核桃炭疽病菌中GH互作蛋白找寻及遗传关系分析
2024-03-18张凯强韩长志
张凯强, 韩长志,2
(1.西南林业大学生物多样性保护学院;2.云南省森林灾害预警与控制重点实验室,云南 昆明 650224)
炭疽菌属(Colletotrichum)是植物病原真菌中的一个重要类群,主要为害花卉、蔬菜、苗木和果树等[1]。核桃(JuglansregiaL.)树是我国重要的木本油料树种,由炭疽菌属真菌引起的核桃炭疽病是该树种的常见病害。此类病菌主要侵害核桃果实,也可侵害叶片、芽和嫩梢等,侵害率高达95%以上,严重影响核桃产业的健康发展[2]。目前已报道的核桃炭疽病菌有5种,分别为胶孢炭疽菌(C.gloeosporioides)、果生炭疽菌(C.fructicola)、暹罗炭疽菌(C.siamense)、尖孢炭疽菌(C.acutatum)和葡萄炭疽菌(C.viniferum),其中,胶孢炭疽菌为主要致病菌[3-7]。
碳水化合物活性酶类(carbohydrate-active enzymes, CAZymes)是植物病原真菌中的一类重要分泌蛋白,不仅参与降解寄主植物细胞壁的过程,还能促进病原真菌在寄主植物表面的附着、侵入及在寄主体内的定殖和营养吸收等过程[8]。糖苷水解酶(glycoside hydrolases, GH)是CAZymes的重要组成类别,在病原真菌侵染植物过程中发挥着非常重要的作用。GH具有诸多亚家族,其数量和种类的差异会导致不同真菌对植物细胞壁多糖分子的降解能力不同,进而表现出不同的寄生特性[9]。如:GH7亚家族可用于降解纤维素[10];GH18可通过底物辅助保留机制催化氨基多糖中β-1,4糖苷键的生物降解[11];GH28主要参与果胶的降解过程[12]。前人对GH的研究主要集中在找寻与鉴定[13]、功能分析[14]、表达分析[15]等方面,有关植物病原真菌中GH蛋白的互作尚未见报道。
为了更好地理解核桃炭疽病菌中GH蛋白的协同作用机制,本研究基于前期所获得的胶孢炭疽菌Cg1中的GH蛋白,通过STRING网络在线分析与其相似性较高的两个物种胶孢炭疽菌Cg-14和果生炭疽菌Nara gc5相关蛋白的互作关系,同时利用MEGA X等软件对其遗传关系进行分析。
1 材料与方法
1.1 数据来源
胶孢炭疽菌Cg1由本课题组长期保存。前期开展了该菌的全基因组测序工作,并利用生物信息学工具筛选出20个GH蛋白,分别将其命名为CgCAZ1-CgCAZ20,将这20条蛋白序列通过STRING网络进行在线分析,发现有3个炭疽菌种与Cg1中的GH蛋白存在对应关系,且每个菌种中均有19个蛋白存在互作关系,选择同源性较高的胶孢炭疽菌Cg-14和果生炭疽菌Nara gc5中的相关蛋白进行互作关系分析。Cg-14和Nara gc5对应的蛋白标识符见表1。其中:Nara gc5中标识符为L2GH97的蛋白注释含有SUN(Sad-1、UNC84)结构域的家族蛋白,并不含有GH保守结构域。蛋白标识符为L2GGX8、L2FGE9、L2FZQ3、L2FX85、L2FTC1、L2G9R4、L2GH97的对应序列经Blastp比对后未发现相关蛋白,故选择相似度最高的蛋白进行后续分析,其相似度分别为90.62%、93.79%、86.93%、87.34%、91.88%、98.85%、87.44%,对应ID如表1所示。
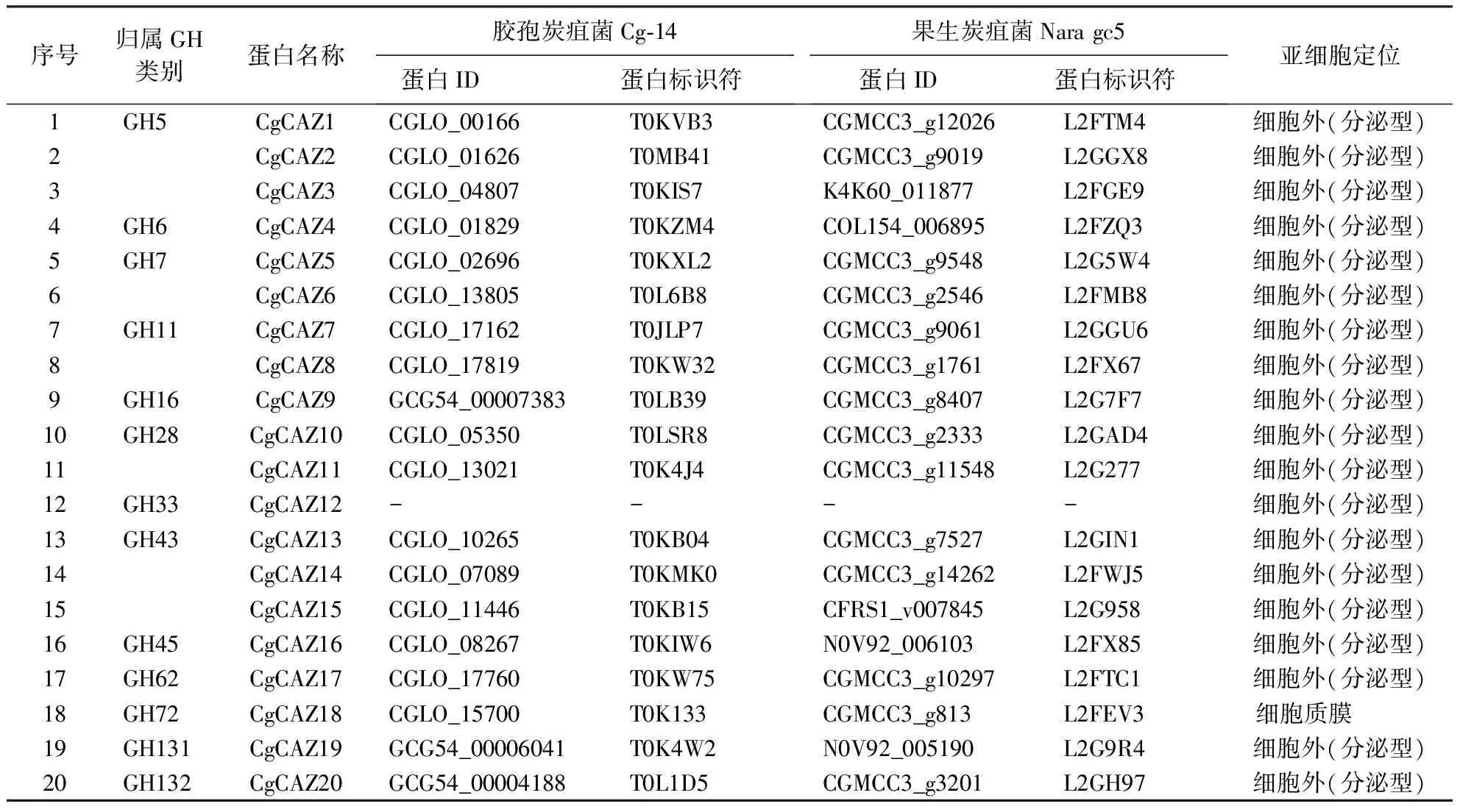
表1 不同炭疽菌中GH蛋白的基本信息1)Table 1 Basic information of GH proteins in different Colletotrichum spp.
1.2 蛋白互作分析方法
利用蛋白互作在线分析软件STRING v11.5[16]对胶孢炭疽菌Cg-14和果生炭疽菌Nara gc5中的37个GH蛋白进行分析。
1.3 分子对接分析方法
通过UniProt数据库[17]下载两种炭疽菌中相互作用关系最多的相关蛋白PDB格式文件,利用GRAMM软件[18]进行分子对接,再用Pymol软件[19]进行可视化分析。
1.4 蛋白亚细胞定位分析方法
利用亚细胞定位分析软件ProtComp v9.0[20]对3种炭疽菌中的57个GH蛋白开展分析。
1.5 蛋白遗传关系分析方法
利用Clustal X[21]和MEGA X软件[22]分别对3种炭疽菌中的57个GH蛋白进行多重比对分析和构建系统进化树。
2 结果与分析
2.1 GH蛋白互作情况预测
胶孢炭疽菌Cg-14中19个GH蛋白的互作关系主要有3种,即不同GH蛋白之间的互作、同一GH亚家族中不同蛋白之间的互作以及单独的GH蛋白(不存在互作蛋白)。以T0MB41蛋白为例,该蛋白属于GH5亚家族,其与T0KVB3、T0KW32、T0KZM4、T0KXL2、T0L6B8存在明显的互作关系,这些蛋白分别属于GH5、GH11、GH6、GH7、GH7亚家族;GH28亚家族中T0LSR8、T0K4J4两个蛋白之间存在互作关系,且不与其他蛋白互作(图1A);T0KMK0、T0K4W2、T0L1D5、T0KB15、T0KB04、T0K133、T0KIW6、T0LB39等为单独的GH蛋白。进一步逐个对上述单独GH蛋白与其他类蛋白的互作关系进行研究,发现T0KMK0、T0K4W2、T0KB15、T0KB04 4个蛋白在默认设置下找不到交互程序,需要继续降低设置严格性才能找到其互作关系。其中,T0K4W2与T0L9Q5、T0JWP1、T0K6D7、T0KZM4、T0KH50、T0LC84、T0KN70、T0L448、T0LNR6、T0K897 10个蛋白具有互作关系(图1B)。
果生炭疽菌Nara gc5中18个GH蛋白的3种互作关系与Cg-14相同。以L2GGX8蛋白为例,该蛋白属于GH5亚家族,其与L2FZQ3、L2G5W4、L2FMB8、L2FX67存在明显的互作关系,这些蛋白分别属于GH6、GH7、GH7、GH11亚家族;GH5亚家族中L2FTM4、L2FGE9两个蛋白以及GH28亚家族中L2GAD4、L2G277两个蛋白之间存在互作关系,且不与其他蛋白互作(图2A)。上述结果表明,不同亚家族的GH蛋白之间存在互作关系,且同一GH亚家族中不同蛋白之间也存在互作关系。此外,存在诸多单独的GH蛋白,主要包括L2G9R4、L2G958、L2FX85、L2FEV3、L2FWJ5、L2G7F7、L2GIN1等。进一步逐个对上述单独GH蛋白与其他类蛋白的互作关系进行研究,发现L2G9R4、L2G958、L2GIN1 3个蛋白在默认设置下找不到交互程序,需要继续降低设置严格性才能找到其互作关系。其中,L2G958与L2G384、L2GCR6、L2GEJ1、L2FKE6、L2G2H7、L2GB80、L2FC38、L2GBF8、L2FXG5、L2GIN1 10个蛋白具有互作关系(图2B)。
综上可知,Cg-14与Nara gc5中GH蛋白的互作情况极为相似,推测Cg1中GH蛋白的互作情况与这两种菌株相同。具体如下:与GH5存在互作关系的蛋白最多,且GH5亚家族中不同蛋白之间也存在互作关系;另外,GH5、GH6、GH7、GH11 4个GH亚家族之间均存在明显互作关系,GH28亚家族中两个蛋白之间互作,但不与其他GH亚家族互作;其余亚家族内部不同蛋白之间均不存在互作关系。通过逐个对单独GH蛋白的互作关系进行研究发现,单独的GH蛋白又与对应菌株中其他类蛋白存在互作关系,且与其存在互作关系的蛋白大多为CAZymes类蛋白,多参与多糖催化、碳水化合物代谢等过程。
2.2 GH蛋白分子对接情况预测
UniProt数据库中未包含现存所有蛋白的晶体结构,因此选择Cg-14和Nara gc5两种炭疽菌中互作关系最多的两种蛋白T0MB41(GH5)、T0KZM4(GH6)以及L2GGX8(GH5)、L2FZQ3(GH6)进行分子对接。通过UniProt数据库下载这4个蛋白的PDB格式文件,其晶体结构包含高置信度和低置信度所有区域(图3)。使用GRAMM软件进行分子对接,再用Pymol软件进行可视化分析。结果显示,两个蛋白之间均存在多个相互作用的氨基酸残基位点(图4),蛋白之间通过氨基酸残基相互作用,形成可以改变其生物活性的复合体,从而产生协同作用。
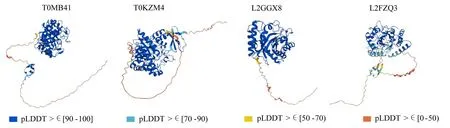
图3 4个GH蛋白晶体结构模型Fig.3 Crystal structure models of 4 GH proteins

A.T0MB41与T0KZM4分子对接晶体结构;B.T0MB41与T0KZM4相互作用氨基酸残基;C.L2GGX8与L2FZQ3分子对接晶体结构; D.L2GGX8与L2FZQ3相互作用氨基酸残基。
2.3 GH蛋白亚细胞定位情况预测
对Cg1、Cg-14、Nara gc5 3种炭疽菌中57个GH蛋白进行亚细胞定位分析,结果表明:绝大多数蛋白(54个)均定位于细胞外(分泌型);仅GH72亚家族的3个蛋白(CgCAZ18、T0K133、L2FEV3)定位于细胞质膜,其通常在细胞壁组分的交联中发挥作用[23]。STRING分析发现,T0K133、L2FEV3两个蛋白在对应菌株GH蛋白中均无明显互作关系。
2.4 GH蛋白遗传关系
对Cg1、Cg-14、Nara gc5 3种炭疽菌中57条GH蛋白序列进行多重序列比对并构建系统发育树,结果发现3种炭疽菌中GH蛋白明显分为四大类(图5)。CgCAZ14(GH43)所在分支中蛋白数量较少,仅有3个,且GH43亚家族中3个蛋白分属于不同分支,亲缘关系较远;其余3个分支中所含蛋白数量较多,分别为20、16、18个。其中:GH5中的CgCAZ1和CgCAZ3属于同一分支,而CgCAZ2单独在一个分支;GH7、GH11、GH28亚家族内部的蛋白均属于同一分支。这表明核桃炭疽病菌中GH蛋白在进化过程中总体较为稳定,少部分蛋白产生了较大程度的分化。

图5 GH蛋白的遗传关系Fig.5 Phylogenetic relationship of GH proteins
3 讨论
植物病原真菌中的分泌蛋白和CAZymes等致病因子在其生长发育及致病过程中发挥着重要作用[8]。随着学术界对核桃炭疽病菌致病因子的不断研究,G蛋白信号调控因子(regulators of G-protein signaling, RGS)[24]、聚酮合酶(polyketide synthase, PSK)[25]、漆酶[26]以及还原酶[27]等致病因子相关蛋白的生物信息学特性得到进一步解析。CAZymes是植物病原真菌侵染过程中突破寄主植物细胞壁的关键因素,主要通过其子类GH蛋白将植物多糖分解成寡糖或单糖,进而破坏细胞壁结构,突破植物表皮障碍[8]。
本研究对与Cg1相似性较高的Cg-14和Nara gc5两个菌株中37个GH蛋白的互作关系进行分析,发现其互作情况极为相似,推测Cg1中GH蛋白的互作情况与这两个菌株相同。其中,与GH5存在互作关系的蛋白最多,GH5与GH6亚家族相关蛋白之间存在的互作关系最多。GH5广泛存在于植物病原细菌与真菌中,通常参与纤维素的水解过程,在植物病原菌侵染寄主植物过程中发挥着重要作用[28]。但GH5亚家族中不同蛋白之间的互作及其与其他亚家族蛋白的互作是否可以提高纤维素水解效率进而加速侵染过程还有待进一步研究。GH5和GH28亚家族内部不同蛋白之间均存在互作关系,且存在多个单独的GH蛋白不与该菌株中其他GH蛋白互作。通过逐一对单独的GH蛋白进行分析发现,单独的GH蛋白均与其他多类蛋白存在互作关系,且这些蛋白大多为CAZymes类蛋白,此类蛋白主要参与多糖催化、碳水化合物代谢等过程。遗传关系分析发现:GH43亚家族中的3个蛋白均属于不同分支,亲缘关系较远,且CgCAZ14(GH43)所在分支中仅有3个蛋白;而GH5中的CgCAZ1和CgCAZ3蛋白属于同一分支,亲缘关系较近,CgCAZ2则与其他两个蛋白亲缘关系较远;GH7、GH11、GH28亚家族内部的蛋白均属于同一分支,亲缘关系较近。本研究预测了两种炭疽菌Cg-14和Nara gc5中GH蛋白的互作关系和遗传关系,而其他炭疽菌属真菌GH蛋白是否存在类似的互作关系,以及同一GH亚家族内不同蛋白在长期进化过程中亲缘关系远近对其功能的影响还有待进一步研究;同时,这些炭疽菌在侵染核桃过程中GH蛋白如何发挥功能,以及不同GH蛋白通过何种方式进行协同作用等均有待探索。
