5种柏科植物叶绿体基因组密码子偏好性分析
2024-03-18黄思琦张麒功叶泽霖覃兴化汪其双林协全郑智位林文锋邹小兴
黄思琦, 张麒功, 叶泽霖, 覃兴化, 汪其双, 林协全, 郑智位, 林文锋, 邹小兴
(1.福建农林大学林学院,福建 福州 350002;2.自然生物资源保育利用福建省高校工程研究中心, 福建 福州 350002;3.福建省德化葛坑国有林场,福建 德化 362509)
秃杉(Taiwaniaflousiana)为柏科(Cupressaceae)台湾杉属(Taiwania)多年生乔木。其树干笔直,生长迅速,木纹美观,质地适中,木质优良,易于加工,是优秀的用材树种和景观树种,具有较高的经济、观赏价值[1]。由于长期的乱砍滥伐和繁育技术问题,其自然分布十分稀少[2]。在我国西南地区,秃杉结果较晚,约需29年才能收获种子[3]。然而,在闽南地区的生产实践中,约31年的秃杉人工林均未见结果,这极大增加了利用有性繁殖进行良种繁育的难度与成本。因此,开展秃杉无性快速繁育的研究十分必要。
嫁接是植物无性繁殖中常用的一种方法,广泛应用于良种保存、快速繁殖和提高产量等方面,能有效减少由性状分离导致的优良性状丢失[4-5]。砧穗之间的亲和性是影响嫁接苗成活率、生长状况和产量的关键因素。本砧嫁接使用与接穗同种的植株作为砧木,具有高度亲和性。然而,由于闽南地区的秃杉无法结果,在生产实践中获取适用于本砧的砧木的难度和成本较高,需要从其他种类中选择适用于秃杉嫁接的砧木,即异砧嫁接。如何选择亲和性高的异种砧木,成为提高秃杉嫁接成功率的研究重点。目前,异砧嫁接的砧木筛选主要从生长和生理层面进行,例如:使用福建山樱花和樱桃作为飞寒樱的砧木进行嫁接[6],筛选出具有较高成活率、抽梢率、新梢基径和长度的福建山樱花作为优良砧木;对于杜鹃红山茶,可选择高州油茶、广宁红花油茶、岑溪软枝油茶和普通油茶作为潜在砧木,通过比较分析嫁接死亡率、嫁接苗叶片代谢产物含量、抗氧化酶活性和光合色素含量,筛选出具有良好亲和性的高州油茶作为优良砧木[7]。
生长和生理指标容易受到环境因素干扰,且存在多种不确定因素。使用稳定且不易受环境影响的指标可以更准确地筛选与接穗亲缘关系相近的异种砧木。随着生物信息学的发展,高通量测序手段和基因层面的分析逐渐被应用于物种亲缘关系的研究。叶绿体基因组因其结构稳定、相对保守等特点,常作为物种进化与遗传鉴定方面的重要依据[8-9]。Junko et al[10]对25种柏科植物的叶绿体基因matK、chlL、trnL、trnF之间的基因间区与trnL的1个内含子进行测序,构建系统发育树,进一步明确各属间的关系。Liu et al[11]通过转录组测序,探究柏科植物各属的进化关系和亲缘关系。密码子是指mRNA上的三联体核苷酸残基序列,每个三联体密码子对应一个氨基酸[12]。在生物进化过程中,自然界中许多物种存在某一种或几种特定的密码子使用频率高于其同义密码子的现象,即密码子使用偏性,又称密码子偏好性[13]。不同物种的密码子具有不同的使用偏好性。有研究表明,亲缘关系较近的物种通常具有相似的密码子使用偏好性[14]。因此,从叶绿体基因组及密码子偏好性角度对秃杉砧木进行筛选,相较于生长和生理角度的筛选更为准确,从而为秃杉无性繁育研究和规模化生产提供理论基础。
本试验使用秃杉与4种待选为砧木的柏科植物,即水杉(Metasequoiaglyptostroboides)、杉木(Cunninghamialanceolata)、福建柏(Fokieniahodginsii)和日本柳杉(Cryptomeriajaponica)的叶绿体基因组,筛选出蛋白编码序列(coding sequence, CDS),并结合密码子偏好性进行分析,研究密码子偏好性形成的主要因素和使用规律,以找出与秃杉共有最优密码子最多的物种。随后,基于叶绿体基因组密码子的相对同义密码子使用度(relative synonymous codon usage, RSCU)、全基因组序列和CDS序列构建系统进化树,为秃杉最优砧木的选择以及嫁接砧木方法的可行性研究提供依据。
1 材料与方法
1.1 序列数据获取与筛选
从NCBI数据库(https://www.ncbi.nlm.nih.gov/)下载5种柏科植物的叶绿体基因组全序列。为减少分析误差,分别筛选出符合以下条件的CDS序列:(1) 剔除重复序列;(2) 确保序列碱基数为3的倍数;(3) 确保序列长度≥300 bp;(4) 每条序列均拥有起始密码子(ATG)与终止密码子(TAG、TGA、TAA),且序列中间无终止密码子[15]。经过筛选后,所得的5种柏科植物叶绿体基因组序列信息如表1所示。

表1 5种柏科植物叶绿体基因组序列信息Table 1 Information of chloroplast genome of 5 Cupressaceae plants
1.2 序列数据分析方法
对筛选出的5种柏科植物的叶绿体基因组CDS序列进行分析[16]。通过在线分析软件CUSP分别统计各物种CDS序列的密码子总GC含量(GCall)与第1、2、3位GC含量(GC1、GC2、GC3);使用分析软件CodonW得到各基因的有效密码子数(effective number of codon, ENC)、RSCU、同义密码子第3位GC含量(GC3s)以及密码子第3位A、T、C、G含量(A3、T3、C3、G3)。
在中性绘图分析[17]中,以GC3为横坐标、GC1与GC2的平均值GC12为纵坐标构建平面直角坐标系,将每个基因对应至坐标系中得到散点图,并绘制回归曲线。若散点沿坐标轴对角线分布,说明碱基突变是造成密码子偏好性的主要因素;反之则以自然选择影响为主。
在ENC-plot绘图分析[18]中,以GC3s为横坐标、ENC为纵坐标构建平面直角坐标系,ENC=2+GC3s+29/[GC3s2+(1-GC3s)2]为标准曲线。将基因定位至坐标系中得到散点图。若散点沿标准曲线分布,表明该基因密码子偏好性主要受碱基突变影响;若偏离标准曲线,表明主要受自然选择影响。
在PR2-plot绘图分析[19]中,以G3/(G3+C3)为横坐标、A3/(A3+T3)为纵坐标构建平面直角坐标系,并绘制二维散点图。以A=T、C=G为中心,根据基因落点位置判断A与T、C与G的使用频率。
在最优密码子分析[20]中,RSCU>1为高频密码子。将各序列按ENC进行升序排列,ENC较高的5个序列作为低表达基因库,较低的5个序列作为高表达基因库,进行RSCU分析并作差,所得ΔRSCU≥0.08为高表达优越密码子。同时符合高频密码子和高表达优越密码子即为最优密码子。
在聚类树与系统发育树构建[18]中,根据5种柏科植物的叶绿体基因组密码子RSCU,使用SPSS进行聚类分析,并构建聚类树。基于叶绿体基因组全序列以及CDS序列,应用MAFFT构建系统发育树。
2 结果与分析
2.1 密码子组成
秃杉、水杉、杉木、福建柏、日本柳杉的叶绿体基因组密码子的GCall分别为36.60%、37.33%、37.16%、36.75%、36.87%,密码子不同位置的GC含量均低于50%,且都呈现出GC1>GC2>GC3的趋势,即A/T含量高于G/C,同时多以A/T结尾(表2)。

表2 5种柏科植物叶绿体基因组CDS序列密码子参数特征Table 2 CDS codon parameters of chloroplast genome of 5 Cupressaceae plants
ENC是反映密码子偏好性强弱的指标,该值越大,该物种的密码子偏好性越弱,反之越强。Wu et al[21]的研究结果表明,ENC≤35时,物种基因密码子偏好性较强。由表2可得,秃杉、水杉、杉木、福建柏、日本柳杉的CDS序列密码子ENC分别为28.70~54.60、30.41~60.39、30.92~54.14、34.31~53.00、33.49~57.14,平均值分别为45.62、46.77、45.94、46.44、46.83,可见,绝大多数CDS序列的ENC高于35,表明其密码子偏好性较弱。
2.2 中性绘图分析
所有基因均落于坐标轴对角线的上方,未沿对角线分布(图1)。其中,杉木的GC12与GC3呈负相关,秃杉、水杉、福建柏和日本柳杉均呈正相关;回归系数为-0.009 7~0.292 7,均无显著相关性。可初步判定这5个物种的CDS密码子偏好性形成因素均以自然选择为主。
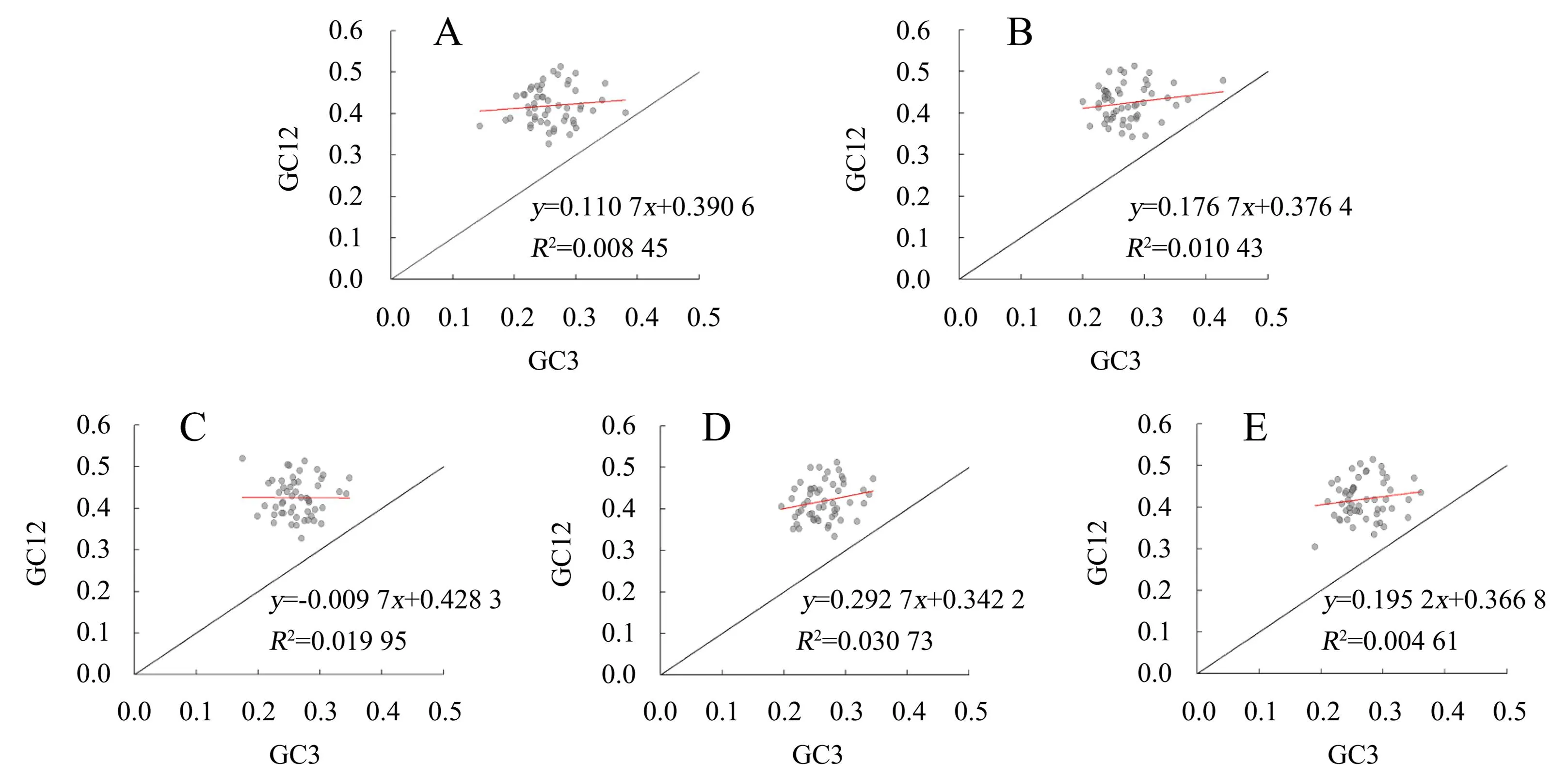
“红线”为密码子散点回归曲线。A.秃杉;B.水杉;C.杉木;D.福建柏;E.日本柳杉。
2.3 ENC-plot绘图分析
5种柏科植物的叶绿体基因组的ENC-plot图结构相似(图2),多数基因离标准曲线都具有一定距离,少部分基因沿标准曲线分布。可见,自然选择是影响多数基因密码子偏好性形成的主要因素,同时其也受到碱基突变的影响。

A.秃杉;B.水杉;C.杉木;D.福建柏;E.日本柳杉。
2.4 PR2-plot绘图分析
5种柏科植物的叶绿体基因分布均不规律(图3),其中,秃杉、水杉、杉木与日本柳杉的叶绿体基因在右下区域分布较多,福建柏则在左下区域分布较多。由此可见,这5种柏科植物叶绿体基因组密码子偏好性的形成源于碱基突变和自然选择共同作用。

A.秃杉;B.水杉;C.杉木;D.福建柏;E.日本柳杉。
2.5 最优密码子分析
根据相对同义密码子分析结果(图4),5种柏科植物叶绿体基因组共有的高频密码子有29个。而最优密码子分析结果(图5)显示,秃杉、水杉、日本柳杉、杉木、福建柏的最优密码子分别为19、17、11、13、18个。其中,密码子AAA、CCA、GCU同时为秃杉、水杉、杉木、福建柏的最优密码子;而UUA、GUA、CAU、CAA、GAU、UCU、CCU、ACU、UGU、AGA、AGU、GGU为这5种柏科植物中3种共有的最优密码子,共12个,其中4个以A结尾,8个以U结尾,没有以G/C结尾的密码子。此外,秃杉分别与水杉、杉木、福建柏、日本柳杉共有9、7、11、7个最优密码子。

图4 5种柏科植物的RSCUFig.4 Relative synonymous codon usage of 5 Cupressaceae plants
2.6 系统发育分析
根据RSCU的聚类分析结果,水杉、杉木、日本柳杉和秃杉被归为同一类群,其中,水杉、杉木和日本柳杉形成一小类,而福建柏则独立成一支(图6A)。这表明水杉、杉木和日本柳杉在密码子偏好性形成模式上相似,而秃杉略有不同。根据叶绿体基因组全序列分析结果,杉木与秃杉的亲缘关系最近,与福建柏、日本柳杉和水杉次之(图6B);根据叶绿体基因组CDS序列分析结果,杉木与秃杉的亲缘关系最为接近,与日本柳杉和水杉次之,而福建柏与其他树种的亲缘关系最远(图6C)。
3 讨论与结论
本研究对5种柏科植物的叶绿体基因组及密码子偏好性进行了分析。结果表明,这5种植物的叶绿体基因组密码子偏好性呈现为GC3>GC2>GC1,即更偏好以A/T结尾。同义密码子的使用度分析也支持这一结果。此外,ENC大于35,说明使用偏好性较弱。中性绘图分析、ENC-plot分析和PR2-plot分析的结果非常接近,表明密码子偏好性的形成源于自然选择和碱基突变共同作用。这些结果表明,由于叶绿体基因组在进化过程中具有相对稳定的结构和较低的重组率,同一科、属的植物物种的密码子偏好性及形成因素具有一致性。这与3种木莲属植物[16]、12种蔷薇科植物[17]、5种冰草属植物[22]、3种含笑属植物[23]的研究结果相似。
通过建立高、低表达基因库进行最优密码子分析,发现AAA、CCA和GCU是秃杉、水杉、杉木与福建柏这4种植物的共有最优密码子。与秃杉的最优密码子进行对比,福建柏拥有最多的共有最优密码子,共有11个,水杉为9个,日本柳杉和杉木为7个。这4种植物与秃杉共有的最优密码子数量差异较小,进一步证明了亲缘关系较近的同一科植物的叶绿体基因组密码子使用偏好性的相似性,巩固了该分析方法的可行性。
基于叶绿体基因组全序列的系统进化树、基于叶绿体基因组CDS序列的系统进化树和RSCU的聚类分析结果(D=5)均显示,秃杉与杉木的亲缘关系最为接近,与福建柏的亲缘关系最远。然而,叶绿体全基因组序列的系统进化树分析结果与RSCU的聚类分析结果略有不同,推测原因可能是全基因组中存在较多具有差异性的非编码序列。因此,可以得出结论,杉木与秃杉的亲缘关系较水杉、福建柏和日本柳杉更为接近。
嫁接是植物无性繁殖技术的重要组成部分。选择与接穗亲和性高的砧木可以有效提升嫁接成功率,保留优良性状并快速繁育良种,提高生产效益。此外,这种方法还能解决由于种子缺失导致的实生砧木缺失问题。因此,选择高亲和力的异种砧木的方法正在不断优化。近年来,随着生物信息学的发展,通过基因组分析物种间的亲缘关系可为生产实践提供重要参考。本研究通过对5种柏科植物的叶绿体基因组和密码子偏好性进行分析,并建立系统发育树验证了该分析方法的较高准确性。这可为优化砧穗亲和性选择方法提供新的理论依据,有助于推广基因组层面分析的应用,并促进珍稀植物秃杉的无性繁育研究。此外,该研究为以杉木作为秃杉的优良异种砧木提供了理论基础,也有益于秃杉资源的可持续利用。在实际生产中,杉木作为秃杉异砧嫁接的砧木,是否优于其他柏科植物种类,需要进一步研究。
