珍稀濒危沉水植物水菜花传粉生物学研究
2024-03-10王銮凤张同同向文倩梁惠婷谭珂申益春
王銮凤 张同同 向文倩 梁惠婷 谭珂 申益春


摘 要: 水菜花(Ottelia cordata)是珍稀瀕危的沉水植物,现仅分布在中国海南岛北部海口至文昌一带的淡水湿地,其造型优美,极具园艺观赏价值。然而,受人类活动影响,水菜花的生境急剧退缩且更加破碎化,濒临灭绝。为探究水菜花的繁殖特征和繁殖规律,该研究分析了水菜花的花部形态、性比、花粉活性、柱头可授性、反射光谱、花粉限制与无融合生殖及其传粉者类型与行为。结果表明:(1)水菜花雌雄异株,雌、雄个体的性比约为1∶2;水菜花雌、雄花形态相似,但雌花略大于雄花。(2)水菜花花粉活性和柱头可授性在花蕾期和开放期均较高。(3)中华蜜蜂(Apis cerana)是水菜花的主要传粉者,传粉效率较高且访问雄花的频率显著高于雌花。(4)水菜花雄蕊、雌蕊间的花色距离显著高于中华蜜蜂辨别的阈值(P<0.001)。(5)在自然授粉和人工授粉下,水菜花坐果率分别为95.29%和98.42%,并且不存在无融合生殖现象。综上所述,水菜花两性模拟的欺骗性传粉并不完美,传粉者可以从视觉上区分雌花和雄花;而由于水菜花雄花数量较多,传粉者低频率的“错访”足以达到水菜花雌花的繁殖需求。因此,在今后开展水菜花迁地保护、人工种群恢复等保育工作时,应重视水菜花种群大小、性别比例和当地传粉者种群数量。
关键词: 雌雄异株, 欺骗性传粉, 繁育系统, 花粉限制, 水鳖科
中图分类号: Q945.5
文献标识码: A
文章编号: 1000-3142(2024)01-0157-10
Pollination biology of the rare and endangered
submerged plant, Ottelia cordata
WANG Luanfeng1,2, ZHANG Tongtong2, XIANG Wenqian2,
LIANG Huiting1, TAN Ke3*, SHEN Yichun1,2*
( 1. Key Laboratory of Genetics and Germplasm Innovation of Tropical Special Forest Trees and Ornamental Plants, Ministry of Education, Hainan
University, Haikou 570228, China; 2. Center for Terrestrial Biodiversity of the South China Sea, Hainan University, Haikou 570228, China;
3. Guangxi Key Laboratory of Plant Conservation and Restoration Ecology in Karst Terrain, Guangxi Institute of Botany, Guangxi
Zhuang Autonomous Region and Chinese Academy of Sciences, Guilin 541006, Guangxi, China )
Abstract: Ottelia cordata is a kind of rare and endangered submerged plant that grows in freshwater wetlands in northern Hainan Island, from Haikou to Wenchang in China. This species is known for its beautiful appearance and the distinct “flower river” landscape, which makes it a high ornamental value. With the intensification of human activities, the habitat of O. cordata has been sharply reduced and fragmented, leading to its endangerment. In order to explore the reproductive characteristics and reproductive laws of O. cordata, the flower morphology, sex ratio, pollen viability, stigma receptivity, reflectance spectrum, pollen limitation and apomixis, and the types and behaviors of pollinators in O. cordata were analyzed. That is, the pollination biology of O. cordata were studied for its further in-situ and ex-situ conservation. The results were as follows: (1) The flowers were dioecism, and the sex ratio of female and male individuals was about 1∶2; the morphologies of male and female flowers were similar, and the female flowers were slightly larger than the male. (2) O. cordata had high pollen viability and stigma receptivity in budding and blooming phases. (3) Apis cerana was the main pollinator of Ottelia cordata, which visited male flowers more frequently than female flowers, with higher pollination efficiency. (4) The distance between the color loci of pistils and stamens was significantly higher than the discrimination threshold of Apis cerana. (5) The fruit setting percentage under natural pollination was similar to that under artificial pollination (95.29% and 98.42%, respectively), and there was no apomixis and low pollen limitation in the natural population of Ottelia cordata. In conclusion, the deceptive pollination of O. cordata simulated by both sexes is imperfect. Pollinators can visually distinguish female and male flowers. However, the large number of male flowers of O. cordata attracted many insects to visit flowers, which increases the probability of “mistake” pollination and is enough for the reproductive needs of female flowers. Therefore, future ex-situ conservation and artificial population restoration should pay more attention to population size, sex ratio and the number of local pollinators.
Key words: dioecism, deceptive pollination, breeding system, pollen limitation, Hydrocharitaceae
湿地生态系统作为全球三大生态系统之一,不仅在涵养水源、抵御洪水、补给地下水等方面具有独特的水调节功能,同时在生物多样性维持、调节局域气候、固碳、净化水质等方面还具有诸多生态功能,被誉为“地球之肾”。但是,人类世(Anthropocene)以来的快速经济发展和城市化进程,以及随之带来的全球气候变暖、水体富营养化、外来物种入侵等一系列环境问题导致湿地急速退化,湿地水生植被种类减少,大量珍稀濒危物种面临着灭绝的风险(李伟,2020;Fluet-Chouinard et al., 2023;祝惠等,2023)。海南岛是我国唯一的热带大陆型岛屿,设立了海南热带雨林国家公园,然而海南湿地尤其是淡水湿地并未得到同等重视。养殖场随意排污、凤眼莲(Eichhornia crassipes)和大薸(Pistia stratiotes)快速入侵等事件依然时常发生,严重威胁着该地区水菜花(Ottelia cordata)、邢氏水蕨(Ceratopteris shingii)、野生稻(Oryza rufipogon )等诸多珍稀濒危水生植物的生长。其中,国家二级保护植物水菜花的个体数量仅4年时间(2017—2020)就下降了11%(Shen et al., 2021),生存状况极不乐观。
水菜花隶属于水鳖科(Hydrocharitaceae)海菜花属(Ottelia),雌雄异株,一年生或多年生沉水植物(Wang et al., 2010)。水菜花一年四季均可开花,其单花花期通常约为1 d。花单性,雌雄异株;花瓣3 枚;雄佛焰苞内有雄花10~30 朵,而雌花内仅1 花;雄蕊12 枚,花柱9~18(Wang et al., 2010)。水菜花花多而大、花期较长、辐射对称、花粉量大,符合典型的泛化传粉系统(张大勇,2004)。水菜花对水体变化较为敏感,对生境要求极为苛刻,喜生长在清澈洁净的流水沟渠及池塘中(于丹等,1998; Chen et al., 2012)。随着人类对淡水湿地的开发、外来生物的入侵、水资源的污染,适宜水菜花生长的环境越来越少。目前,仅零星分布于缅甸、泰国、柬埔寨和中国南方地区,在我国广西种群灭绝后,现仅分布在海南北部火山岩淡水湿地(何景彪等,1991; 于丹等,1998; Chen et al., 2012; 韦毅刚,2019; Shen et al., 2021),可作为海南淡水湿地的“伞护种”开展保育工作。
水菜花花瓣洁白、一抹淡黄点缀在花中央,一簇簇、一丛丛随波荡漾在清澈的溪面,在海南淡水湿地呈现出独特的“花河”景观。水菜花的浮水叶宽披针形或长卵形露出水面,衬托着“花河”;其沉水叶长椭圆形、披针形或带形,全缘,在水下随波逐流,具极高的园艺观赏价值(赵佐成等,1984)。东南亚部分国家还将水菜花当作蔬菜,制作沙拉或者蔬菜汤,并且水菜花具有一定的药用价值(Zhang et al., 2020)。由此可见,水菜花还是海南淡水湿地的“旗舰种”。尽管我国生物多样性保护意识逐渐提升,海南对淡水湿地的保护也稳步加强,但水菜花的适生区仍在逐渐减小(Shen et al., 2021)。因此,开展水菜花的繁殖和保育生物学研究极为迫切。本研究以海口昌旺溪湿地保护小区水菜花自然种群为研究对象,通过比较雌、雄花的花部形态、性比、花粉活性、柱头可授性、反射光谱以及检测花粉限制与无融合生殖,拟探讨:(1)水菜花自然种群的繁殖特征;(2)水菜花是否存在显著的花粉限制导致物种濒危;(3)水菜花的繁殖规律。以期为后续的就地或迁地保护提供理论依据。
1 材料与方法
1.1 试验材料和研究地点
2021 年12月至2022年4月,在海南省海口昌旺溪湿地保护小区(110°17′ E、19°47′ N)进行水菜花野外试验。该区域内水菜花自然种群长势良好,其生境受人为干扰较弱,便于开展水菜花自然种群的传粉生物学研究(图1)。
1.2 性比和花部特征
在研究样地中,设置10 个2 m × 2 m的样方,用于统计水菜花雌、雄株性比。随机选取待开放的雌、雄花苞各10 朵,每天9:00—21:00观察并统计单花状态及开花进程。随后,随机选取完全开放的雌、雄花各20 朵,用游标卡尺测定花瓣(长和宽)长度、性器官长度、花梗长度、蜜导半径、花萼直径等参数。
1.3 花粉活性和柱头可授性的检测
采用蓝墨水染色法测定花粉活性(王伟伟等,2018)。将5 枚不同时期(花蕾期、开放期Ⅰ、开放期Ⅱ、开放期Ⅲ和凋谢期)的花药放置在2 mL的离心管中,设置4 个重复,之后滴入1 μL浓度1%的蓝墨水溶液,室温下静置10 min后用光学显微镜观察。随机选取5 个分离的视野,每个视野不少于30 粒花粉,花粉被染成蓝色表示花粉无活性(王伟伟等, 2018)。
采用联苯胺-过氧化氢法对柱头可授性进行测定(Dafni & Maués, 1998)。详细记录柱头上气泡开始冒泡的时间和柱头周围的颜色变化,并根据柱头先端产生气泡数和染色深淺程度判定柱头的活性大小。其中,蓝色且产生大量气泡的柱头为有活性柱头,气泡较少或无气泡的柱头则为无活性柱头(Dafni & Maués, 1998)。
1.4 访花者行为观察
参照龚燕兵和黄双全(2007)的方法,选择晴朗天气观测水菜花的访花者。记录不同访花者的访花时间、访花频率及种类等,并判断水菜花的主要传粉者。此外,为了更好地记录访花昆虫的访花过程和觅食行为,我们还利用4 台全景4 K高清户外摄影机(萤虫S6运动相机)架设在样地进行拍摄记录。
1.5 反射光谱分析
选取雌、雄花各5 朵的花瓣、蜜导、花药、柱头和叶片,使用便携式光谱仪(赛曼S3000-UV-NIR)测定光谱反射率。同时将测得的光谱数据标记到膜翅目蜂类昆虫视觉颜色的六边形模型中(Chittka et al., 1992; Chittka & Kevan, 2005)。使用多元方差分析(MANOVA)检测水菜花雌、雄花之间坐标是否存在显著性差异,以坐标(x和y)为变量,用Pillais trace进行F检验。差异不显著则表明昆虫无法区分花色;反之,再使用t检验,比较两两花色间的颜色距离是否在蜜蜂的颜色辨别阈值内(Giurfa, 2004)。
1.6 花粉限制和无融合生殖的检测
分别统计雌、雄花各10 朵的胚珠和花粉数量。在统计单花的花粉数量时,用解剖针小心将雄花花药内壁上的所有花粉颗粒充分剥离,并移入4 mL的50%乙酸溶液中,在光学显微镜下记录来自4 μL混合液体的花粉颗粒,重复10 次。将每朵雌花的胚珠数量在光学显微镜下观察并进行计数统计。
为了检测水菜花是否存在花粉限制和无融合生殖现象,我们对比了人工授粉和自然授粉的坐果率和结实率。本研究共设置3 个试验处理,每个处理重复10次。(1)直接套袋:对雌花不做任何处理,在花未开放前直接套袋;(2)人工授粉:在柱头成熟时涂抹适量的花粉后并套袋;(3)自然授粉。待种子成熟后,采用自然授粉和人工授粉的结实率来计算花粉限制的强度(pollen limitation,PL),PL=(1-C/X) × 100,式中C和X分别代表在自然和人工授粉下每个果实的种子产量(Yang et al., 2020)。当PL等于0就意味着其不受到花粉限制,PL越趋近于0表示其受到的花粉限制越小;相反,当PL越趋近于1则表示其受到的花粉限制越大(Larson & Barrett, 2000)。
1.7 数据统计分析
利用SPSS 26.0 for Windows软件中的MANOVA以及one-way ANOVA进行统计分析,统计数据用x[TX-1mm]±s表示。
2 结果与分析
2.1 性比和花部特征
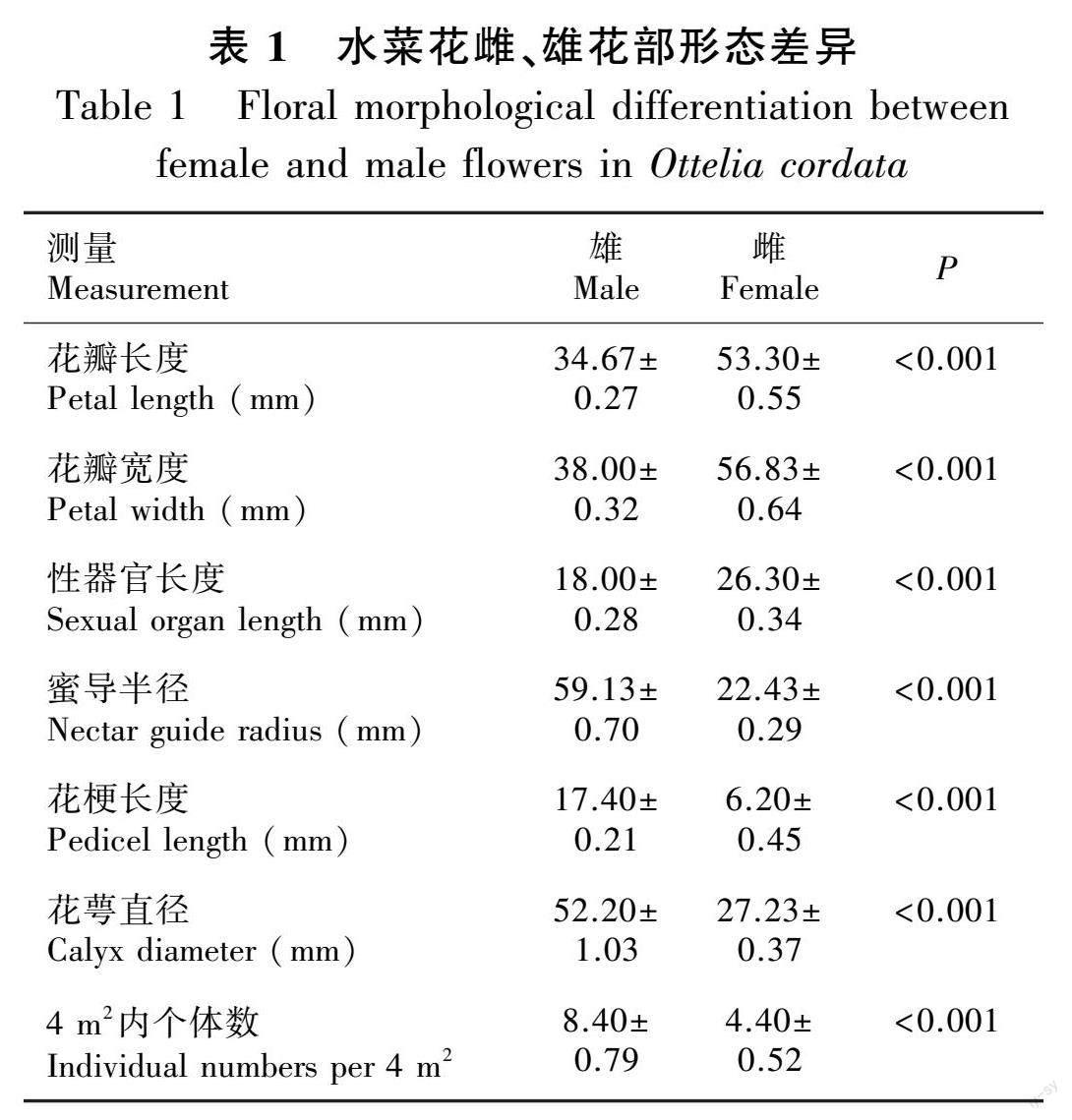
由表1可知,昌望溪水菜花自然种群内,雌、雄个体的性比约为1∶2,雄性个体数量显著高于雌性。水菜花雌、雄花在颜色和形状上极为相似,但雌花的花瓣、性器官、蜜导、花萼和花梗都明显大于雄花。
由图2可知,水菜花单花开放过程可分为花蕾期、开放期Ⅰ、开放期Ⅱ、开放期Ⅲ和凋谢期5个过程。(1)花蕾期,雌、雄花瓣由萼片包裹,雌、雄蕊缓慢伸长,均由浅绿色变为黄色,花药在花冠裂片内便已开裂散粉;(2)开放期Ⅰ,雌、雄花蕾逐渐变大,花瓣主要以左旋、右旋或同时开裂,此时,花丝和柱头颜色由黄色变深黄色,该阶段花药散粉达到高峰,花瓣内侧沾满花粉;(3)开放期Ⅱ,雌、雄花瓣逐渐舒展开,花药散粉变少;(4)开放期Ⅲ,雌、雄花瓣张开,花药散粉结束,花丝颜色由深黄色变橘黄色;(5)凋谢期,雌花的花瓣凋谢,柱头授粉成功后呈褐色,雄花花药逐渐由橘黄色变锈黄色,花瓣衰败。大约在1 个月后,蒴果便会自然开裂。
2.2 花粉活性和柱头可授性的检测
由图3可知,水菜花花粉活性和柱头可授性变化趋势基本相同。花蕾期(花冠裂片刚要打开)花粉已具有较高的活性(93.4%),柱头可授性也较高(80.2%);花蕾期和开放期的花粉活性和柱头可授性均保持在80%以上。花瓣完全盛开后,花粉活性和柱头可授性开始下降;凋谢期花粉活性已降至7%,柱头可授性仅为21%。
2.3 访花者行为观察
本研究观测到膜翅目(Hymenoptera)和双翅目(Diptera)的5 种昆虫访问水菜花(图4),昆虫访花高峰期集中在上午10:00—11:00(图5)。其中,膜翅目昆虫为中华蜜蜂(Apis cerana)和淡脉隧蜂属昆虫(Lasioglossum sp.);双翅目昆虫为黑带食蚜蝇属昆虫(Episyrphus sp.)、蝇科昆虫(Muscidae sp.)和丽蝇科昆虫(Calliphoridae sp.)(图4)。携粉量最大的是中华蜜蜂和淡脉隧蜂属昆虫(图4),并且它们访问雄花的频率和时间均显著高于雌花(P<0.001)(图5,图6),而其他昆虫几乎不或者很少携带花粉。
中華蜜蜂访问雄花的频率最高,约为访问雌花频率的3 倍,但其访花时间仅为5~17 s(图5,图6)。淡脉隧蜂属昆虫访花时间较长,可达1~2 min,但其访花频率不及中华蜜蜂的1/3(图5,图6)。当这两种膜翅目昆虫访花时,常以蜜导部位或花药为降落点,沿着蜜导方向爬行,其头部、胸部和腹部会沾满花粉(图4)。因此,我们认为中华蜜蜂是水菜花的主要传粉者且传粉效率较高,淡脉隧蜂属昆虫为潜在传粉者。
2.4 反射光谱分析
水菜花反射光谱分析结果显示,水菜花雌、雄花瓣颜色位点(Pillais轨迹=0.564,F=4.518,P=0.055)和蜜导颜色位点(Pillais轨迹=0.366,F=2.017,P=0.203)无明显颜色差异(图7,图8)。然而,雌蕊和雄蕊存在显著的颜色差异(0.54 vs 0.06,t=8.123,df=9,P < 0.001),而且水菜花雌蕊、雄蕊之间的花色距离明显高于蜜蜂的辨别阈值,即水菜花的主要传粉者中华蜜蜂可以区分水菜花的雌蕊和雄蕊。
2.5 花粉限制和无融合生殖的检测
水菜花的花粉和胚珠数量分别为(119 223 ± 12 085.282)和(4 256±133.079)。水菜花坐果率和结实率在自然授粉下分别为95.29%和100%,在人工授粉下则为98.42%和100%;花粉限制(PL)值为0.15,说明水菜花野外种群存在较低花粉限制。直接套袋处理下,水菜花的坐果率和结实率均为0,表明其繁育系统不存在无融合生殖现象。
3 讨论与结论
3.1 水菜花的繁育特征
水菜花虽属于沉水植物,但传粉过程须在水上完成,水下完成果实生长发育。本研究中水菜花的主要传粉者是中华蜜蜂,这与其同属植物海菜花(Ottilia acuminata)的结果一致(Yang et al., 2020)。不同的是,淡脉隧蜂属昆虫访问水菜花的访花时间较长,但全身携带花粉,可能是水菜花潜在的主要传粉者,而海菜花的则为蝇类(Yang et al., 2020),这可能与当地的传粉者环境(pollinator environment)相關。
花粉是水菜花为访花者提供的主要回报物。本研究中,雄花的花粉量大,中华蜜蜂访问雄花的频率高达每小时110 次。中华蜜蜂降落在雄花上后,就以头部钻入花冠筒中采集花粉,访花时间为8~17 s,全身就能沾满花粉。而雌花无花粉,中华蜜蜂访问雌花的频率相对较低,仅为访问雄花频率的1/3,并且在雌花成熟柱头上停留时间较短,仅5 s左右便飞离。尽管如此,水菜花自然授粉的坐果率高达95%以上,说明中华蜜蜂传粉效率高,较低的访花频率和时间就已经能满足水菜花雌花的繁殖需求。
自然界85%的单性花都有花大小二型性(Delph et al., 1996),水菜花也具有花大小二型性现象,即同一植物的雌雄花在大小上有明显的差异。不同的是,绝大多数温带动物传粉的雄花往往都有更大的花和更显著的花展示(Bell, 1985; Delph et al., 1996; Eckhart, 1999; Costich & Meagher, 2001),而生活在热带地区的水菜花恰恰相反,水菜花雌花的花瓣、性器官、蜜导、花萼和花梗都明显大于雄花,这种现象在热带生态系统中也比较多见(Delph et al., 1996; Humeau et al., 2003)。这可能也与花数量有关,花展示的理论模型认为花大小和花数量之间存在负相关(Harder & Barrett, 1996),水菜花雄花的数量远大于雌花,雄株一个佛焰苞内便有雄花10~30 朵,而雌株佛焰苞内仅1 朵雌花;并且雄株的数量也是雌性个体的2 倍,进一步增加了雄花的数量。同时,花形成花序后对传粉者的吸引显然要大于单花,甚至也大于单花效应的总和(Mulligan & Kevan, 1973)。再加上雌花没有传粉报酬,只能通过更大的花展示等方式来吸引传粉者,以此在广告效应上赶上或者超过雄花(Renner & Feil, 1993)。
3.2 水菜花两性模拟的欺骗性传粉
水菜花的雌、雄花在颜色和形状上极为相似,二者花瓣白色,花中央均有黄色蜜导,相同的结果也体现在蜂类的视觉上,仅雌蕊(花柱9~18 枚)和雄蕊(12 枚)的反射光谱存在一定的差异,这可能与雄蕊上存在大量花粉有关,花粉粒的存在导致光谱纯度更高,最终使传粉者可以区分雄蕊和雌蕊(Yang et al., 2020)。由此可见,没有传粉报酬的雌花拟态有报酬(花粉)的雄花进行两性模拟(intersexual mimicry)的欺骗性传粉,这与其同属植物海菜花完全相同(Yang et al., 2020),并且在欺骗传粉中较为常见(Baker, 1976; Dafni, 1984)。
雄花不仅具有传粉者报酬,并且花朵数量也远多于雌花,为此,传粉者势必会对雌花产生歧视,从而更倾向于访问报酬更多的雄花( gren et al., 1986; Dukas, 1987; Vaughton & Ramsey, 1998)。本研究的反射光谱分析结果显示,雌蕊和雄蕊存在明显的颜色差异,说明蜜蜂也能够从视觉上对两者进行区分。实际上,我们在野外观察到的结果也正是如此,相比雌花,中华蜜蜂对雄花有明显的访花偏好,存在对雌花的访花歧视,可能导致花粉限制(pollen limitation)。然而,本研究结果显示,水菜花中并未出现明显的花粉限制(PL=0.15),甚至还要低于224 种动物媒介开花植物的平均花粉限制(PL=0.4)(Larson & Barrett, 2000),以及其同属植物海菜花(PL=0.34)(Yang et al., 2020)。综上所述,我们认为,尽管中华蜜蜂对雌花存在一定的访花歧视,但传粉结果非常成功,水菜花极高的自然结实率和坐果率也证实了该结果。
由于传粉昆虫的学习行为,欺骗性空花的访问经历会使昆虫避开那些没有报酬的刺激,下一次避免造访同一类型的花(Simonds & Plowright, 2004; 任宗昕等,2012)。但水菜花在昌旺溪形成壮观的“花河”景观,庞大的雄花数量吸引了大量访花者,相比之下,出现频率较低的无奖励雌花对传粉者本身学习行为惩罚较小,甚至可以忽略(Schaefer & Ruxton, 2009; Yang et al., 2020)。由此可见,雌花受到了负频率依赖选择(negative frequency-dependent selection)作用,当样地内的水菜花雄花频率升高后,雌花也获得了更多的传粉者造访。这种选择作用在兰科植物中比较常见,如人为增加Dactylorhiza sambucina居群内黄色表型比例后,稀有的紫花表型繁殖成功率也随之上升,反之亦然,以此保证稀有表型在种群内的稳定(Gigord et al., 2001),这可能也是自然种群中水菜花雄性比例偏高的关键原因之一。
性比和传粉者数量可能是导致水菜花传粉成功的关键原因(ster & Eriksson, 2007; Xia et al., 2013; Yang et al., 2020)。由于传粉者对花辨别的准确性并不是一个严格的要求,并且不同的传粉者个体之间也会存在差异(Chittka et al., 2003)。随着传粉者访问雄花频率的增加,传粉者对雌花“错访”频率也随之增加。虽然“模仿”或“欺骗”可以被传粉者识别,但仍有部分传粉者“意外”访问雌花,并导致“错访”低频率的发生(Krishna & Somanathan, 2018; Yang et al., 2020);兼之,中华蜜蜂传粉效率较高,低频率的“错访”就已经能满足雌花的繁殖需求。由此推断,当大量传粉者访问水菜花种群时,它们对雌花的访问也会增加,可以抵消传粉歧视可能引发的花粉限制。在这种情况下,不完全的两性模拟欺骗传粉是有效的。
综上所述,维持水菜花这一濒危植物的繁殖与更新,需要保证雌花传粉者的传粉效率和较高访花频率(Xia et al., 2013),而这主要取决于种群的大小、性别比例和传粉者的类型与数量。当种群数量急剧萎缩时,水菜花可能会出现较高的传粉限制,传粉系统将更加脆弱,难以维持。因此,开展水菜花保育和人工种群恢复等工作时,应重点关注种群的规模,雌雄个体比例,以及当地的传粉者环境。
致谢 感谢海南湿地保护协会周缘老师、海口畓榃湿地研究所卢刚老师以及海南大学许宇辉、王若宾同学对本论文野外工作给予的幫助,感谢海南大学任明迅教授和张哲博士对本论文的指导,以及各位审稿人提供的宝贵意见!
参考文献:
G REN J, ELMQVIST T, TUNLID A, 1986. Pollination by deceit, floral sex ratios and seed set in dioecious Rubus chamaemorus L. [J]. Oecologia, 70(3): 332-338.
BAKER HG, 1976. ‘Mistake pollination as a reproductive system with special reference to the Caricaceae [C]. London: Linnean Society Symposium: 161-169.
BELL G, 1985. On the function of flowers [J]. Proc Royal Soc B Biol Sci, 224(1235): 223-266.
CHEN LY, CHEN JM, GITURU RW, et al., 2012. Generic phylogeny, historical biogeography and character evolution of the cosmopolitan aquatic plant family Hydrocharitaceae [J]. BMC Evol Biol, 12(1): 30.
CHITTKA L, BEIE W, HERTEL H, et al., 1992. Opponent colour coding is a universal strategy to evaluate the photoreceptor inputs in Hymenoptera [J]. J Comp Physiol, 170(5): 545-563.
CHITTKA L, DYER AG, BOCK F, et al., 2003. Bees trade off foraging speed for accuracy [J]. Nature, 424(6947): 388.
CHITTKA L, KEVAN PG, 2005. Flower colors as advertisement [M]// DAFNI A, KEVAN PG, HUSBAND B. Enviroquest. Canada: Practical Pollination Biology: 157-206.
COSTICH DE, MEAGHER TR, 2001. Impacts of floral gender and whole-plant gender on floral evolution in Ecballium elaterium (Cucurbitaceae) [J]. Biol J Linn Soc, 74(4): 475-487.
DAFNI A, 1984. Mimicry and deception in pollination [J]. Ann Rev Ecol Syst, 15: 259-278.
DAFNI A, MAUES MM, 1998. A rapid and simple procedure to determine stigma receptivity [J]. Sex Plant Reprod, 11(3): 177-180.
DELPH LF, GALLOWAY LF, STANTON ML, 1996. Sexual dimorphism in flower size [J]. Am Nat, 148(2): 299-320.
DUKAS R, 1987. Foraging behavior of three bee species in a natural mimicry system: female flowers which mimic male flowers in Ecballium elaterium (Cucurbitaceae) [J]. Oecologia, 74(2): 256-263.
ECKHART VM, 1999. Sexual dimorphism in flowers and inflorescences [M]// GEBER MA, DAWSON TE, DELPH LF. Gender and sexual dimorphism in flowering plants. Berlin: Springer: 123-148.
FLUET-CHOUINARD E, STOCKER BD, ZHANG Z, et al., 2023. Extensive global wetland loss over the past three centuries [J]. Nature, 614(9): 281-286.
GIGORD LDB, MACNAIR MR, SMITHSON A, 2001. Negative frequency-dependent selection maintains a dramatic flower color polymorphism in the rewardless orchid Dactylorhiza sambucina (L.) Soò [J]. Proc Natl Acad Sci USA, 98(11): 6253-6255.
GIURFA M, 2004. Conditioning procedure and color discrimination in the honeybee Apis mellifera [J]. Naturwissenschaften, 91(5): 228-231.
GONG YB, HUANG SQ, 2007. On methodology of foraging behavior of pollinating insects [J]. Biodivers Sci, 15(6): 576-583. [龚燕兵, 黄双全, 2007. 传粉昆虫行为的研究方法探讨 [J]. 生物多样性, 15(6): 576-583.]
HARDER LD, BARRETT SCH, 1996. Pollen dispersal and mating patterns in animal-pollinated plants [M]//LLOYD DG. Floral biology. Boston: Springer: 140-190.
HE JB, SUN XZ, ZHONG Y, et al., 1991. Study on the branches of the genus Camellia [J]. J Wuhan Bot Res, 9(2): 121-129. [何景彪, 孙祥钟, 钟扬, 等, 1991. 海菜花属的分支学研究 [J]. 武汉植物学研究, 9(2): 121-129.]
HUMEAU L, PAILLER T, THOMPSON JD, 2003. Flower size dimorphism in diclinous plants native to La Réunion Island [J]. Plant Syst Evol, 240(1/4): 163-173.
KRISHNA S, SOMANATHAN H, 2018. Intersexual mimicry and flowering phenology facilitate pollination in a dioecious habitat specialist species,Myristica fatua (Myristicaceae) [J]. Plant Ecol, 219(10): 1247-1257.
LARSON BMH, BARRETT SCH, 2000. A comparative analysis of pollen limitation in flowering plants [J]. Biol J Linn Soc, 69(4): 503-520.
LI W, 2020. Research and practice of aquatic plant diversity conservation in China [J]. Yangtze River, 51(1): 104-112. [李偉, 2020. 我国水生植物多样性保护的研究与实践 [J]. 人民长江, 51(1): 104-112.]
MULLIGAN GA, KEVAN PG, 1973. Color, brightness, and other floral characteristics attracting insects to the blossoms of some Canadian weeds [J]. Can J Bot, 51(10): 1939-1952.
STER M, ERIKSSON O, 2007. Sex ratio mediated pollen limitation in the dioecious herb Antennaria dioica [J]. Ecoscience, 14(3): 387-398.
REN ZX, WANG H, LUO YB, 2012. Deceptive pollination of orchids [J]. Biodivers Sci, 20(3): 270-279. [任宗昕, 王红, 罗毅波, 2012. 兰科植物欺骗性传粉 [J]. 生物多样性, 20(3): 270-279.]
RENNER SS, FEIL JP, 1993. Pollinators of tropical dioecious angiosperms [J]. Am J Bot, 80(9): 1100-1107.
SCHAEFER HM, RUXTON GD, 2009. Deception in plants: mimicry or perceptual exploitation? [J]. Trend Ecol Evol, 24(12): 676-685.
SHEN YC, LEI J, SONG XQ, et al., 2021. Annual population dynamics and their influencing factors for an endangered submerged macrophyte (Ottelia cordata) [J]. Front Ecol Evol, 9: 688304.
SIMONDS V, PLOWRIGHT CMS, 2004. How do bumblebees first find flowers? Unlearned approach responses and habituation [J]. Anim Behav, 67(3): 379-386.
VAUGHTON G, RAMSEY M, 1998. Floral display, pollinator visitation and reproductive success in the dioecious perennial herb Wurmbea dioica (Liliaceae) [J]. Oecologia, 115(1/2): 93-101.
WANG QF, GUO YH, ROBERT RH, et al., 2010. Hydrocharitaceae [M]. WU ZY, RAVEN PH, HONG DY. Flora of China. Beijing: Science Press; Missouri Botanical Garden Press: 23.
WANG WW, YAO L, HE KY, et al., 2018. Comparative study on in vitro pollen germination and viability evaluation of different willow species [J]. J Jiangsu For Sci Technol, 45(4): 1-7. [王伟伟, 姚玲, 何开跃, 等, 2018. 不同柳树花粉离体培养及活力测定方法比较研究 [J]. 江苏林业科技, 45(4): 1-7.]
WEI YG, 2019. The distribution and conservation status of native plants in Guangxi [M]. Beijing: Forestry Publishing House: 601. [韦毅刚, 2019. 广西本土植物及其濒危状况 [M]. 北京: 中国林业出版社: 601.]
XIA J, LU J, WANG ZX, et al., 2013. Pollen limitation and Allee effect related to population size and sex ratio in the endangered Ottelia acuminata (Hydrocharitaceae): implications for conservation and reintroduction [J]. Plant Biol, 15(2): 376-383.
YANG J, NIU Y, SUN WB, et al., 2020. Intersexual mimicry and imperfect deceit of a threatened aquatic herb Ottelia acuminata [J]. J Syst Evol, 60(2): 377-385.
YU D, CHONG YX, TU MH, et al., 1998. Study on the threatened aquatic higher plant species of China [J]. Biodivers Sci, 6(1): 13-21. [于丹, 種云霄, 涂芒辉, 等, 1998. 中国水生高等植物受危种的研究 [J]. 生物多样性, 6(1): 13-21.]
ZHANG DY, 2004. Plant life-history evolution and reproductive ecology [M]. Beijing: Science Press: 155-361. [张大勇, 2004. 植物生活史进化与繁殖生态学 [M]. 北京: 科学出版社: 155-361.]
ZHANG Y, LI JW, SAN MM, et al., 2020. The secret of health in daily cuisine: typical healthy vegetables in local markets in central Myanmar [J]. J Ethnobiol Ethnomed, 16(1): 73.
ZHAO ZC, SUN XZ, WANG HQ, et al., 1984. A study on the ecology of the fresh water Hydrocharidaceae in South China [J]. Acta Ecol Sin, 4(4): 354-363. [赵佐成, 孙祥钟, 王徽勤, 1984. 华南地区淡水水鳌科植物的生态特征和群落学观察 [J]. 生态学报, 4(4): 354-363.]
ZHU H, WU HT, XING XX, et al., 2023. Achievement of wetland protection and restoration and development strategies in China [J]. Bull Chin Acad Sci, 38(3): 365-375. [祝惠, 武海涛, 邢晓旭, 等, 2023. 中国湿地保护修复成效及发展策略 [J]. 中国科学院院刊, 38(3): 365-375.]
( 责任编辑 邓斯丽 )
收稿日期: 2023-04-05
基金项目: 海南省自然科学基金青年基金(321QN185); 海南省自然科学基金高层次人才项目(422RC594); 广西科学院基本业务费项目(CQZ-C-1901); 广西喀斯特植物保育与恢复生态学重点实验室基金(22-035-26)。
第一作者: 王銮凤(1997-),硕士研究生,研究方向为风景园林植物与应用,(E-mail)752074511@qq.com。
* 通信作者: 谭珂,博士,研究方向为植物适应与演化、生物地理学和保护生物学,(E-mail)tanke@gxib.cn; 申益春,博士,研究方向为湿地植物景观与多样性保育,(E-mail)1279412884@qq.com。
