狗脊多糖调控lncRNA NEAT1延缓椎间盘终板软骨细胞衰老的作用研究
2024-03-15付长龙涂海水张文键陈毅涛陆诗雨李超金灵璐郑春松
付长龙 涂海水 张文键 陈毅涛 陆诗雨 李超 金灵璐 郑春松

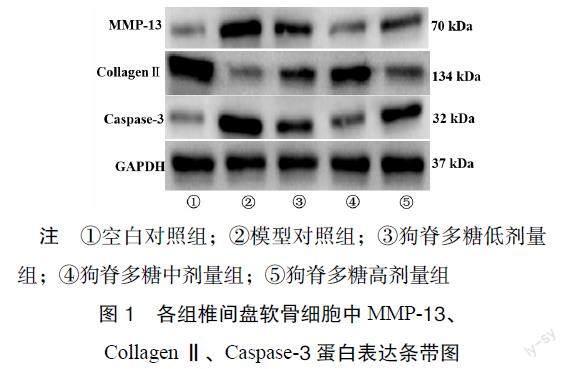
【摘 要】目的:探讨狗脊多糖通过调节lncRNA NEAT1延缓椎间盘终板软骨细胞衰老的作用机制。方法:将15只C57BL/6小鼠的椎间盘终板软骨细胞进行体外培养;经10 ng·mL-1白细胞介素-1β(IL-1β)干预24 h诱导软骨细胞;将软骨细胞随机分为空白对照组、模型对照组(10 ng·mL-1 IL-1β)和狗脊多糖各剂量组(100,200,400 μg·mL-1);Real-time PCR及Western blot分别检测狗脊多糖对IL-1β诱导软骨细胞lncRNA NEAT1、衰老相关通路p53-p21中p53、p21水平及衰老相关分泌表型的基质金属蛋白酶-13(MMP-13)、CollagenⅡ、Caspase-3蛋白表达情况;流式细胞术检测各组软骨细胞凋亡率情况。结果:Real-time PCR结果显示,经IL-1β诱导的椎间盘终板软骨细胞中lncRNA NEAT1、p21、p53水平显著增高;与模型对照组比较,狗脊多糖各剂量组lncRNA NEAT1、p21、p53水平显著降低;与空白对照组比较,经IL-1β诱导的椎间盘终板软骨细胞中MMP-13、Caspase-3蛋白表达含量显著增高,CollagenⅡ蛋白表达降低;经狗脊多糖干预后,可显著改善MMP-13、CollagenⅡ、Caspase-3蛋白表达;此外,与模型对照组比较,狗脊多糖各剂量组椎间盘终板软骨细胞凋亡率呈现下降趋势。结论:狗脊多糖通过调控lncRNA NEAT1影响p53、p21、MMP-13、CollagenⅡ、Caspase-3表达,降低软骨细胞凋亡率,进而延缓椎间盘终板软骨细胞衰老。
【关键词】 椎间盘退变;狗脊多糖;軟骨细胞;长链非编码RNA NEAT1;衰老;小鼠
The Effect of Cibotium Barometz Polysaccharides Regulating lncRNA NEAT1 to Delay the Aging of Intervertebral Disc Endplate Chondrocytes
FU Chang-long,TU Hai-shui,ZHANG Wen-jian,CHEN Yi-tao,LU Shi-yu,LI Chao,JIN Ling-lu,ZHENG
Chun-song
【ABSTRACT】Objective:To explore the mechanism of cibotium barometz polysaccharides in delaying the aging of intervertebral disc endplate chondrocytes by regulating lncRNA NEAT1.Methods:The intervertebral disc endplate chondrocytes of 15 C57BL/6 mice were cultured in vitro.10 ng·mL-1 of interleukin-1β(IL-1β)was used for intervention for 24 hours to induce chondrocytes.Chondrocytes were randomly divided into a blank control group and a model control group(10 ng·mL-1 IL-1β)and different dosage groups of cibotium barometz polysaccharides(100,200,400 μg·mL-1).Real time PCR and Western blot were used to detect the effect of cibotium barometz polysaccharides on IL-1β induced lncRNA NEAT1,the levels of p53,p21 in aging related pathway p53-p21 and the expression of aging related secretory phenotype MMP-13,
CollagenⅡ,and Caspase-3 protein.Flow cytometry was used to detect the apoptosis rate of chondrocytes in each group.Results:Real time PCR results showed that the levels of lncRNA NEAT1,p21 and p53 in IL-1β induced intervertebral disc endplate chondrocytes were significantly increased;compared with the model control group,the levels of lncRNA NEAT1,p21,and p53 in each dose group of cibotium barometz polysaccharides were significantly reduced;compared with the blank control group,the expression levels of MMP-13 and Caspase-3 proteins in IL-1β induced intervertebral disc endplate chondrocytes were significantly increased,while the expression of CollagenⅡ protein was reduced.After intervention with cibotium barometz polysaccharides,the expression of MMP-13,CollagenⅡ,and Caspase-3 proteins could be significantly improved.In addition,compared with the model control group,the apoptosis rate of intervertebral disc endplate chondrocytes in the cibotium barometz polysaccharide group showed a decreasing trend.Conclusion:Cibotium barometz polysaccharides affect the expression of p53,p21,MMP-13,Collagen Ⅱ,and Caspase-3 by regulating lncRNA NEAT1,reducing the apoptosis rate of chondrocytes and delaying the aging of intervertebral disc endplate chondrocytes.
【Keywords】 intervertebral disc degeneration;cibotium barometz polysaccharides;chondrocytes;lncRNA NEAT1;aging;mice
脊柱关节处椎间盘退变是引起患者躯干部疼痛的重要原因之一。随着椎间盘退变的加剧可导致多种脊柱关节疾病的发生、发展[1]。鉴于软骨终板为椎间盘(缺乏血供组织)提供了绝大多数营养,一旦软骨终板受不良因素(异常力学、炎症等)诱导,可破坏其正常结构与功能稳态,进而导致椎间盘退变。细胞衰老通常是机体应对细胞损伤后的应激反应,但在过度损伤及年龄增长等因素刺激下,原有的衰老-清除-再生程序发生失衡,加剧细胞出现或进入周期阻滞状态,同时诱导衰老相关标志物表达异常及衰老相关通路(例如p53-p21、p16等)处于激活状态,对机体退变产生重要影响[2-3]。终板软骨细胞衰老在加剧椎间盘退变过程中扮演了重要角色[4]。另有研究发现,长链非编码RNA(lncRNA)NEAT1在调控骨关节疾病的进程中发挥了重要作用[5]。
课题组既往研究证实,狗脊多糖可改善关节软骨细胞中Ras、Raf、MEK1/2、ERK1/2含量及一氧化氮(NO)、超氧化物歧化酶(SOD)水平,延缓椎间盘软骨细胞退变[6]。此外,狗脊多糖可影响基质金属蛋白酶(MMPs)-3、解整合素金属肽酶-4、解整合素金属肽酶-5、聚集蛋白多糖表达,维持软骨细胞外基质趋于稳态[7]。因此,本研究从lncRNA NEAT1调控细胞衰老角度,进一步探讨狗脊多糖对软骨细胞衰老相关通路及衰老相关分泌表型因子调控的作用机制。
1 实验材料
1.1 实验动物 健康SPF级3周龄C57BL/6小鼠15只,雄性,由上海斯莱克实验动物有限责任公司提供,实验动物生产许可证号SCXK(沪)
2017-0005;清洁级医学实验动物环境设施由福建中医药大学实验动物中心提供,实验动物使用许可证号SYXK(闽)2019-0007。动物处置及实验方法符合福建中医药大学实验动物伦理委员会要求(伦理批号FJTCM IACUC-2020048)。
1.2 主要试剂 MMP-13一抗(Proteintech,批号00089106);Caspase-3一抗(Proteintech,批号00092560);CollagenⅡ(Proteintech,批号00124977);GAPDH兔抗(Proteintech,批号00098110);qPCR反应试剂盒(Vazyme,批号7E410L0);PCR引物设计与合成(福州尚亚生物)。
1.3 主要仪器 DNA扩增仪(PE,型号9600);凝胶成像系统(Bio-RAD,型号GELDOC2000);CO2恒温培养箱(上海一恒科技,型号BB16/BB5060);倒置相差显微镜(OLYMPUS,型号IX70)。
2 方 法
2.1 狗脊多糖的制备 狗脊多糖制备与浓度使用参照本课题组前期实验[7]。称取狗脊药材50 g,依次经粉碎与筛检、脱脂,重复提取2次(均为10倍体积蒸馏水·次-1/(2 h)-1,收集提取液2次,抽滤减压,浓缩体积至100 mL;加入无水乙醇沉淀过夜,离心机以1000 r·min-1离心20 min,离心半径10 cm;将分离出的沉淀再进行冻干,粗多糖浸泡后再溶于蒸馏水(200 g·L-1),采用Sevag法除蛋白質,放于超低温冰箱保存。
2.2 软骨细胞的分离培养、造模 15只C57BL/6小鼠经异氟烷(体积分数为5%)麻醉后脱颈处死,逐层解剖至终板软骨组织层,分离并收集软骨组织,后依次经PBS洗净、修块、胰蛋白酶(质量分数为0.25%)消化、PBS再漂洗、Ⅱ型胶原酶溶液(质量分数为0.2%)消化等操作,将含有终板软骨细胞的悬液接种于培养皿中,转移至培养箱中孵化培养、传代[7]。IL-1β(10 ng·mL-1)诱导12 h制备软骨细胞衰老模型[8]。
2.3 Real-time PCR检测各组软骨细胞中lncRNA NEAT1、p53、p21相对表达水平 将培养好的软骨细胞分为空白对照组,模型对照组和狗脊多糖低、中、高剂量组。狗脊多糖各剂量组细胞在加入10 ng·mL-1 IL-1β干预12 h后,分别更换100,200,400 μg·mL-1 狗脊多糖继续培养48 h[6]。提取各组软骨细胞总RNA(Trizol法),逆转录成cDNA,待Real-time PCR反应(反应条件为95 ℃预变性30 s;95 ℃变性10 s,60 ℃退火30 s持续40个循环;熔解曲线设置为65~95 ℃,梯度升温,每5秒升高0.5 ℃)完成后,对各组基因表达结果采用2-ΔΔct法分析。Real-time PCR相关引物见表1。
2.4 Western blot检测各组软骨细胞的MMP-13、CollagenⅡ、Caspase-3蛋白含量 待狗脊多糖干预后,提取总蛋白,依次经BCA定量分析、配制分离胶和浓缩胶、上样、电泳、转膜、封闭,孵育一抗、二抗后进行显影、成像,分析数据。
2.5 流式细胞术检测狗脊多糖干预后软骨细胞中凋亡率情况 采用AnnexinⅤ-FITC/PI双染法检测。用胰酶(无EDTA)消化收集软骨细胞,严格按照试剂盒(Annexin-Ⅴ-FITC/PI)说明书操作,于流式管加入100 μL Binding buffer(1×),将FITC和PI各5 μL混匀(避光反应15 min),再加入400 μL Binding buffer(1×)充分混匀。
2.6 统计学方法 采用SPSS 26.0软件进行统计分析。计量资料以表示,采用t检验与单因素方差分析;计数资料采用χ2检验。以P < 0.05为差异有统计学意义。
3 结 果
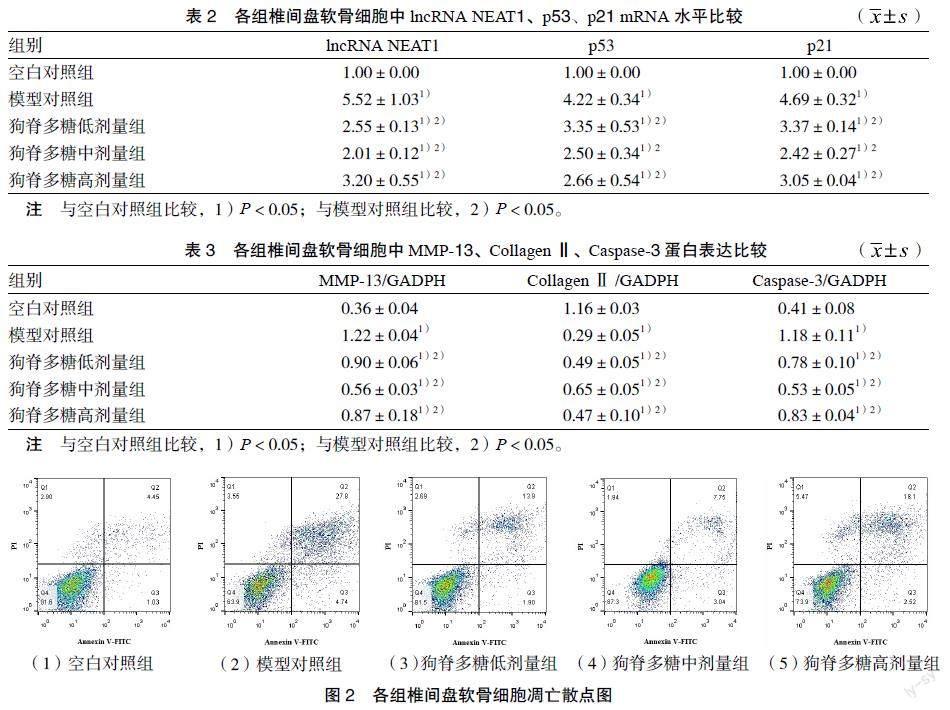
3.1 各组椎间盘软骨细胞中lncRNA NEAT1、p53、p21 mRNA水平比较 Real-time PCR结果显示,与模型对照组比较,狗脊多糖各剂量组lncRNA NEAT1、p53、p21水平显著降低(P < 0.05)。见表2。
3.2 各组软骨细胞中MMP-13、CollagenⅡ、Caspase-3蛋白表达比较 Western blot结果显示,狗脊多糖作用于IL-1β诱导软骨细胞后,可改善MMP-13、CollagenⅡ、Caspase-3蛋白含量表达。与模型对照组比较,狗脊多糖各剂量组的MMP-13、Caspase-3蛋白含量表达显著降低,而CollagenⅡ显著增高(P < 0.05)。见图1、表3。
3.3 各组椎间盘软骨细胞凋亡率比较 采用流式细胞术检测细胞凋亡率。与空白对照组比较,模型对照组软骨细胞凋亡率显著增高(P < 0.05);与模型对照组比较,狗脊多糖各剂量组软骨细胞凋亡率显著降低(P < 0.05)。见图2、表4。
4 讨 论
脊柱关节处椎间盘退变属中医学“痹证”范畴,即其病位在“肝肾”,然病在“筋骨”。随着肝肾日渐亏虚,引起筋骨失养、腠理空虚,正气不足以驱逐外邪,故风寒湿邪滞留关节而发病。因此,补肾柔肝、除痹消痛是其重要治则[9]。中药狗脊始载于《神农本草经》,其性温,味苦甘,入肝肾经,具有补肝肾、壮筋骨之效,切合痹证的治疗病机,是临床用于治疗脊柱关节疾病的常用药物之一[10]。狗脊多糖是中药狗脊的重要活性物质成分。既往研究证实,狗脊多糖可促进软骨细胞增殖,抑制氧化应激及软骨细胞凋亡[11-13]。
近年来,大量证据表明,骨关节疾病的发生、发展与相关衰老细胞数量的增加密切相关[14]。一旦终板软骨细胞经异常因素诱导发生衰老,其细胞合成蛋白质能力减弱,导致软骨细胞外基质稳态失衡,造成软骨终板结构紊乱,进而加剧脊柱关节退变[15]。另有研究发现,衰老相关分泌表型(MMPs等)表达异常加剧了终板软骨细胞衰老[16]。Ⅱ型胶原作为构成细胞外基质中胶原蛋白成分的主要物质,一旦发生异常降解,可引起软骨的抗压与弹性功能下降[17]。MMP-13作为降解Ⅱ型胶原的关键酶类之一,可进一步加重软骨退变与衰老[18]。已有研究证实,IL-1β(10 ng·mL-1)可诱导终板软骨细胞发生衰老,而衰老的终板软骨细胞中相关衰老标志物p53、p21蛋白呈现表达异常增高状态[8]。
本研究結果亦发现,10 ng·mL-1 IL-1β诱导终板软骨细胞后,其p53、p21水平及MMP-13、Caspase-3蛋白含量显著增高,而Collagen Ⅱ含量显著降低。经狗脊多糖干预后,可显著降低IL-1β诱导的软骨细胞中p21、p53基因水平,并可改善MMP-13、Collagen Ⅱ、Caspase-3蛋白表达,并降低软骨细胞凋亡率。究其原因,狗脊多糖的作用机制可能与调控lncRNA NEAT1密切相关。lncRNA NEAT1已被证实在退变软骨中上调表达,并通过促进软骨细胞凋亡加剧软骨退行性变[19]。此外,lncRNA NEAT1的异常表达与衰老相关信号分子p53的转录调控密切相关[20]。本研究结果表明,经IL-1β诱导的软骨细胞中lncRNA NEAT1水平增高,与p53变化具有一致性,进而提示lncRNA NEAT1可能参与了对终板软骨细胞衰老的调控;经狗脊多糖干预后,lncRNA NEAT1水平显著降低。
综上所述,狗脊多糖可通过影响p53、p21、MMP-13、CollagenⅡ、Caspase-3表达,降低软骨细胞凋亡率,进而延缓椎间盘终板软骨细胞衰老,其作用机制与调控lncRNA NEAT1有关。然而,本研究当前仍存在不足,例如狗脊多糖调控何种与lncRNA NEAT1相关的竞争性内源RNA途径发挥延缓软骨衰老的深层次作用机制仍需做进一步探讨。
参考文献
[1] 朱超,阮狄克.椎间盘退变机制研究进展[J].中国骨与关节杂志,2022,11(9):700-706.
[2] 张艳琳,黄国付,邹璟,等.终板软骨细胞衰老在腰椎间盘退变中的研究进展[J].医学研究杂志,2022,51(7):173-176,134.
[3] HERRANZ N,GIL J.Mechanisms and functions of cellular senescence[J].J Clin Invest,2018,28(4):1238-1246.
[4] BUCKWALTER JA.Aging and degeneration of the human intervertebral disc[J].Spine,1995,20(11):1307-1314.
[5] LIU F,LIU X,YANG Y,et al.NEAT1/miR-193a-3p/SOX5 axis regulates cartilage matrix degradation in human osteoarthritis[J].Cell Biol Int,2020,44(4):947-957.
[6] 付长龙,梅阳阳,李民,等.基于Ras-Raf-MEK1/2-ERK1/2信号通路探究狗脊多糖延缓大鼠椎间盘软骨细胞退变[J].中华中医药杂志,2019,34(7):3311-3314.
[7] 付长龙,梅阳阳,金灵璐,等.狗脊多糖调控lncRNA GAS5维持软骨细胞外基质稳态的机制[J].中华中医药杂志,2022,37(12):7379-7383.
[8] 周年,胡侦明,林鑫,等.SIRT1去乙酰化修饰p53抑制IL-1β介导的软骨终板细胞衰老[J].第三军医大学学报,2018,40(22):2040-2046.
[9] 苑珍珍,杨召,许海委.中药抑制椎间盘退变的分子机制研究[J].中国矫形外科杂志,2022,30(1):44-47.
[10] 付长龙,梅阳阳,李民,等.狗脊多糖对硝普钠诱导退变大鼠软骨细胞氧自由基影响的研究[J].风湿病与关节炎,2018,7(6):5-9,14.
[11] 孙群周.狗脊多糖通过miR-181c调控IL-1β介导的骨关节炎软骨细胞增殖和凋亡[J].中国老年学杂志,2021,41(11):2398-2402.
[12] HUANG D,HOU X,ZHANG D,et al.Two novel polysaccharides from rhizomes of Cibotium barometz promote bone formation via activating the BMP2/SMAD1 signaling pathway in MC3T3-E1 cells[J].Carbohydr Polym,2020,231(231):115732-115746.
[13] FU C,ZHENG C,LIN J,et al.Cibotium barometz polysaccharides stimulate chondrocyte proliferation in vitro by promoting G1/S cell cycle transition[J].Mol Med Rep,2017,15(5):3027-3034.
[14] 张广智,武作龙,贺学岗,等.细胞衰老与椎间盘退变的相关性研究进展[J].生命科学研究,2021,25(1):58-63,94.
[15] 徐偉,廖冬发,王娟,等.细胞衰老在骨关节炎中作用的研究进展[J].中国矫形外科杂志,2022,30(15):1386-1390.
[16] 赵继荣,杨正汉,马俊飞,等.中医药干预基质金属蛋白酶表达治疗椎间盘退变研究进展[J].中国实验方剂学杂志,2023,29(5):272-282.
[17] KUMAVAT R,KUMAR V,MALHOTRA R,et al.Biomarkers of joint damage in osteoarthritis:current status and future directions[J].Mediators Inflamm,2021(9):1-15.
[18] HU Q,ECKER M.Overview of MMP-13 as a promising target for the treatment of osteoarthritis[J].Int J Mol Sci,2021,22(4):1742-1762.
[19] FU C,QIU Z,HUANG Y,et al.Achyranthes bidentata polysaccharides alleviate endoplasmic reticulum stress in osteoarthritis via lncRNA NEAT1/miR-377-3p pathway[J].Biomed Pharmacother,2022,154(1):113551-113564.
[20] MA T,LI H,LIU H,et al.Neat1 promotes acute kidney injury to chronic kidney disease by facilitating tubular epithelial cells apoptosis via sequestering miR-129-
5p[J].Mol Ther,2022,30(10):3313-3332.
收稿日期:2023-10-08;修回日期:2023-11-15
