陆地棉HD-Zip 家族全基因组鉴定及响应非生物胁迫的表达分析
2024-03-12吴翠翠肖水平
吴翠翠 肖水平
(1.山西农业大学棉花研究所,运城 044000;2.江西省经济作物研究所,南昌 330000;3.棉花生物育种与综合利用全国重点实验室,安阳 455000)
棉花作为我国主要经济作物和主要天然纤维来源,已成为我国除粮食以外不可缺少的战略储备物资,但部分地区土壤盐碱化严重、极端高温、干旱天气和“倒春寒”现象限制了棉花产量和质量。因此,解析棉花抗逆分子机制,将有助于通过遗传育种的方式,培育高产优质抗逆的棉花新品种。近年来,随着多个棉属物种基因组测序工作的相继完成[1-8],在基因组水平上揭示了棉花的进化机制和棉花资源遗传多样性的原因,为棉花产量、品质和抗性的提高提供了丰富的基因组学资源。
HD-Zip 家族蛋白是植物特有的转录因子,编码含有60 个保守氨基酸同源结构域(HD)的同源异型盒(Homeobox)蛋白。在植物中,第一个含有HD 的基因在玉米(Zea mays)中被报道,发现了一个控制叶片分化机制的Knotted1[9]。目前,在拟南芥、水稻、小麦、玉米、马铃薯、大麦、番茄、向日葵和苜蓿中均发现了该基因[10-17]。HD-Zip 蛋白包括HD 结合结构域和与HD 紧密相连的亮氨酸拉链(Zip)结构域,并介导蛋白质二聚体的形成[10]。根据HD-Zip 的结构特点、序列保守性、理化功能等特征,将其分为4 个亚家族,即HD-ZipI-IV[10,18]。研究表明,HD-Zip 家族基因在许多植物中具有重要的功能。
HD-ZipI 亚家族主要在逆境胁迫、叶片发育和光信号转导中发挥重要作用[19]。拟南芥HD-Zip I 亚家族中参与低温和干旱胁迫的AtHB6、AtHB7、AtHB12通过关闭气孔、增加水分的吸收和调节相应的胁迫基因来抵抗胁迫的影响[20-22]。在水稻HD-ZipI 亚家族中,干旱处理下过表达Oshox4 可以减少节间伸长,延长营养生长期;Oshox4 还可以调节GA 的信号转导[11]。此外,Oshox22 能够通过ABA 介导的信号转导途径影响ABA 的生物合成,调节干旱和盐分响应[23]。
HD-ZipII 蛋白参与光响应、避光和生长素信号传导[24]。研究表明,AtHB2 参与光响应信号和通过生长素途径调节植物结构[25-26]。此外,避荫反应的负调控因子HFR1/SICS1 调控AtHB2 的表达,使植物在被遮蔽时能保持各方面的稳态[27]。HAT2、AtHB17 均参与生长素信号的传导过程[28-29]。
HD-ZipIII 蛋白参与调控侧向器官起始、胚胎发生、叶极性和分生组织等功能[24]。拟南芥HDZipIII 类中只 有5 个成员:AtHB8、PHAVOLUTA/AtHB9、PHABULOSA/AtHB14、CORONA/AtHB15 和REVOLUTA/IFL1[29]。REV、PHV 及PHB 在胚胎 发育过程中控制顶端形成的模式[30],此外,PHB 和PHV 通过抑制Kanadi 导致侧器官的黄化以及木质部的形成[31]。在诱导木质部分化方面,AtHB8 也可能发挥作用[32]。
HD-ZipIV 蛋白在表皮细胞分化、毛状体形成、花青素积累和根发育过程中发挥重要作用[24]。AtGL2 是最早被证明对表皮毛分化有重要作用的一个基因[33]。AtPDF2 和AtML1 也被证明参与表皮细胞分化过程[34]。研究表明,ANL2 在控制叶片花青素沉积、细胞壁机械特性调节中起作用[35]。此外,该亚家族在棉花纤维发育中研究较多。GhHD1 通过WRKY 转录因子和钙信号通路基因来调节乙烯和ROS(活性氧)的水平,从而正调控棉花纤维的起始[36]。海岛棉中的同源基因GbHD1 通过转座子的插入导致茎毛数量的减少[37]。陆地棉中Tyl 类型的反转录转座子优先插入GhHD1 的A 亚组使棉花茎毛数量减少[38]。GaHOX1 在gl2 突变体拟南芥中过表达可以恢复其表皮毛缺失表型[39]。GhHOX3 与GhHD1 互作激活下游基因GhRDL1 和GhEXPA1 的表达进而促进纤维细胞的伸长[40]。
棉花中关于HD-Zip 家族基因在非生物胁迫中的研究鲜有报道。本研究从全基因组范围系统鉴定陆地棉中HDZ 基因家族成员,对GhHDZ 家族进行系统的生物信息学分析,并基于已建立的陆地棉转录组数据分析该家族响应冷、热、盐、干旱胁迫的表达特征,通过RT-qPCR 对部分GhHDZ 基因在4 种非生物胁迫处理下的表达模式进行鉴定,并对GhHDZ146 进行了功能分析,为陆地棉GhHDZ 家族基因在响应逆境胁迫等方面的功能研究奠定基础。
1 材料与方法
1.1 材料
采用陆地棉品种TM-1 种植在培养箱中,条件是16 h 光照/8 h 黑暗,温度26-28℃。在生长至三片真叶时进行非生物胁迫,分别是冷(4℃)、热(40℃)、盐(250 mmol/L NaCl)和 干旱(30%PEG6000),对照为水处理。在处理后的3、6、12和24 h 收集叶片冷冻在液氮中,-80℃保存,每个样品取3 个独立的生物学重复。
1.2 方法
1.2.1 GhHDZ 基因家族的鉴定、理化性质及亚细胞定位预测分析 使用CottonFGD(https://cottonfgd.net/)网站搜索并下载陆地棉的基因组、编码序列和蛋白质序列(ZJU,TM-1,version 2.1)。从PFAM 数据库中下载HD(Homeodomain)的隐马尔可夫模型(hidden Markov Models,HMM)文件PF00046 和PF02183,利用HMMER3.0 软件(http://www.hmmer.org),e-value 值取1e-10,初步筛选出陆地棉HDZ家族基因。使用NCBI Web Batch CD-Search Tool[41](https://www.ncbi.nlm.nih.gov/Structure/bwrpsb/bwrpsb.cgi)进一步进行筛选并鉴定GhHDZ 基因。
利用在 线网站 ExPASy[42](https://web.expasy.org/compute_pi/)分析GhHDZ 家族蛋白的氨基酸数目、蛋白质长度、等电点、分子量以及亲水性等理化性质。同时,利用WoLF-PSORT[43](https://wolfpso rt.hgc.jp/)和CELLO v2.5[44](http://cello.life.nctu.edu.tw/)网站预测GhHDZ 蛋白的亚细胞定位。
1.2.2 陆地棉GhHDZ 的系统进化树、染色体定位和基因结构分析 使用Clustal X[45]对所筛选的205 个GhHDZ 蛋白序列进行多序列比对,使用MEGA7.0 软件的邻接法[46]构建GhHDZ 家族的系统发育树。利用在线Evolview(https://www.omicsclass.com/article/671)修饰GhHDZ 的系统进化树。
使用MapChart 软件分析GhHDZ 在染色体上的分布。利用在线MEME 网站(http://memesuite.org/)用于识别保守的蛋白质基序[47]。利用在线GSDS 2.0(http://gsds.gao-lab.org/)绘制陆地棉GhHDZ 家族基因外显子-内含子基因结构图[48]。并用TBtools 软件[49]绘制系统发育树、基因结构和保守蛋白基序合图,使用来自MEME 网站的XML 文件、来自系统发育树分析的NWK 文件和陆地棉基因组的GFF3文件。
1.2.3 GhHDZ 家族基因顺式元件、共线性分析及基因复制分析 从CottonFGD 下载陆地棉GhHDZ 家族基因起始密码子上游2 kb 序列的启动子区。使用在线PlantCARE 网站(http://bioinformatics.psb.ugent.be/webtools/plantcare/htmL/)[50]预测了GhHDZ 家族基因的顺式作用元件。筛选并分析与植物生长发育、植物激素和非生物胁迫具有特定表达模式的顺式作用元件。
利用MCScanX[51]程序进行GhHDZ 家族基因的共线性分析,采用TBtools 软件绘制GhHDZ 家族同源基因对间的共线性Circos 图,并利用KaKs_Calculator 2.0 软件[52]计算同源基因对之间的非同义替换(Ka)和同义替换(Ks)的Ka/Ks 值,对各基因对之间的选择压进行分析[49]。
1.2.4 利用转录组数据分析GhHDZ 家族基因的表达模式 从NCBI 数据库下载棉花TM-1 非生物胁迫(4℃低温、37℃高温、200 mmol/L NaCl 盐胁迫、200 g/L PEG6000 溶液模拟干旱胁迫)转录组测序数据(登录号:PRJNA248163)。通过TBtools 软件生成表达热图,所有基因表达水平均通过log2(FPKM+1)标准化显示[49]。
1.2.5 非生物胁迫处理陆地棉总RNA 的提取及实时荧光定量PCR 非生物胁迫处理叶片总RNA 的提取和反转录、实时荧光定量等试验方法参照文献[53]。内参基因为GhHis3(GH_D03G0404),采用Primer3 软件设计引物(表1)。采用2-ΔΔCt方法计算数据[54]。

表1 所用相关引物Table 1 Primers used in the study
1.2.6 GhHDZ146 的亚细胞定位 克隆GhHDZ146的全长CDS 序列,并与经Sal I 和BamH I 双酶切线性化的PRI101-35S-GFP 载体连接。将上述融合载体转化到农杆菌GV3101 中,通过农杆菌介导的方法瞬时转化烟草叶片,以空载体pRI101-35S-GFP 为对照[55]。注射烟草后避光培养36-48 h,用激光共聚焦显微镜SP8(莱卡,德国)观察叶片中的GFP(green fluorescent protein)荧光。所用亚定位扩增引物详见表1。
1.2.7 过表达GhHDZ146 拟南芥的转化及盐胁迫 将上述构建的GhHDZ146 亚细胞定位载体作为过表达载体,转化到根癌农杆菌菌株 GV3101 后,使用浸花法浸染拟南芥(哥伦比亚生态型)[56]。浸染后暗处理24 h 后转移至培养箱中,在20-22℃ 16 h 光照/8 h 黑暗下生长,7 d 后再浸染一次,直至T0代种子收获。在含有Kan+(50 μg/mL)的1/2 MS 固体培养板进行T0、T1、T2、T3代种子的阳性株筛选。
选取3 个T3代过表达转基因株系以及野生型材料,在无抗1/2 MS 培养基上长至5 d 时,将长势均匀的幼苗移至新的含200 mmol/L NaCl 的1/2 MS 培养基的方皿板上,每个方皿中包含3 个过表达株系和1 个WT,3 个重复,每重复每株系移入10 颗苗,进行胁迫试验。于光照培养箱中垂直培养7 d 后,分别进行主根及根毛拍照。采用ImageJ 软件进行根长和根毛长度的测量,其中,根毛长度测量方法参考付艳茹等[57]方法进行,根毛长度测量从根部成熟区根毛生长的起始位置开始,从下往上量取5 mm长度区域内的根毛进行测量。
2 结果
2.1 陆地棉GhHDZ基因家族成员鉴定及理化性质分析
在陆地棉中共检测到205 个GhHDZ 家族基因,根据基因在染色体上分布位置,依次命名为GhHDZ1-GhHDZ205(附表1)。
通过分析GhHDZ 基因的理化性质(附表1),205 个GhHDZ 基因编码的蛋白质长度为95(GhHDZ88)-1 750 aa(GhHDZ43)。外显子数目为1-19,变异范围比较广泛,其中,含有3 个外显子的基因数目最多为75 个,而外显子数目为5、7、14、16、17 和19 含有外显子数目较少为1 或者2。等电点为4.40(GhHDZ68)-10.05(GhHDZ125),平均为6.86;GhHDZ 的亲水性为-1.21(GhHDZ109)--0.083(Gh-HDZ102),平均亲水性为-0.703,说明GhHDZ 为亲水性蛋白。相对分子量为11.154(GhHDZ88)-197.851 kD(GhHDZ43),平均相对分子量为46.647 kD。亚细胞定位预测结果表明,186 个基因定位于细胞核,12 个基因定位于叶绿体,6 个定位于细胞骨架,只有1 个基因定位于液泡中。90%以上GhHDZ 定位于细胞核,表明这些蛋白可能作为转录因子调控陆地棉的生长发育。
2.2 GhHDZ基因家族的系统发育分析
为了揭示GhHDZ 蛋白之间的系统发育关系,建立一个无根的系统发育树来评估陆地棉HDZ 之间的遗传进化关系。结果(图1)表明,GhHDZ 家族蛋白可划分为4 个亚组,分别命名为HD-ZipI、HDZipII、HD-ZipIII、HD-ZipIV。陆地棉中,HD-ZipI 亚组中含有73 个GhHDZ 基因,是含基因数量最多的一组;HD-ZipII-IV 成员的数量分别为36、59 和37(图1)。
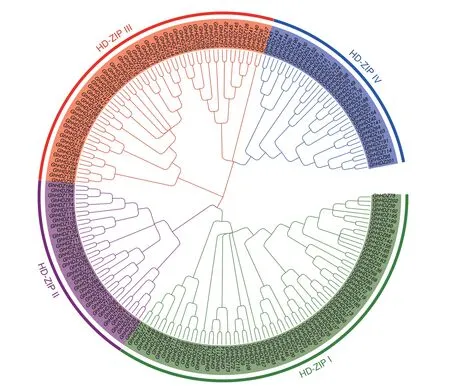
图1 陆地棉HDZ 蛋白的系统发育分析Fig.1 Phylogenetic analysis of HDZ proteins from G.hirsutum L.
2.3 GhHDZ 家族的染色体定位
为了更好地了解GhHDZ 家族的基因组分布机制,绘制GhHDZ 的染色体图谱(图2)。在At 亚基因组中分布有102 个基因,而Dt 亚基因组中分布有103 个基因,可见在At 亚基因组和Dt 亚基因组的GhHDZ 基因数目分布基本一致。分别对各染色体的基因进行分析,在Dt12 上分布了最多的GhHDZ基因(16 个),而在At12 上有14 个GhHDZ 基因;在At05 和At11 上都含 有13 个GhHDZ 基 因,以及Dt05 和Dt11 上分别含有11 和12 个GhHDZ 基因,以上是所有分布在10 个以上GhHDZ 基因的染色体。此外,在At01、Dt03、Dt10 染色体上各有9 个GhHDZ 基因;有4 个染色体上各分布有8 个GhHDZ 基因,分别是At03、At07、Dt01 和Dt08;而在At08、At10、Dt07 和Dt13 染色体上各有7 个GhHDZ 基 因;在At02、At13 和Dt06 上各有6 个GhHDZ 基因;有2 个染色体各有5 个GhHDZ 基因,分布是At06 和Dt02;在At04、At09、Dt04 上各分布3 个GhHDZ 基因,而只有Dt09 上有2 个GhHDZ基因(图2)。上述结果表明,GhHDZ 基因在GhAt和GhDt 亚组上基本呈现均匀的分布模式。

图2 GhHDZ 基因家族的染色体定位分布Fig.2 Chromosomal locations of GhHDZ gene family
2.4 GhHDZs的共线性分析和Ka/Ks分析
通过分析GhHDZ 基因的共线性关系,并鉴定陆地棉中各GhHDZ 基因的旁系同源基因对(图3)。共鉴定出377 对旁系同源基因,其中372 对为片段复制,占基因对总数的98.7%。同时检测到5 个GhHDZ 串联重复基因对(图3),分别是GhHDZ16和GhHDZ17、GhHDZ43 和GhHDZ44、GhHDZ123 和GhHDZ124、GhHDZ143 和GhHDZ144、GhHDZ165和GhHDZ166。综上分析,发现片段复制对GhHDZ家族基因的进化占据主导地位。
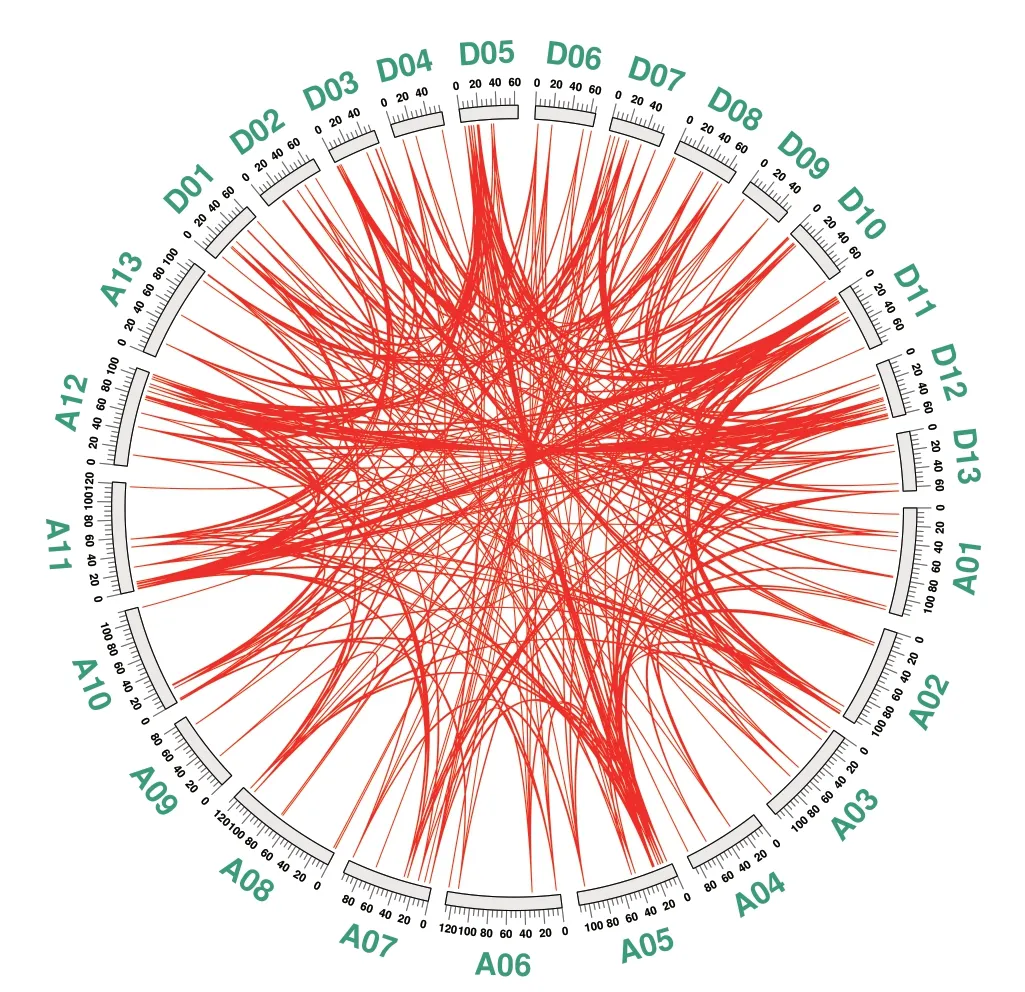
图3 GhHDZ 基因家族的共线性分析Fig.3 Syntenic relationship of GhHDZ gene family
此外,通过计算377 对GhHDZ 同源基因对的Ka/Ks 值,并根据Ka/Ks 值推测了重复基因对的选择压力。在所有377 对基因中,共有337 对有选择压力,其中,有318 个重复基因对的Ka/Ks 值<0.5,占比94.4%;有16 个重复基因对的Ka/Ks 值为0.5-1.0,占比4.7%,说明共有334 对同源基因对为负向选择,即功能同质化(附表2)。此外,GhHDZ52/GhHDZ151 基因对的Ka/Ks 值为2.0,远大于1,而GhHDZ51/GhHDZ150、GhHDZ95/GhHDZ197 基因对的Ka/Ks 值分别为1.45 和1.1,说明这些同源基因对为正向选择。上述结果说明,纯化选择对GhHDZ 的同源基因对形成至关重要(附表2)。
2.5 GhHDZ基因的保守基序和基因结构分析
为了解GhHDZ 基因家族不同成员之间的进化关系,分析GhHDZ 基因家族的保守蛋白质基序(图4)。结果表明,共发现10 个保守基序,命名为Motif 1-Motif 10,GhHDZ 家族基序种类从2-9 不等(图4-b),同一亚组内存在相似的基序分布。在HDZipIII 和HD-ZipIV 中基序种类的跨度范围都比较大,从2-9 种不等,同时这两个亚组也是含有基序数量最多的。在HD-ZipI 和HD-ZipII 中保守基序的分布比较均一,跨度从2-5 个不等。发现绝大多数GhHDZ 家族基因含有Motif 1、Motif 2 和Motif 8 基序,说明这3 个基序在GhHDZ 家族中高度保守。

图4 GhHDZ 家族的系统发育、基因结构和基序分析Fig.4 Phylogenetic,gene structure,and motif analyses of GhHDZ family
另外,通过分析GhHDZ 的基因结构,发现HDZipI 中GhHDZ 基因的外显子数目为1-4 个;HDZipII 的外显子数目多数是3 个,少数是4 个;HDZipIII 中外显子数目跨度范围最大,为2-18 个;HD-ZipIV 的外显子从7-19 不等,平均值为11,是外显子数目最多的一组(图4-c)。其中,有13 个基因没有内含子,而包含16-19 个外显子的GhHDZ 基因有22 个(图4-c)。值得注意的是,同一亚组内的基因具有相似的内含子和外显子排列,表明GhHDZ基因的外显子-内含子结构与系统发育高度相关。
2.6 GhHDZ家族的顺式元件分析
通过对GhHDZ 基因启动子上游的顺式元件进行分析,发现GhHDZ 基因的启动子区包括结构型和诱导型作用元件,可分为三大类共22 种功能元件(附表3,图5)。其中,关于生长发育类顺式元件包含了11 种功能元件,激素响应类的顺式元件只有3 种功能元件,非生物胁迫类的顺式元件有8 种功能元件(图5)。
在生长发育类别中,主要有11 个功能元件类型,包括41 种基序。其中,光反应顺式元件极其丰富,包括G-Box、ACE、MRE、GA-motif、TCT-motif等31 个基序类型,有204 个(占比99.5%)GhHDZ基因的启动子区都含有该类元件。此外,有66 个(32.2%)GhHDZ 基因的启动子区含有玉米醇溶蛋白代谢调节元件(O2-site),38 个(18.5%)含有分生组织表达元件(CAT-box),28 个(13.7%)含有昼夜节律调控元件(circadian),分别有23 个(11.2%)GhHDZ 基因的启动子区含有MYBHv1 结合位点元件(CCAAT-box)和胚乳表达元件(GCN4),18 个(8.8%)含有种子特异调控元件(RY),各有11 个(4.9%)基因含有类黄酮生物合成基因调控元件(MBSI)和栅栏叶肉细胞分化元件(HD-ZipI),10 个(4.9%)含有细胞周期调节元件(MSA-like),另有6 个(2.9%)含有最大激发子介导的激活元件(富含AT 序列)(附表S3)。
GhHDZ 家族基因中与非生物胁迫响应相关的一系列顺式作用元件包含8 种功能元件,10 种元件基序。其中,有171 个(83.4%)GhHDZ 基因的启动子区有厌氧诱导元件(ARE),有96 个(46.8%)有水杨酸应答元件(TCA 和SARE),91 个(44.4%)有MeJA 应答元件(CGTCA 和TGACG),77 个(37.6%)有干旱诱导功能(MBS),64 个(31.2%)有防御和胁迫应答元件(富含TC 基序),51 个(24.9%)有低温反应元件(LTR 基序),缺氧特异诱导元件(GC)和创伤反应元件(WUN)分别有18 个(8.8%)和12 个(5.9%)(附表3)。
与植物激素响应相关的共鉴定出3 种功能元件,包含6 种基序。其中,有120 个(58.5%)GhHDZ基因的启动子区含有脱落酸应答元件(ABRE),91个(44.4%)含有赤霉素应答元件(GARE、P-box和TATC-box),48 个(23.4%)含有生长素应答元件(AuxRE 和TGA-box)(附表3)。
总之,GhHDZ 基因的大多数成员具有光反应元件(99.5%)、厌氧诱导元件(83.4%)和脱落酸反应元件(58.5%),水杨酸应答元件(46.8%),表明这些相对丰富的元件在GhHDZ 基因家族中高度保守,在响应生长发育、激素响应和非生物胁迫反应中发挥了关键作用。
2.7 GhHDZ家族在非生物胁迫中的表达模式
利用从NCBI(登录号:PRJNA248163)下载非生物胁迫的转录组数据,研究GhHDZ 各基因的表达模式。根据进化树划分的4 个亚组GhHDZ 基因分别进行分析。
对陆地棉HD-ZipI 亚组基因冷胁迫处理的转录组分析,发现GhHDZ12、GhHDZ23、GhHDZ46、GhHDZ119、GhHDZ120、GhHDZ146 在1 h 或3 h 的表达量最高,在12 h 的表达量最低。在高温胁迫下,GhHDZ12、GhHDZ46、GhHDZ66、GhHDZ92、GhHDZ119、GhHDZ146、GhHDZ169、GhHDZ193在1 h 的表达量最低,在3、6 和12 h 的表达量逐渐变大,在12 h 达到最高。在盐胁迫下,GhHDZ46、GhHDZ66、GhHDZ119、GhHDZ146、GhHDZ169 在1 h 的表达量较高,在3 和6 h 降低,12 h 达到最高;GhHDZ12、GhHDZ92、GhHDZ177、GhHDZ193 在1 h 最低,12 h 达到最高。干旱胁迫下,GhHDZ12、GhHDZ66、GhHDZ119、GhHDZ169 在1 h 的表达量较高,在3 和6 h 表达量下降,24 h 表达量达到高峰;GhHDZ46、GhHDZ92、GhHDZ146、GhHDZ193 在1 h 的表达量最低,3、6 和12 h 逐渐升高,12 h 达到高峰(图6)。
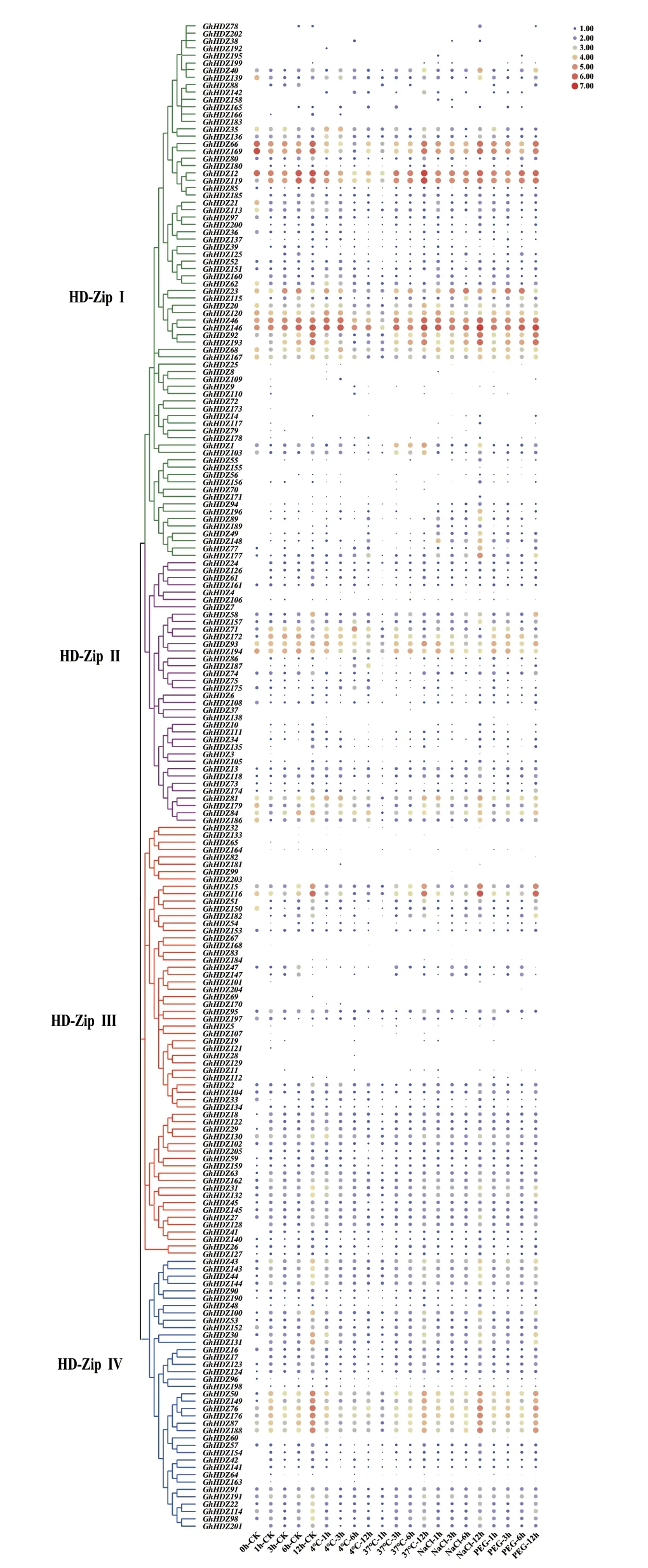
图6 GhHDZ 家族基因非生物胁迫下的表达模式Fig.6 Expression patterns of GhHDZ family genes under abiotic stress
对陆地棉HD-ZipII 亚组基因在4 种非生物胁迫处理的转录组分析,发现有26 个GhHDZ 基因的FPKM 值几乎为零,有6 个基因在各处理的表达量相对较高(图6)。其中,GhHDZ81、GhHDZ84 在冷、热、盐、干旱胁迫中表现为正向响应或者反向响应。
对陆地棉HD-ZipIII 亚组基因在4 种非生物胁迫处理的转录组分析,发现有38 个GhHDZ 基因未检测到转录本,其FPKM 值几乎为零(图6)。冷胁迫下,GhHDZ 基因表达量较低。热胁迫下,GhHDZ15、GhHDZ116 随着时间增长表达量越高,在12 h 达到高峰。GhHDZ15、GhHDZ116 在盐胁迫和干旱胁迫下的表现较为一致,在1 h 的表达量高于3 h,12 h 达到高峰。可见,GhHDZ15、GhHDZ116 在非生物胁迫中表达量较高。
对陆地棉HD-ZipIV 亚组基因在4 种非生物胁迫处理的转录组分析,发现有11 个GhHDZ 基因的FPKM 值几乎为零(图6)。在冷胁迫下,Gh-HDZ50、GhHDZ76、GhHDZ176 在1 h 的表达量 最高,在3 和6 h 逐渐下降,在12 h 升高。在热胁迫下,GhHDZ50、GhHDZ76、GhHDZ87、GhHDZ149、GhHDZ176、GhHDZ188 在1、3、6 和12 h 的表达量逐渐升高,12 h 达到峰值。在盐胁迫下,GhHDZ50、GhHDZ76、GhHDZ87、GhHDZ149、GhHDZ176、GhHDZ188 在1 h 表达量较高,在3 h 表达量下降,在6 和12 h 逐渐升高,12 h 达到高峰。在干旱胁迫 下,GhHDZ50、GhHDZ76、GhHD188 在1、3、6和12 h 的表达量逐渐升高,12 h 表达量最高。GhHDZ50/76/176/188 在冷、热、盐和干旱胁迫的处理中表达量较高。
2.8 10个GhHDZ基因在4种非生物胁迫下的RTqPCR分析
为了确定GhHDZ 在冷、热、盐和干旱胁迫下的表达水平,对陆地棉TM-1 幼苗进行非生物胁迫处理,并在0、3、6、12 和24 h 时间点取样。根据前述转录组热图分析结果,从中筛选了10 个在非生物胁迫中高表达的基因进行RT-qPCR 鉴定(图7)。
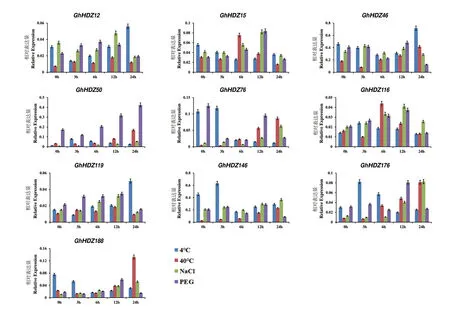
图7 10 个GhHDZs 在不同时间对非生物胁迫的相对表达量Fig.7 Relative expressions of 10 GhHDZs to abiotic stress at different time
在冷胁迫下,GhHDZ15、GhHDZ50、GhHDZ146、GhHDZ176 的RT-qPCR 结果和转 录组表达数据较一致。从RT-qPCR 结果可知,GhHDZ15、GhHDZ188 在0 h 的表达量最高,之后逐步下降,说明2 个基因负向响应冷胁迫;GhHDZ50、GhHDZ76、GhHDZ116、GhHDZ146、GhHDZ176 在3 h 的表达量最高,GhHDZ12、GhHDZ46、GhHDZ119 在24 h的表达量达到高峰,说明这些基因正向响应冷害刺激。上述结果表明,大部分筛选的基因对冷胁迫处理较敏感,反应迅速。
在热胁迫下,GhHDZ12、GhHDZ119 的RTqPCR 分析和转录组表达模式结果较一致(图7)。从RT-qPCR 结果可知,在0 和3 h 没有GhHDZ 基因达到高峰,GhHDZ15、GhHDZ116 在6 h 的表达量较高,GhHDZ12、GhHDZ119 在12 h 的表达量较高,GhHDZ46、GhHDZ50、GhHDZ76、GhHDZ146、GhHDZ176、GhHDZ188 在24 h 的表达量最高,说明这些基因正向响应热胁迫。
在盐胁迫下,GhHDZ12、GhHDZ46、GhHDZ116、GhHDZ119 的转录组表达模式和RT-qPCR分析结果较一致(图7)。从RT-qPCR 结果可知,在0 和6 h 没有GhHDZ 基因达到高 峰;GhHDZ46 基本不受盐胁迫影响;GhHDZ12、GhHDZ15、GhHDZ116、GhHDZ119 在12 h 的表达量达到顶峰,但其中GhHDZ12 表达量提高不明显,不到0 h 的1.5 倍;GhHDZ50、GhHDZ76、GhHDZ146、GhHDZ176、GhHDZ188 在24 h 的表达量最高,说明这些基因正向响应盐胁迫。
在干旱胁迫下,GhHDZ15、GhHDZ46、GhHDZ76、GhHDZ116、GhHDZ119、GhHDZ146、GhHDZ176、GhHDZ188 的转录组表达模式和RT-qPCR分析结果较一致(图7)。从RT-qPCR 结果可知,在3 h 没有GhHDZ 基因达到高峰;GhHDZ76 在0 h 的表达量较高,随着时间的推移,基本呈下降趋势,说明该基因负向响应干旱胁迫;GhHDZ12 在6 h 的表达量较高;GhHDZ15、GhHDZ46、GhHDZ116、GhHDZ119、GhHDZ146、GhHDZ176、GhHDZ188在12 h 的表达量达到顶峰,其中GhHDZ46、Gh-HDZ146 表达量增加不明显,不到0 h 的1.5 倍;GhHDZ50 在24 h 的表达量最高。上述结果说明,除GhHDZ46、GhHDZ76、GhHDZ146 外,其余基因正向响应干旱胁迫。
2.9 GhHDZ146的亚细胞定位分析
蛋白质的亚细胞定位与其功能密切相关。根据转录组和RT-qPCR 的结果表明,GhHDZ146 在低温、高温、盐胁迫和干旱胁迫处理下,在0、3、6、12 和24 h 的表达量均有明显不同,因此,选择GhHDZ146 进行亚细胞定位分析。结果(图8)表明,GhLTPG146-GFP 融合表达蛋白定位于细胞核中,表明GhLTPG146 可能作为转录因子在棉花的纤维发育中发挥作用。
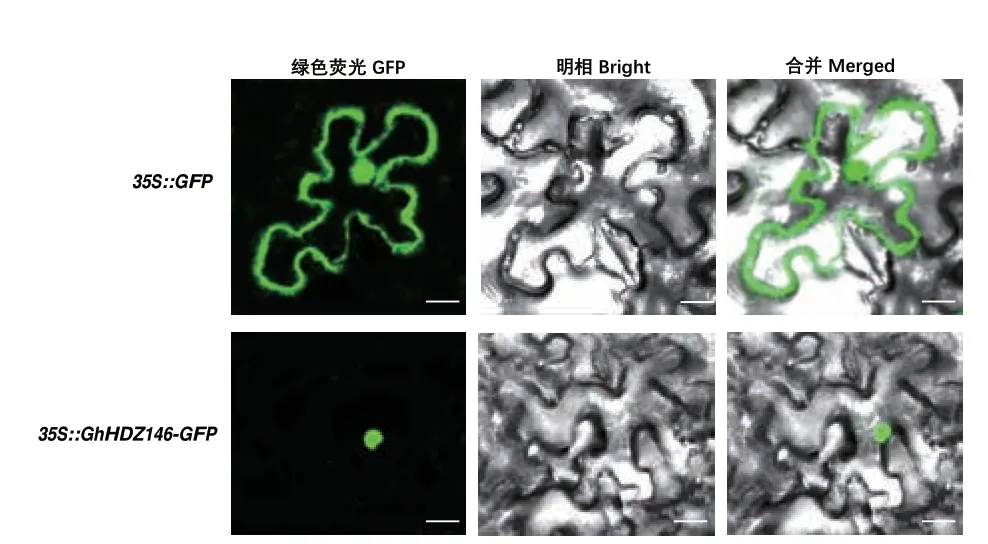
图8 GhHDZ146-GFP 融合蛋白在烟草叶片中的亚细胞定位(Bar=50 μm)Fig.8 Subcellular localizations of GhHDZ146 proteins in tobacco leaves (Bar=50 μm)
2.10 GhHDZ146的过表达转化拟南芥及盐胁迫试验
选择3 个GhHDZ146 过表达拟南芥T3代株系进行盐胁迫试验。结果(图9)表明,在200 mmol/L NaCl 胁迫处理下,OE5 和OE7 株系的主根长度分别为2.91 和2.82 cm,显著长于WT 的主根长度(2.36 cm);另外,OE2 株系的主根长度为2.98 cm,极显著长于WT 的根长。此外,进一步的根毛长度测量结果表明,OE2、OE5 和OE7 株系的根毛长度分别为91.3、94.5 和98.2 μm,均极显著长于WT 的根毛长度(68.3 μm)。上述结果表明,GhHDZ146 过表达拟南芥的耐盐性显著增强,认为GhHDZ146 对棉花的耐盐性有重要作用。
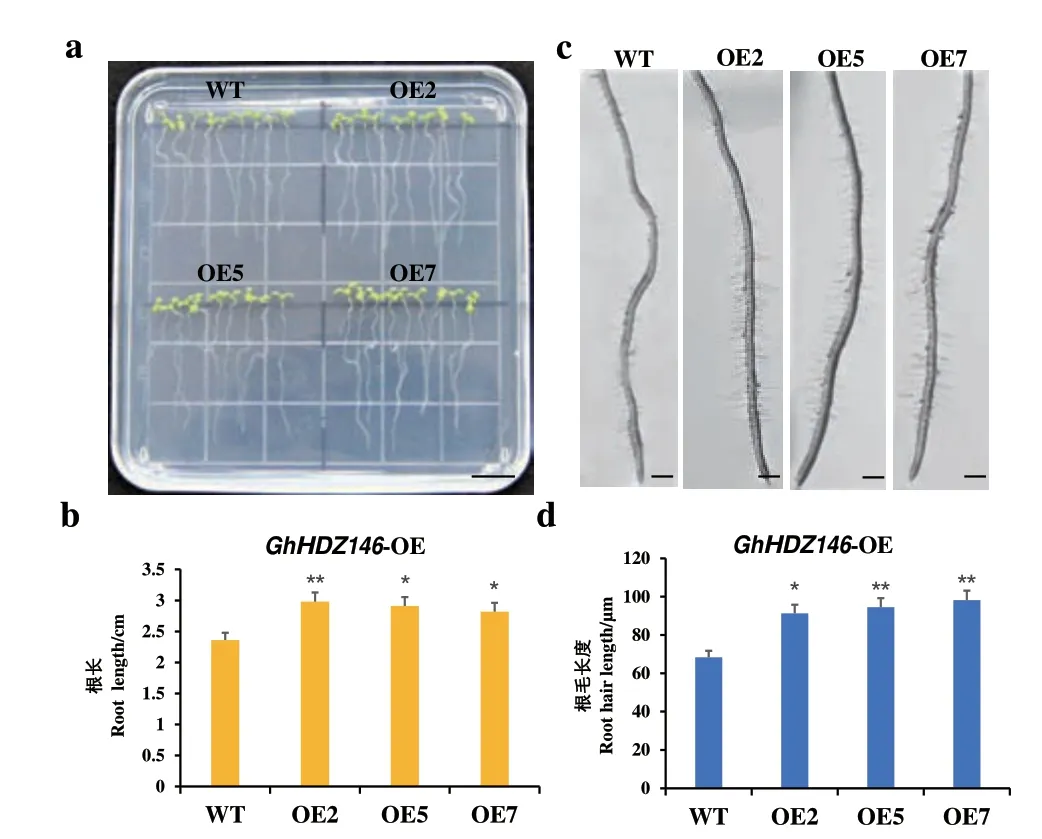
图9 GhHDZ146 过表达拟南芥的盐胁迫鉴定Fig.9 Salt stress identification of GhHDZ146 overexpression in Arabidopsis thaliana
进一步将GhHDZ146 与拟南芥、水稻等物种中已知功能的HD-Zip 家族基因进行了聚类分析,结果(图10)表明,GhHDZ146 与拟南芥AtHB1、AtHB6、AtHB13 的亲缘关系较近。

图10 GhHDZ146 与其他物种已知功能基因的聚类Fig.10 Clustering of GhHDZ146 and known functional genes in other species
3 讨论
HD-Zip 家族是植物特有的转录因子家族之一,许多植物中HD-Zip 基因家族的成员被依次鉴定,例如拟南芥、水稻[10,58]、葡萄[59]、大豆[60-61]、木薯[62]、小麦[12]、茶树[63]和马铃薯[14]等。在本研究中,从陆地棉TM-1 全基因组范围内中共鉴定出205 个成员。将陆地棉中HD-Zip 成员的数量与已报道的拟南芥[10](48 个)、水稻[58](49 个)、葡萄[59](33 个)、大豆[60](88 个)、木薯[62](57 个)、小麦(46 个)[12]、茶树[63](33 个)、马铃薯[14](43 个)以及芝麻[64](45 个)等植物相比,其家族成员数最多,而且不同物种之间其家族成员数量与其基因组大小并无一定的相关性。此外研究表明,基因家族的扩展主要依赖于串联、片段和全基因组等复制形式[65]。结合上述多个物种HD-Zip 基因家族成员分析,推测出现这种现象的原因可能是植物HD-Zip 家族基因在进化过程中不同物种间发生的片段复制和串联复制频率不同所致。
大量报道表明,植物HD-Zip 家族基因参与了对各种非生物胁迫的响应,如小麦[12]、马铃薯[14]、茶树[63]和芝麻[64]等物种中的HD-Zip 成员。然而,在陆地棉中还没有对冷、热、盐分或干旱做出反应的HD-Zip 基因的报道,本研究首次对陆地棉HDZip 家族基因在冷、热、盐或干旱胁迫条件下的表达模式进行了系统分析。转录组数据分析表明约45%GhHDZ 家族成员的表达水平受到冷、热、盐分或干旱胁迫的影响,对其中表达量较高的10 个基因进行了4 种非生物胁迫处理下的RT-qPCR 鉴定,结果也进一步证实部分陆地棉HD-Zip 家族基因参与了非生物胁迫响应,且各基因响应非生物胁迫的模式不同,本研究结果与其他植物中的HD-Zip 基因家族研究结果相似。
基因的进化树分析有利于给定植物物种中基因功能的预测。作为一个植物特异的转录因子家族,HD-Zip 基因在多种植物种类中已进行了广泛的研究。本研究中,GhHDZ146 与拟南芥AtHB1、AtHB6、AtHB13 的亲缘关系较近。据报道,在拟南芥当中AtHB1 在盐胁迫和低温胁迫下表达量下降[19],而本研究中转录组分析结合荧光定量验证均表明,GhHDZ146 正向响应热、盐和干旱胁迫,但在低温胁迫下与AtHB1 处理结果相似,整体上呈下降趋势,说明不同物种间同源基因的功能可能存在一些差异。另外,拟南芥AtHB6 在发育中的叶子、根和心皮中表达,但在遭受水分亏缺、渗透胁迫或用脱落酸(ABA)进行外源处理的幼苗中显著上调,表明该基因与逆境胁迫相关[66]。AtHB13 通过与JUNGBRUNNEN1(JUB1)启动子结合,参与抵抗干旱、高盐胁迫响应过程[67]。上述研究表明与拟南芥AtHB1、AtHB6、AtHB13 近缘的GhHDZ146 可能参与了非生物胁迫响应。本研究进一步通过分子遗传试验证实了GhHDZ146 在拟南芥中异源过表达可显著增强对盐胁迫的耐受性,且该基因定位于细胞核中,其可能作为转录因子发挥调控作用。然而,后续还需进一步通过棉花VIGS(病毒诱导的基因沉默)和棉花遗传转化等试验来进一步验证相关功能。本研究为陆地棉HD-Zip 基因参与非生物胁迫响应提供了可靠的证据,并为相关基因功能研究奠定了扎实的基础
4 结论
在全基因组范围内从陆地棉中系统鉴定了205个HD-Zip 家族成员。片段复制对GhHDZ 家族的扩展起主要作用,GhHDZ 家族基因具有潜在的生物学功能多样性。不同基因对各种胁迫的响应不一,且表达模式存在差异。GhHDZ146 定位于细胞核,异源过表达该基因可显著增强转基因拟南芥的耐盐性。
文章所有附表数据请到本刊官网下载(http://biotech.aiijournal.com/CN/1002-5464/home.shtml)。
