,增加粗蛋白质含量,提升全株玉米的营养价值[4],且枯草芽孢杆菌进入反刍动物消化道仍发挥益生作用。因此,菌酶协同发酵是实现饲料原料高值化利用的重要途径。
1 植物纤维的降解机制
植物纤维主要由纤维素、半纤维素和木质素组成。纤维素仅含通过氢键网络连接的晶体结构的葡萄糖链,半纤维素具有木糖链主链和各种乙酰化糖单元的侧链,木质素是由苯丙烷单体组成的复杂聚合物[5]。纤维素的生物降解涉及纤维素酶(内切葡聚糖酶)、纤维二糖水解酶(外切葡聚糖酶)和β-葡萄糖苷酶的协同作用[6]。内切葡聚糖酶切割纤维素链的无定形区域,使纤维二糖水解酶在链末端发挥作用。纤维二糖水解酶、内切葡聚糖酶和纤维素酶水解纤维素有“两步”过程,首先在几分钟内纤维长度减少,然后在60~90 min内无定形纤维素溶解(占初始纤维素的约2%),此外,膨胀素也能通过无定形纤维素来促进纤维断裂[7]。纤维素酶优先水解低序纤维素,纤维长度减少、发生膨胀并产生断裂[8]。纤维素酶作用于纤维素底物,是天然结晶纤维素完全酶促水解的关键。最后,β-葡萄糖苷酶将纤维二糖水解为葡萄糖。
结晶纤维素和其他顽固多糖(如几丁质)可被细菌和真菌中发现的溶解多糖单加氧酶降解[9]。里氏木霉生产的纤维素酶7B对纤维素的初始水解和溶胀的联合作用暴露了单个微纤维和微纤维束,导致纤维结构松散并在纤维表面暴露出微纤维[10]。半纤维素水解是内切酶(在内部裂解主链)、外切酶(释放单糖)以及相关酶(切割聚合物或寡糖侧链以最终释放单糖或二糖)协同作用的结果。此外,玉米中葡萄糖醛阿拉伯木聚糖的不溶性和高苯酚含量使木聚糖酶降解效率降低,体内和体外研究均表明脱支酶(阿拉伯呋喃糖苷酶、α-葡萄糖醛酸酶和酯酶等)能通过破坏酚键显著增加纤维的降解率[11]。微生物分泌的纤维降解酶将纤维素降解为畜禽可利用的单糖或寡糖。
2 菌酶协同发酵中的菌及其生产的酶
2.1 真菌生产的纤维降解酶
微生物在木质素的降解过程中产生多种水解酶,包括木质素过氧化物酶、纤维素酶、木聚糖酶、漆酶等[12]。木质素是一种芳香族聚合物,由愈创木基、紫丁香基和对羟基苯基3种亚基组成。黄孢原毛平革菌是白腐真菌的一种,其产生的木质素过氧化物酶于1982年被发现[13]。使用黄孢原毛平革菌、变色栓菌和平菇降解玉米秸秆能提高纤维素、半纤维素和木质素的降解率[14]。白腐真菌具有强大的木质素降解酶系统,能通过分泌漆酶、锰过氧化物酶、木质素过氧化物酶等木质素分解酶参与降解木质素[15]。漆酶属于多铜氧化酶家族,包括抗坏血酸氧化酶、氧化铁酶、亚硝酸还原酶和铜蓝蛋白等,其是由许多具有不同底物特异性和不同生物功能的蛋白质组成的酶,铜氧还蛋白样结构域使所有多铜氧化酶能将氧气还原为水,而不产生有害的副产物[16]。除白腐真菌能降解木质素外,部分细菌如红球菌、恶臭假单胞菌、芽孢杆菌、新鞘氨醇菌和弗氏柠檬酸杆菌也表现出木质素降解能力[17]。
真菌已被广泛用于生产纤维降解酶[18],具体见表1。里氏木霉常用于工业化生产纤维素酶和半纤维素酶[19]。有研究表明,里氏木霉细胞内糖转运蛋白利用乳糖协助其生产纤维素酶[20]。斜卧青霉以纤维素、脱淀粉麦麸和稻草为碳源能生产阿魏酸酯酶和纤维素酶[21]。在自然界广泛存在的酵母菌是子囊菌门真菌,具有降解纤维素和木聚糖的酶活性[22]。褐腐真菌能生物降解木材,降低其半纤维素和纤维素的含量和纤维素的结晶度[23]。

表1 真菌生产的纤维降解酶
真菌处理玉米秸秆常用于工业生产半纤维素酶,黑曲霉、巨大芽孢杆菌和酵母菌是主要用于生产木聚糖酶的菌种,木聚糖酶是分解半纤维素的主要酶类。黑曲霉和巨大芽孢杆菌分别接种于碱处理玉米秸秆,黑曲霉和巨大芽孢杆菌产生的木聚糖酶在40 ℃培养30和45 min后分别达到最高的活性(3.67和3.36 U/mL);pH 5.0时黑曲霉产生的木聚糖酶最高活性达4.58 U/mL,pH 6.2时巨大芽孢杆菌产生的木聚糖酶最高活性达3.58 U/mL[24]。巴斯德毕赤酵母产生的木聚糖酶在最适条件下活性达442.53 U/mL[25]。酿酒酵母MK-157在Sabouraud葡萄糖肉汤中30 ℃下培养48 h,将酵母细胞固定在碱处理玉米芯基质上,将固定化基质转移到含有1%木聚糖的矿物盐培养基中,并在30 ℃下孵育24 h,可生产木聚糖酶[26]。酿酒酵母SCPW17能发酵玉米芯生产纤维素酶、木聚糖酶和木质素酶,活性分别为17.63 U/gds、29.35 U/gds和150.75 μmol/min[27]。此外,使用黑曲霉F12发酵咖啡渣和麦麸的混合物组,120 h后β-甘露聚糖酶和β-糖苷酶的产量分别为51.97和72.54 U/g[28]。
2.2 细菌生产的纤维降解酶
将地球微生物组计划发布的序列与测序的原核基因组之间进行大规模比对,Li等[29]认为几乎所有原核生物群落都具有降解纤维素、木聚糖和几丁质的功能潜力。部分细菌利用不同底物生产的纤维素降解酶的种类见表2。原小单孢菌属VP111处理麦秆和甘蔗渣产生纤维素酶、木聚糖酶和果胶酶,其中纤维素酶水解多种纤维素底物,包括羧甲基纤维素钠、Whatman 1号滤纸、微晶纤维素、对硝基苯基-β-D-吡喃葡萄糖苷、昆布多糖和纤维素粉[30]。枯草芽孢杆菌亚种JJBS250以甘蔗渣为底物固态发酵生产木聚糖酶,产量为20.35 U/g底物[31]。嗜热地芽孢杆菌GS90在pH为8和温度为55 ℃条件下生产的木聚糖酶比活性最佳为1 671.4 U/mg[32]。高地芽孢杆菌RS3025利用碱处理的稻草废弃物生产木聚糖酶,72 h获得最大木聚糖酶活性(2 518.51 U/mL)[33]。芽孢杆菌属以麦麸为唯一发酵底物,144 h生产木聚糖酶2 943 U/g[34]。沙福芽孢杆菌M35和高地芽孢杆菌J208发酵柑橘皮和麦麸的混合底物,可同时生产木聚糖酶和果胶酶[35]。

表2 细菌生产的纤维降解酶
枯草芽孢杆菌能降解沉香粉的纤维素,纤维素酶活性高达0.22 U/mL,并且对花椒和黄檀具有类似作用[36]。在玉米秸秆中添加枯草芽孢杆菌、地衣芽孢杆菌、蜡样芽孢杆菌和链霉菌形成共降解系统,可促进子囊菌门产生更多腐殖质前体,提高纤维素分解效率[37]。假单胞菌属、黄杆菌属、固氮螺菌属、毛球菌属、不动杆菌属和短波单胞菌属降解秸秆14 d后,滤纸酶和内切葡聚糖酶的活性分别为2.55和2.34 U/mL;28 d后,木聚糖酶、漆酶和木质素过氧化物酶活性分别为9.86、132.16和85.43 U/mL;高通量测序结果发现变形菌门、厚壁菌门和拟杆菌门是秸秆不同降解阶段的主要菌门[38]。地芽孢杆菌属除具有高纤维素酶活性外,还具有良好的木聚糖酶、纤维二糖酶和淀粉酶活性,适用于生产各种生物技术和工业应用的酶[39]。芽孢杆菌PM06能在淀粉和羧甲基纤维素作为唯一碳源的培养基中生长,α-淀粉酶和纤维素酶在48 h时获得最大产量[40]。
甘露聚糖是半纤维素的主要成分,广泛存在于植物组织中,β-甘露聚糖酶是主要的甘露聚糖降解酶,可由不同的植物、动物、放线菌、真菌和细菌产生[41]。地衣芽孢杆菌HDYM-04以葡萄糖或魔芋粉为碳源时,24 h β-甘露聚糖酶活性分别为288.13和696.35 U/mL[42]。枯草芽孢杆菌US191在0.25 g/L刺槐豆胶、5 g/L咖啡渣粉、5 g/L枣子粉和5 g/L仙人掌果粉的条件下发酵生产β-甘露聚糖酶,产量为8.75 U/mL[43]。Thermobacillusxylanilyticus是一种嗜热半纤维素分解细菌,可用于生产热稳定性半纤维素酶,可使用麦麸和麦秆作为碳源生产半纤维素酶[44]。
木质素的降解率影响纤维素和半纤维素的降解率。木质素在堆肥处理中难以降解,进而影响纤维素和半纤维素的降解。堆肥处理的半纤维素、纤维素和木质素的降解率分别为19.9%、14.7%和12.5%,添加混合微生物处理后降解率分别提高至44.4%、34.9%和39.2%[45]。漆酶、木质素过氧化物酶和锰过氧化物酶处理玉米秸秆的试验发现漆酶是混合酶反应的起始酶,3种酶协同破坏木质素的关键结构并打开苯环,产生更多酸性化合物,木质素降解率达25.79%[46]。枯草芽孢杆菌和地衣芽孢杆菌都能分泌漆酶,漆酶降解木质素的能力已被充分证明[47]。解淀粉芽孢杆菌发酵降解秸秆15 d后木质素降解率为28.55%,与真菌达到的降解率接近[48]。因此,提高木质素的降解率是提高可溶性糖含量的有效途径。
2.3 其他酶
饲料中常使用植酸酶、脂肪酶、蛋白酶等提升其营养价值。植酸酶是饲料中的常用酶,能改善单胃动物生长性能。枯草芽孢杆菌JJBS250在30 ℃、pH 5.0的条件下发酵麦麸48 h后,分泌出高植酸酶滴度[49]。嗜热真菌发酵米糠,添加0.5%硫酸铵为氮源,在45 ℃下培养9 d,植酸酶、内切葡聚糖酶和木聚糖酶产量分别为58、12和48 U/g底物[50]。微生物脂肪酶催化长链脂肪酸的水解,芽孢杆菌属、无色杆菌属、产碱菌属、节杆菌属、假单胞菌属和属于真菌的青霉属、镰刀菌属、曲霉属能用于生产脂肪酶[51]。乳酸菌能将糖发酵为乳酸并产生蛋白水解酶,蛋白水解酶在细菌细胞内合成,大部分分泌到细胞外,其主要功能是将蛋白质水解为细菌能吸收的成分[52]。果胶酶是水解果胶的混合酶,主要存在于微生物和高等植物中,已能利用微生物尤其是真菌进行商业化制备[53]。淀粉酶可将淀粉水解成糊精和更小的葡萄糖单元聚合物,热稳定淀粉酶占淀粉酶市场的近25%[54]。
3 菌酶协同发酵的饲料原料
3.1 高纤维饲料原料
菌酶协同发酵饲料主要分为2种类型:一是单独发酵某种饲料原料后配制饲粮;二是配制全价饲料后添加菌酶进行发酵。根据饲料原料的营养特性使用不同的菌酶组合处理,其目的主要是降解纤维素和消除抗营养因子,菌酶协同发酵高纤维饲料原料的效果见表3。高纤维饲料原料以玉米秸秆为代表,全球玉米秸秆年产量约10亿t,主要被焚烧、掩埋和堆肥,难以形成经济效益。玉米秸秆主要可分为纤维皮和纤维芯两部分,纤维皮由纤维素和木质素组成,纤维芯主要包含半纤维素和少量木质素[55-56]。利用黄孢原毛平革菌、纤维素酶和木聚糖酶青贮玉米秸秆19 d后,其中性洗涤纤维(NDF)、酸性洗涤纤维(ADF)和酸性洗涤木质素(ADL)含量比未添加菌酶青贮玉米秸秆分别降低23.4%、20.13%和7.68%[57]。甜高粱与植物乳杆菌、纤维素酶和木聚糖酶青贮21 d,pH降至4.2以下,乳酸/乙酸>2.0,乳酸/总有机酸>0.65,实现甜高粱青贮效果优化[58]。在全株玉米和花生蔓中添加植物乳杆菌和纤维素酶,可使pH、ADF和NDF含量降低,乳酸含量增加,乳酸杆菌、克雷伯氏菌、沙雷氏菌和魏斯氏菌为主要微生物[59]。使用黑曲霉、嗜酸乳杆菌、布氏乳杆菌、产朊假丝酵母、纤维素酶和木聚糖酶室温密封发酵香菇菌糠30 d能改善其营养价值,使NDF、ADF、ADL、钙和磷含量下降,粗蛋白质和粗脂肪含量提高[60]。使用酿酒酵母、黑曲霉、枯草芽孢杆菌和植酸酶在30 ℃恒温培养箱中发酵辣木茎秆粉7 d,可提高其乳酸和蛋白质含量,降低NDF、ADF和植酸含量[61]。在苜蓿和羊草中添加植物乳杆菌和纤维素酶混合室温青贮1~30 d,pH随青贮时间的增加而降低,30 d时NDF含量、微生物Shannon指数和Simpson指数降低,乳酸菌的相对丰度增加,乳酸和乙酸的浓度提高[62]。桑叶中添加干酪乳杆菌和纤维素酶混合青贮,28 ℃发酵30 d,乳酸、真蛋白质和总黄酮含量提高,抗氧化活性和微生物蛋白产量增加,pH、氨态氮和纤维含量较低[63]。

表3 菌酶协同发酵高纤维饲料原料的效果
3.2 高蛋白质饲料原料
我国畜禽养殖中主要使用玉米-豆粕型饲粮,但豆粕含有抗营养因子如大豆球蛋白、β-伴大豆球蛋白、胰蛋白抑制剂和植酸,损害动物肠道健康,降低生产性能[64]。菌酶协同发酵可消除蛋白质饲料原料中的抗营养因子,其对高蛋白质饲料原料的效果见表4。使用枯草芽孢杆菌和碱性蛋白酶30 ℃发酵豆粕72 h,β-伴大豆球蛋白消除率为75.27%,氨基酸含量提高3.96倍,并能提高乳酸、柠檬酸、乙酸等有机酸的含量,显著改善豆粕的营养价值和适口性[65]。使用酿酒酵母、枯草芽孢杆菌、干酪乳杆菌和蛋白酶发酵豆粕,37 ℃发酵72 h,水苏糖、棉籽糖、大豆球蛋白、β-伴大豆球蛋白、胰蛋白酶抑制因子等抗营养因子消除率增加[66]。花生粕存在抗营养因子、过敏蛋白和黄曲霉毒素等[67]。使用短乳杆菌、纤维素酶、植酸酶和酸性蛋白酶发酵处理花生粕48 h,粗蛋白质含量由46.4%提高至50.6%,总酸含量由0.6%提高至4.7%,且抗氧化性明显增强[68]。

表4 菌酶协同发酵高蛋白质饲料原料的效果
菌酶协同发酵可提高豆粕与高纤维饲料原料混合后的多糖含量。豆粕、玉米芯和麸皮混合后加入枯草芽孢杆菌、地衣芽孢杆菌、植物乳杆菌、酵母菌、纤维素酶、木聚糖酶、果胶酶和甘露聚糖酶36 ℃发酵72 h,木聚糖含量达1.421 mg/g[69]。豆粕、黄芪、绞股蓝和马齿苋混合后加入植物乳杆菌、枯草芽孢杆菌、纤维素酶和果胶酶,在料液比为1∶1.2(g/mL)、发酵温度35 ℃、发酵时间60 h、接种量12%的最适发酵工艺下,粗多糖含量为280.46 mg/g[70]。
4 菌酶协同发酵饲料在畜禽生产中的应用
4.1 在猪生产中的应用
菌酶协同发酵饲料能有效降低仔猪腹泻率,提高仔猪和育肥猪的生长性能,对肠道菌群组成也具有积极作用,其在猪生产中的应用效果见表5。枯草芽孢杆菌、植物乳杆菌、纤维素酶、蛋白酶、甘露聚糖酶、α-淀粉酶和葡聚糖酶协同发酵复方中草药与玉米-豆粕型饲粮,第1步是37 ℃好氧发酵20 h,第2步是37 ℃厌氧发酵5 d,将得到的菌酶协同发酵饲料饲喂杜×长×大断奶仔猪28 d,可改善断奶仔猪的生长状况,减少腹泻,提高机体免疫和抗氧化功能[71]。将植物乳杆菌、戊糖片球菌、黑曲霉、蛋白酶、淀粉酶、脂肪酶和纤维素酶发酵12 h的玉米-豆粕型饲粮饲喂杜×长×大猪21 d,可提高营养物质的消化率,改善仔猪的肠道健康,降低腹泻率,提高生长性能,且效果相当于或优于添加金霉素饲粮[72]。在1 kg玉米芯混合料加入2.5%的复合益生菌(发酵乳杆菌、酿酒酵母和枯草芽孢杆菌)和0.06%~0.08%的非淀粉多糖酶协同发酵制备发酵饲料,发酵温度25~30 ℃,发酵时间5~7 d,使用发酵饲料替代饲粮中6%的玉米和豆粕,能显著增加杜×长×大猪的日采食量、日增重和养分消化率,降低料重比,增加肠道中乳酸菌等有益菌的相对丰度,显著减少肠道和粪便中大肠杆菌等致病菌的相对丰度,提高免疫能力,显著增加回肠黏膜中细胞因子含量和分泌型免疫球蛋白A(sIgA)、白细胞介素-8(IL-8)、肿瘤坏死因子-α(TNF-α)基因表达水平,提高生长性能、屠宰性能和肉品质,改善其肠道微生态平衡和免疫力[73]。芽孢杆菌、粪肠球菌和复合酶制剂厌氧发酵玉米-豆粕型饲粮48 h,可降解饲料中80%的大豆抗原蛋白,且乳酸含量达到122.18 mmol/kg,替代10%基础饲粮后饲喂杜×长×大仔猪24 d,采食量和平均日增重提高[74]。
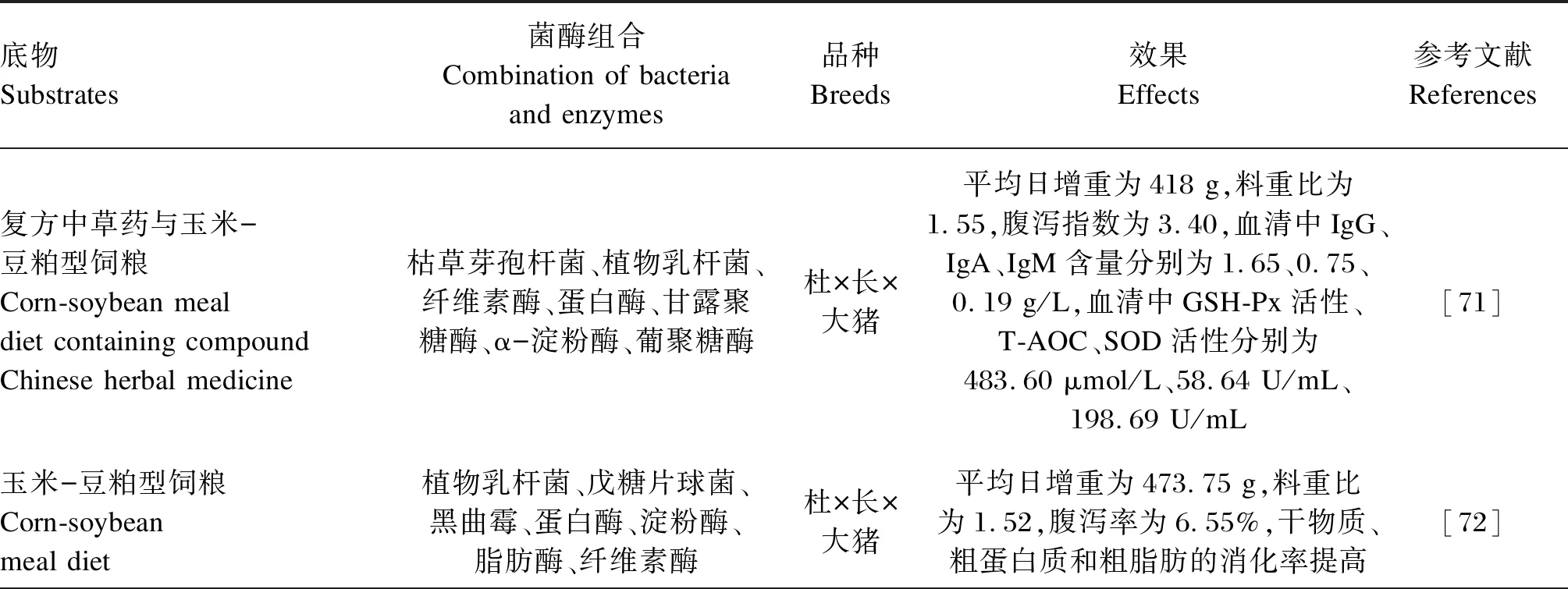
表5 菌酶协同发酵饲料在猪生产中的应用效果
4.2 在反刍动物生产中的应用
使用菌酶协同发酵高纤维饲料原料饲喂反刍动物,一方面可提高营养物质的表观消化率,另一方面可补充瘤胃益生菌,具体效果见表6。将鼠李糖乳杆菌和枝顶花纤维素酶添加到柱花草中发酵,能提高青贮饲料的发酵品质,尼龙袋法测定海南黑山羊对菌酶协同青贮柱花草的瘤胃降解率,结果表明该菌酶组合可能具有有益的协同作用[75]。植物乳杆菌、布氏乳杆菌、纤维素酶、木聚糖酶、β-葡聚糖酶、果胶酶和漆酶密封发酵水稻秸秆45 d,配制精粗比为5∶5的全混合日粮,饲喂肉羊68 d,结果表明菌酶制剂的合理组合有利于促进肉羊对营养物质的消化利用,其效果达到或超过羊草水平[76]。枯草芽孢杆菌、酵母菌、纤维素酶、葡聚糖酶和木聚糖酶塑封袋密封发酵金针菇菌糠12 d,在精料补充料中添加30%,饲喂湖羊60 d,可提高湖羊的生长性能、营养物质表观消化率和抗氧化能力[77]。使用乳酸菌和纤维素酶发酵蜡熟期全株玉米,将其饲喂荷斯坦经产奶牛70 d,奶牛的采食量和泌乳量提高,牛乳品质和奶牛机体免疫功能得到改善[78]。此外,使用益生菌(动物乳杆菌、费氏丙酸杆菌、地衣芽孢杆菌、枯草芽孢杆菌、屎肠球菌)和酶(淀粉酶、半纤维素酶和木聚糖酶)在模拟高产奶牛瘤胃环境的发酵罐中参与干燥粉碎的青贮玉米秸秆的消化,发酵罐发酵条件为39 ℃、连续注入氮气维持厌氧环境,3.05 mL/min的恒定速率注入人工唾液,丁酸盐的摩尔比例增加,而丙酸盐的摩尔比例下降,但没有导致体外瘤胃发酵发生重大改变[79]。
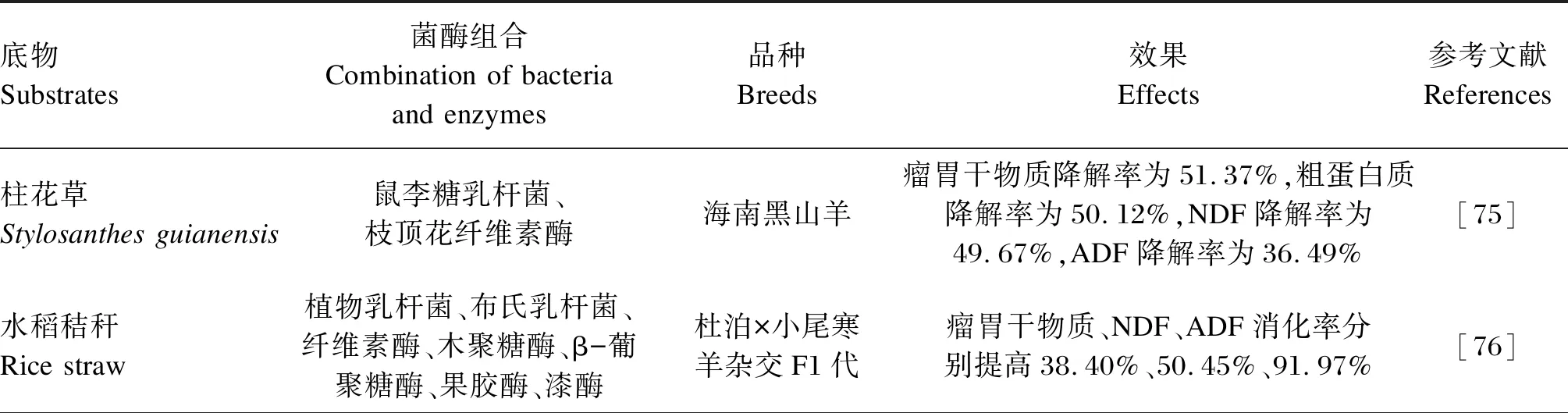
表6 菌酶协同发酵饲料在反刍动物生产中的应用效果
4.3 在家禽生产中的应用
菌酶协同发酵饲料在家禽生产中的应用效果见表7。给罗斯308肉鸡饲喂添加益生菌(包括罗伊氏乳杆菌、唾液乳杆菌、屎肠球菌、动物双歧杆菌、乳酸片球菌)+木聚糖酶组合的饲粮,饲喂21 d后,料重比和血清非酯化游离脂肪酸含量降低,肝糖原含量升高[80]。将植物乳杆菌、酿酒酵母、甘露聚糖酶、纤维素酶和α-半乳糖苷酶添加在呼吸袋中,30 ℃协同发酵棕榈仁粕3 d,粗蛋白质含量提高了15.10%,粗纤维和NDF含量分别降低了25.10%和21.96%,17种氨基酸总量增加了7.12%,将最佳条件下发酵5 d的棕榈仁粕替代基础饲粮的15%后饲喂爱拔益加(AA)肉鸡7 d,其表观代谢能和16种氨基酸的标准回肠消化率增加[81]。使用乳酸菌、酵母菌、芽孢杆菌、木聚糖酶、β-葡聚糖酶、甘露聚糖酶、纤维素酶和果胶酶协同发酵玉米-豆粕型饲粮5~7 d,将其饲喂河田肉鸡90 d,结果表明饲粮中添加5%菌酶协同发酵饲料有利于提高鸡肉风味物质含量,加强了氨基酸、次级产物及免疫功能等代谢通路基因的上调差异表达,改善了肠道菌群结构,对病原菌免疫、肠道健康起到了一定的作用[82]。

表7 菌酶协同发酵饲料在家禽生产中的应用效果
菌酶协同发酵饲料饲喂蛋鸡能提高鸡蛋品质,降低胆固醇含量。将乳酸杆菌、酵母菌、纤维素酶和蛋白酶协同发酵的饲料等蛋白质替代25%的豆粕蛋白质,饲喂京红蛋鸡49 d,改善了其生产性能和蛋品质,提高了养分表观消化率,降低了粪中氨气排放[83]。使用乳酸菌、酵母菌、芽孢杆菌和非淀粉多糖酶发酵配合饲料,发酵菌株提前用红糖水活化,发酵原料与1%复合多维、0.07%非淀粉多糖酶和活化好的复合菌株共同混合,补充水分至30%,装入带单向透气孔的聚乙烯(PE)膜袋中密封,25 ℃发酵5~7 d,将该菌酶协同发酵饲料等量替代基础饲粮的5%,饲喂商品蛋鸡346 d,料蛋比、破蛋率以及蛋黄中脂肪和胆固醇含量显著降低,产蛋率、蛋黄比率和蛋壳强度显著提高,蛋黄中总饱和脂肪酸含量提高了4.75%,总单不饱和脂肪酸含量提高了2.63%,总多不饱和脂肪酸含量提高了18.24%[84]。
5 小 结
豆粕减量替代行动是国家粮食安全中的重要环节,非常规饲料原料的开发利用和现有饲料原料的高值化利用是替代畜禽饲粮中豆粕的主要途径,而利用益生菌(多种微生物)和工业生产的酶制剂协同青贮或发酵饲料原料是实现饲料原料高值化利用的重要方法,乳杆菌属、芽孢杆菌属、木质素过氧化物酶、纤维素酶和木聚糖酶等是常用的菌酶协同组合。反刍动物生产中主要使用的是高纤维饲料原料的菌酶协同发酵,单胃动物和家禽生产中主要使用的是全混合日粮或蛋白质饲料原料的菌酶协同发酵。菌酶协同发酵单一饲料原料或配合饲料可提高饲料营养价值和改善适口性,对畜禽生长性能和肠道微生物群落组成也具有积极作用。但菌酶协同发酵饲料的最适菌酶组合及其对畜禽生长性能和肠道微生物群落改变的具体机制还有待深入研究。
