虎耳草属的资源分类及育种进展
2024-03-10唐世梅张霞陈倩如蔡文淇李炎林田代科
唐世梅 张霞 陈倩如 蔡文淇 李炎林 田代科

摘 要: 虎耳草屬(Saxifraga Tourn. ex L.)资源丰富且分布广泛,全球有440~500个种,其分类一直受到植物学家的关注。该属具有重要的观赏和药用价值,欧洲多国非常重视其观赏资源的开发利用。虎耳草属的品种培育距今已有150多年的历史,到2022年国际虎耳草协会(The Saxifrage Society)网站收录了1 692个品种,但只有1个品种来自中国。可见,我国虽然是虎耳草属的多样性中心之一,但对其观赏资源的开发利用远远落后于欧美甚至日本。该文从虎耳草属的种质资源、分类概况、育种进展等方面进行综述,并简要介绍该属资源的利用现状,为今后我国虎耳草属的分类、育种及应用提供借鉴。结果表明:(1)虎耳草种质资源丰富,但属下系统演化关系仍存在诸多问题,有待整合形态和系统发育学手段开展系统而深入的研究。(2)该属品种主要通过杂交育种和变异筛选的方式育成,英国、捷克共和国、德国、荷兰为培育品种最多的国家。(3)我国对该属的品种选育工作起步晚,育成品种少且育种方式单一。
关键词: 虎耳草属, 栽培品种, 种质资源, 育种, 研究进展
中图分类号: Q945; Q949
文献标识码: A
文章编号: 1000-3142(2024)01-0193-14
Advances on taxonomy and breeding of
Saxifraga Tourn. ex L.
TANG Shimei1,2,3,4, ZHANG Xia2,4, CHEN Qianru2,4,
CAI Wenqi2,4, LI Yanlin2,4, TIAN Daike1,3*
( 1. Chenshan Science Research Center of CAS Center for Excellence in Molecular Plant Sciences, Shanghai Chenshan Botanical Garden, Shanghai
201602, China; 2. College of Horticulture, Hunan Agricultural University, Changsha 410128, China; 3. Shanghai Key Laboratory of
Plant Functional Genomics and Resources, Shanghai 201602, China; 4. Hunan Key Provincial Mid-Subtropical High Quality
Flower and Tree Breeding and Utilization Engineering Technology Research Center, Changsha 410128, China )
Abstract: Saxifraga Tourn. ex L. is rich in species diversity with about 440-500 species and broad distribution. The taxonomical issues of this genus have been continuously concerned by the botanists. Saxifraga has important ornamental and medicinal values. Some of European countries have made great efforts on the exploitation and utilization of its ornamental resources. China is one of the diversity centers of Saxifraga, but it has been lagged far behind Europe, USA and even Japan in exploitation and utilization on ornamental value of this genus. The world breeding work in Saxifraga started 150 years ago. By the end of 2022, the 1 692 names of cultivars had been on-line listed by The Saxifrage Society, but only one came from China. In this paper, the germplasm resources of Saxifraga, and progress on its classification and breeding are reviewed, and also the utilization of its ornamental resources is briefly introduced, which will provide an important reference for taxonomical research, breeding and application of this genus in China. The results are as follows: (1) Saxifraga is rich in germplasms, but many scientific issues remain in the evolutionary relationships of its subordinate systems, which require a systematic and in-depth investigation by integrating morphological and molecular biology methods. (2) The cultivars of this genus are mainly raised through cross-breeding and mutant selection, and the majority of cultivars are from the UK, Czech Republic, Germany, and the Netherlands. (3) The breeding of Saxifraga started very late in China, with only few cultivars from one breeding method.
Key words: Saxifraga, cultivar, germplasm resource, breeding, research progress
虎耳草属(Saxifraga Tourn. ex L.)是虎耳草科(Saxifragaceae)中最大的属,也是分类最为复杂、困难的属之一(潘锦堂,1992;张梦华,2018)。该属主要由耐寒性的多年生草本组成,稀一年或二年生草本,花瓣5,通常为辐射对称,稀两侧对称,雄蕊10,花丝棒状或钻形(潘锦堂,1992)。虎耳草属的很多种类为药材,在化学成分和药理活性方面,国内外对部分种类开展过比较深入的研究。其中,已知的药理活性包括抗菌抗病毒疗效、抗炎抗氧化功能以及抗突变、抗肿瘤、抗雌雄激素的作用,甚至有护肝、诱导成纤维细胞凋亡、降血糖、镇咳作用等(左国营等,2007;于淑玲,2009;李玉兰,2011)。挪威虎耳草(S. oppositifolia L.)的叶子被用作茶,其叶和花可食用,因其富含维生素C而作为因纽特人饮食中必不可少的一部分(Nyvoll, 2019)。此外,虎耳草屬的高山类群具有其他大部分高山植物所不具有的优良特性。例如,该属很多种类寿命长、可全年观叶、多花、花色多样(白、黄、浅黄、橙、红、淡紫、紫和粉色)、抗虫性好以及栽培条件下往往能保持野生状态下的植物形态,而许多其他高山植物在花园里的表现通常不如在野外(Sellars, 2019;Malcolm, 2008)。鉴于虎耳草属的优良观赏特性,在全球育种者的长期努力下,培育出大量品种并在园林中应用。截至2022年底,负责虎耳草属栽培品种国际登录的权威机构The Saxifrage Society(国际虎耳草协会)网站(https://www.saxifrage.org/)已收录了1 692个虎耳草属的品种名称。在国外,部分观赏价值高的虎耳草属种类和品种已被广泛应用于花境布置、岩石园点缀、盆栽观赏等,而国内对虎耳草属观赏价值的开发利用程度还远远不够,目前仅局限于虎耳草(S. stolonifera Curt.)一种。研究方面主要关注虎耳草属的药用价值,忽视了其观赏价值(Takeda et al., 2020;Kawahara et al., 2021)。近年来,虎耳草因其漂亮的株形、叶形、叶色和极强的栽培适应性等优点而在国内越来越多地被作为盆栽观赏、盆景点缀、岩石园和地被植物等进行应用(孔令亚等,2014;刘嘉等,2016)。但是,该属其他种类的观赏价值开发利用方面还是空白,至今还没有培育出一个杂交品种,与国外相比差距甚大。为了让科技工作者和大众尽快了解这种差距,促使我国园艺界对虎耳草属资源调查分类和其观赏价值开发利用的重视,本文对虎耳草属的全球资源及分类状况、虎耳草属的育种历史、进展和方法等进行分析总结,并对我国虎耳草属育种和观赏价值的开发利用提出一些有益思考和建议。
1 虎耳草属资源及分类
虎耳草属全球有440~500个种,约占虎耳草科的三分之二(Tkach et al., 2015)。虎耳草属以适应寒冷的多年生草本植物为主,广泛分布于北半球。该属起源于北美的落基山,向南扩散到南美洲的安第斯山和火地岛,在北部经白令海峡扩散到欧亚大陆。在这些地区,其高度的生物多样性可与蒿属(Artemisia L.)、薹草属(Carex L.)、毛茛属(Ranunculus L.)和早熟禾属(Poa L.)等相媲美(Ebersbach et al., 2016)。虎耳草属生于林缘、草原、冻土地带、岩坡石隙,其多样性中心包括中国—喜马拉雅山脉、高加索山脉、阿尔卑斯山脉以及北极的山区(Elven et al., 2011)。虎耳草属在生长习性、形态、营养特性、生殖特性以及花粉粒和种子的微观形态上都表现出显著多样性,因此该属在研究高山植物系统演化、物种进化等领域常作为模式类群(Ebersbach et al., 2018)。在繁殖方式方面,虎耳草属除了主要通过种子繁殖外,有些种类还可通过匍匐茎、球茎、珠芽等方式繁殖。该属的授粉方式通常为昆虫传粉,少数自交授粉(Zhmylev, 2001)。我国虎耳草属南北均产,主要分布于西南、青海和甘肃等省的高山地区;其中云南分布最多,约100种,多生长在海拔3 000 m以上的高山岩石上及石缝间(Kunming Institute of Botany, Chinese Academy of Sciences, 2006)。
虎耳草属组间、组内,或种下分类,均有植物学者关注研究,使用的分类方式包括传统形态分类、分子系统学(核糖体ITS片段、叶绿体片段、叶绿体基因组、ddRAD-seq等)及数量分类学(Zhang et al., 2015; Fujii et al., 2020)方式等。林奈(1753)正式建立了虎耳草属,当时该属仅含31种但无属下分类。Haworth (1803)首次对该属进行了属下等级分类,将其划分为6个组[Sect. Nudicaules Haw.、石荷叶组(Sect. Irregulares Haw.)、舌形组(Sect. Ligulatae Haw.)、山羊臭组(Sect. Ciliatae Haw.)、Sect. Lobatae Haw.、Sect. Stoloniferae Haw.]。虎耳草属最早的分类系统由Engler和Irmscher(1916—1919)建立,后来由Gornall(1987)修订,包括15个组、19个亚组和34个系,当前被国际虎耳草协会使用。近年来,虎耳草属分子系统的研究获得迅速发展,相关研究表明该属独立为一个单系,并与莛虎耳草属(Micranthes Haw.)互为姊妹支系(Xiang et al., 2012;Tkach et al., 2015;Deng et al., 2015),Tkach等(2015)利用254种虎耳草属植物在分子系统发育分析中确定了虎耳草属的主要分支,并与以往的3个分类处理进行了比较,确认了该属至少有13个组和9个亚组。对虎耳草属的组间关系研究表明,虎耳草组(Sect. Saxifraga)与山羊臭组互为姊妹单系(Vargas, 2000; Gao et al., 2015),舌形组与糙叶组 [Sect. Trachyphyllum (Gaudin) W. D. J. Koch]是多系类群(Dechaine et al., 2013;Dechaine, 2014)。Conti等(1999)根据ITS和matK基因对分布在欧洲的舌形组进行了分析,结果显示,舌形组是一个多系类群,该组的一些类群与垫状组(Sect. Porphyrion Tausch)聚成一支;张得均等(2008)根据分子系统地理研究成果表明山羊臭组分支内所涉及的5个亚组分为两大支。Zhang等(2020)利用 ITS、trnL-F、PSBA-trnH和matK重组了石荷叶组的系统发育关系,结果支持东亚的石荷叶组与北美的Sect. Heterisia Raf.各为一支。此外,近年来与该属近缘种间(Gao et al.,2017;Magota et al., 2018;Magota et al., 2021)或种内(Abbott & Comes, 2003;Jrgensen et al., 2006;Winkler et al., 2012;Magota et al., 2021)遗传分化相关的研究也有报道,其中涉及最多的物种包括广泛分布于欧洲(西班牙、意大利和希腊北部)、亚洲(喜马拉雅山)和北美(南至科罗拉多)的挪威虎耳草,该种有8~9个亚种(Malcolm, 2008),以及广泛分布于日本的齿瓣虎耳草(S. fortunei Hook. f.)等。
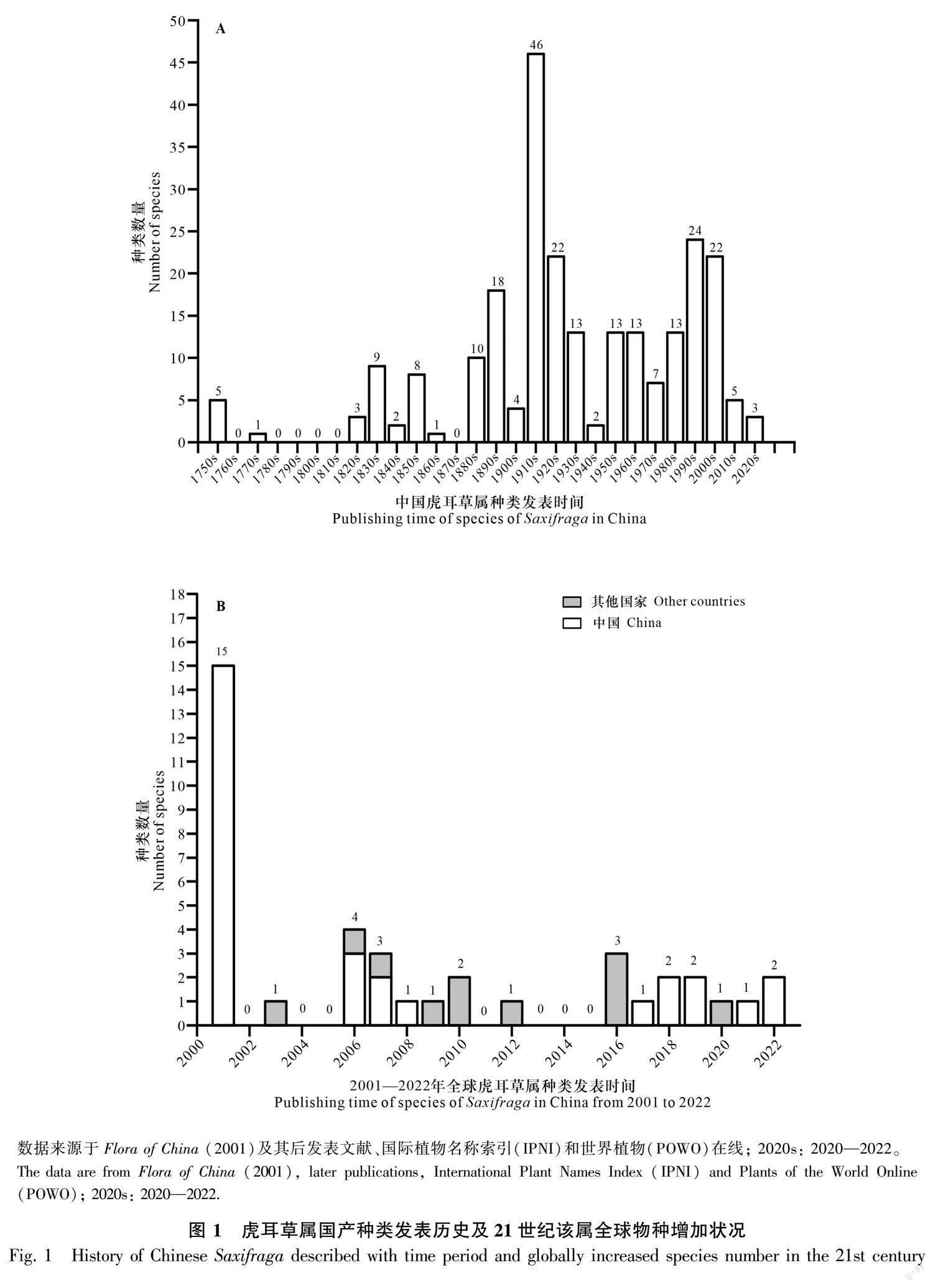
在我国,虎耳草一名始载于南宋医药学家王介著的《履巉岩本草》(张慧等,2021)。1992年潘景堂确定我国有虎耳草属植物203种,并将其分为8个组:双喙组 [Sect. Birostres (Gornall) C. Y. Wu & J. T. Pan]、斑点组 [Sect. Punctatae (Engl.) J. T. Pan]、石荷叶组、球茎组(Sect. Mesogyne Sternb.)、小花组 [Sect. Micranthes (Haw.) D. Don]、山羊臭组、舌形组和点地梅形组(Sect. Aretiaria Sternb.)(潘锦堂,1992)。Flora of China (FOC)记载国产该属植物216种,重新分为7组:小花组、石荷叶组、糙叶组、山羊臭组、球茎组、垫状组、虎耳草组(Pan et al., 2001),同以往相比变化较大。在物种方面,国产虎耳草属已有245种发表(图1:A),其中FOC中收录了216种,2001—2022年发表了29种(图1:B)。挪威虎耳草在1753年被命名,是国产虎耳草属最早发表的种类,这与林奈命名虎耳草属的时间相同(Linnaeus, 1753)。此后,1760—1820年仅有1个虎耳草属种类被命名;1910—1920年是命名发表的高峰期,共有46种被发表,与虎耳草属最早的分类系统建立时间相对应(Engler & Irmscher, 1958;Engler, 1916—1919);第二个发表高峰期为1990—2000年,共有24种被描述发表。
从全球范围来看,根据国际植物名称索引(International Plant Names Index, IPNI)(https://www.ipni.org/)和世界植物在线(Plants of the World Online, POWO)(https://powo.science.kew.org/)的数据查询结果(图1:B,表1),2001—2022年共有40个种类发表,其中最多的一年(2001年)发表15种,均来自中国,有7年(2002,2004,2005,2011,2013—2015)全球没有种类发表。在最近这22年期间,我国发表29种,其中2001年发表数量较集中,此后每年发表的种类很少。
2 虎耳草属育种进展
2.1 育种历史、品种数量及其特点
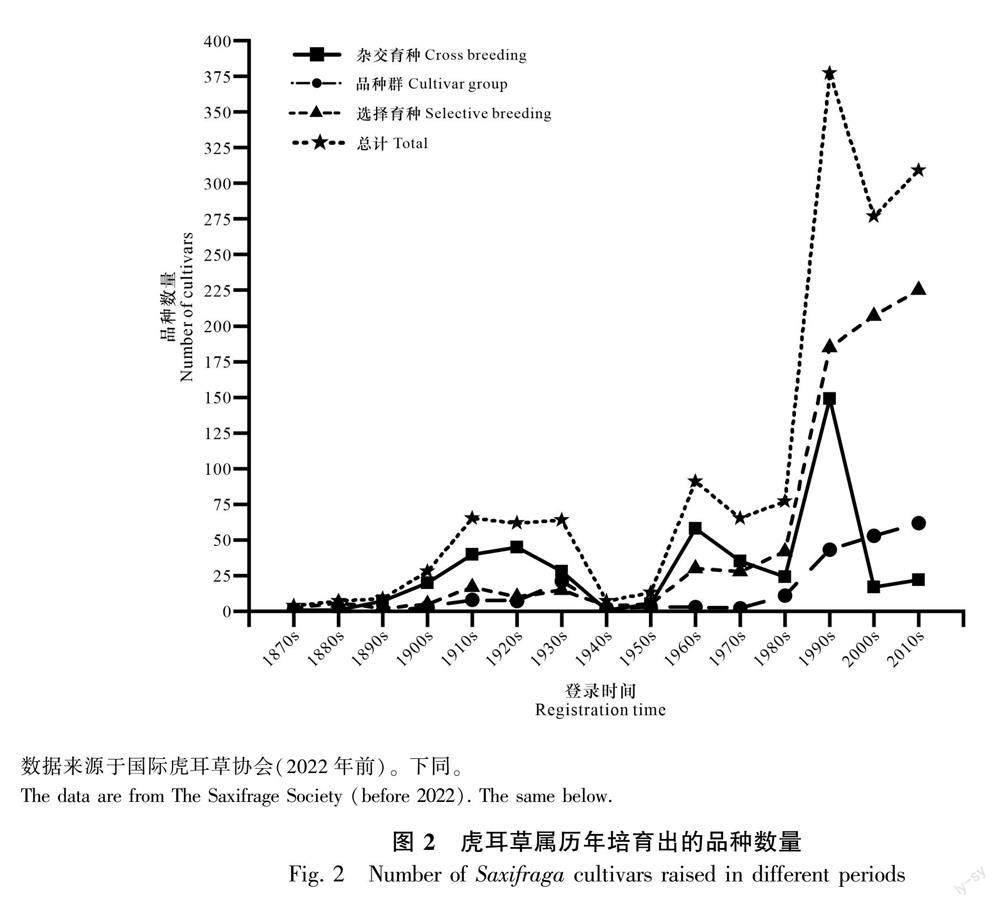
虎耳草属的育种历史已有150多年。通过对国际虎耳草协会的品种数据库进行检索和统计分析发现,共检索到1870—2020年有培育时间记载的虎耳草属品种1 455个,其中,奥地利的Kerner和Anton于1870年培育的‘Guthrieana是最早有明确育种者姓名及品种培育年份记载的品种。按照每10年培育出的品种数量统计发现波动较大,如1870—1880年培育的虎耳草属品种最少,仅4个;随后品种培育总数逐渐增加,到1930—1940年和1940—1950年又分别减少到7和13个;而1990—2000年培育的品种最多,达377个(图2)。自1890年以来培育的虎耳草属品种数量占比66.19%,相比1890年以前,这是一个惊人的数据,说明了近40年以来该属品种培育的快速興起。
根据不同虎耳草属品种的来源对其进行划分,可分为3类,即来源于直接选育(920个)、杂交(454个)和品种群(296个),各占比54.66%、27.75%和17.59%。1880—1980年育出的虎耳草属品种主要来源于杂交选育;1980—2000年育出的虎耳草属品种主要来源于原种的直接选育(434个)和杂交育种(190个),各占比59.37%和25.99%;2000年之后以选择育种方式为主(图2)。
按照品种所培育的国家分析,据不完全统计共有来自16个国家培育了1 487个虎耳草属品种。其中,英国、捷克共和国、德国、荷兰和美国培育的品种数分别为609、390、197、120和78个,合计占总数的93.75%;其他国家培育的品种少,如意大利、日本、瑞士和爱尔兰分别培育了27、27、19、6个品种;加拿大、奥地利和法国仅分别培育了5、3、2个品种,中国、挪威、瑞典和比利时均只分别培育了1个品种(图3)。
根据不同年份和国家对虎耳草属品种进行检索,收录品种中均有年份和国家记载的虎耳草属品种共计1 380个。从育种历史上看,育种历史最为悠久的国家有英国、瑞士和奥地利,在1870—1880年就有虎耳草属的品种选育;其次,德国的虎耳草属品种记录首次出现在1890—1900年;随后,爱尔兰和法国在1910—1920年有虎耳草属品种育出。从不同国家所培育的虎耳草属品种数量比较,英国排名第一,共培育了556个(40.29%);虽然捷克共和国在1930—1940年才出现虎耳草属品种记载,时间均晚于上述欧洲6国,但其培育的品种总量仅次于英国,为383个(27.75%);排在第三位的为德国,共计168个(12.17%);而瑞士和奥地利虽然在虎耳草属的育种方面起步早,但育出的品种总数分别仅有14和2个。英国和德国分别自1870年和1890年以来均有虎耳草属品种培育出;荷兰和美国培育虎耳草属品种起步较晚,分别于1990年和1960年才有品种出现,但品种总数分别居第4、第5位。特别是荷兰,仅1900—2020年30年间的品种数就达到全球第4位,可见该国的育种发展速度之快。反观我国,虎耳草属品种的首次登录时间为2014年(Tian, 2014),并且为该属首个有正式名称的品种。因此,从育种历史和培育的品种数量上看,我国与发达国家相差甚远。
2.2 虎耳草属的育种方式
2.2.1 直接选育 直接选育是培育虎耳草属新品种最常用、最有效的方法。特别是自1990年以来,通过直接选育的方式将野生变种、变型或变异个体培育成符合观赏目标的园艺品种成为虎耳草属的主要育种方法。通过此方式培育的品种有912个,占总品种数的53.90%,分别来源于Sect. Porphyrion Tausch(591个)、舌形组(189个)、石荷叶组(106个)、虎耳草组(24个)和Sect. Gymnopera D. Don(2个)5个组。国际虎耳草协会对部分直接选育的品种来源的记录并没有都具体到种,甚至部分只记录到组、亚组或系,推测其可能是由于原始育种记录信息不全的缘故,尤其是早期培育的品种。
垫状组大约有110个种分布于欧洲、喜马拉雅山脉和中国西南部,大多是株高仅几厘米的垫状植物(Brochmann & Hapnes, 2001)。值得注意的是,表2中的S. burseriana和S. marginata都属于Ser. Kabschia,而Ser. Kabschia和Ser. Engleria 都是Subsect. Porophyllum的下级分类单位,挪威虎耳草则来自于Subsect. Oppositifoliae。由此计算出表中对Subsect. Porophyllum直接选育的品种有527个,说明育种家热衷于通过对Subsect. Porophyllum直接选育来筛选新品种,也说明Subsect. Porophyllum的原种具有较高的观赏价值且变异丰富;挪威虎耳草也是来自于Sect. Porphyrion的下级分类单元对叶亚组中值得关注的类群,由此种筛选出28个品种。
舌形组包括在欧洲和非洲北部的约10个种(Brochmann & Hapnes,2001),其中锥花虎耳草(S. paniculata)和匙叶虎耳草(S. cochlearis)的分布非常广泛,其他种的分布则呈现破碎化(Malcolm, 2008)。按照Gornall(1987)的分类,舌形组中大多数都有美丽的白花,但也有一些开红色或粉色花的优良品种,如‘Fosters Red‘Hare Knoll Beauty ‘Rosea (Sellars, 2019)。通过对该组进行直接选育所产生的品种共166个,包括从锥花虎耳草和匙叶虎耳草分别直接选育的53和12个品种(表2)。锥花虎耳草为多年生植物,非常耐寒、不易染病,也不需要过多打理(Malcolm, 2008),这可能是它能被广泛选育并应用于园林中的重要原因之一。虽然舌形组有约10个种,但对该组的资源开发利用与有近110个种的垫状组相比并不逊色。
石荷叶组有10~20种,分布于亚洲东部,直立且株高(含花序)可达40 cm,有大的带叶柄的基生叶(Gornall, 1994)。据作者不完全统计,虎耳草属的品种中通过齿瓣虎耳草选择育种培育出的品种共计86个。1993—2015年由英国皇家园艺学会(Royal Horticultural Society,RHS)给51个虎耳草属品种颁发了花园优秀奖(Award of Garden Merit,AGM),其中70.59%的品種来自选择育种,有14个品种来自齿瓣虎耳草,其中13个品种在2015年获奖。不仅如此,由于日本的27个品种中有24个来自齿瓣虎耳草,其原因可能是齿瓣虎耳草的变种——大文字草(S. fortunei var. incisolobata Engl. & Irmsch.)的野生资源分布广泛,在日本的北海道、本州、四国、九州等地均产,多样性高,自然变异大,叶、花形、色各异,花大而多,因此大文字草品种繁多,美不胜收(图4:A)。大文字草的花瓣不等长,上侧3瓣明显短于下侧2瓣,从而组成一个“大”字,由此得名(刘嘉等,2016)。同为石荷叶组的虎耳草不仅是药材还是一种野菜,如在中国有虎耳草炒鸡蛋;在日本则拥有更多千奇百怪的做法,如虎耳草叶天妇罗(一种油炸食物)、凉拌、煮汤等,人们对它味道的评价是具有一股青草香,颇为奇特。日本育种者培育的虎耳草品种‘七变化(‘Nana Henge)、‘御所车(‘Tricolor) 和‘极姬雪の下(‘Gokuhime Yukinoxita)等很受国内外爱好者的欢迎(图4:B, C, F)。‘七变化在冬季的新叶为纯白色,随着生长叶片转为绿白相间,老叶会泛起红晕,甚至有全株雪白的情况,变化多端,十分新奇;‘御所车的叶片有不规则的白色或浅粉色镶边,叶中部绿色或绿中带粉,典雅华美;‘极姬雪の下植株矮小,叶片小而密集,最大叶仅如无名指指甲般大小,娇小玲珑,十分可爱,是做微型盆景的上佳材料(陶隽超等,2016)。除此之外,Tian(2014)从野外发现的虎耳草变异株筛选培育而成的新品种‘黑魁(‘Hei Kui),叶片莲座状紧凑排列,植株健壮、株型优美,其叶莲座状排列,叶表面脉间规则分布大小不等的纺锤形黑眉状斑纹,观赏性高,极具园林应用潜力(图4:G),在北京举行的“第二届园林植物新品种新技术交流会”被评为最具潜力新品种(张哲,2016)。‘黑魁虎耳草也是我国首个自主培育和首个国际登录的虎耳草品种(田代科,2015)。此外‘雪纹(‘Xue Wen)和‘天目恩赐(‘Tianmu Enci)为上海辰山植物园2020年申报完成国际登录的品种(图4:D, E),前者叶片表面沿着叶脉具有宽窄不等的白色条纹,该品种尽管在我国园林应用的历史悠久,但在完成国际登录前一直没有具体的品种名,同更为常见的纯绿色叶片个体一起在民间统称为虎耳草;后者为新选育出的新品种,叶片表面脉区白纹狭窄,脉间为深绿和黑条斑(但对光照较敏感),密被粉红色长毛,生长旺盛,观赏价值高,尚未大量推广(田代科,2022)。
虎耳草组分布最广(欧洲、非洲北部、亚洲、北美和南美洲),该组由70~85个种组成;Sect. Gymnopera(4个种)都分布在欧洲的山区(Webb & Gornall, 1989)。虎耳草组的物种数虽然远多于舌形组和石荷叶组,并且在物种分布范围上占优势,但数据显示通过直接选育出的该组品种数却相对较少。实际上, 全球育种者统一把该组部分观赏价值较高的种或杂交品种归为了Saxifrage Mossy Group栽培群。
2.2.2 杂交育种 杂交育种是选育新品种的重要培育方式之一。在已登录的虎耳草属所有杂交品种中,垫状组的组内种间或品种间杂交产生的杂交品种共计456个(98.06%),并且均来自Subsect. Porophyllum 的3个亚系间或亚系内(Kabschia、Engleria、Tetrameridium)杂交,与Malcolm(2008)的专著记载相同;舌形组的组内种间或品种间的杂交品种共计仅9个(1.94%),来自5个杂交组合,但未见组间杂交品种的记载。垫状组共登录有92个杂交组合,使用最多的是杂交组合是S. × megaseaeflora(S. aretioides、S. burseriana、S. lilacina、S. media),共培育出67个品种;其次是S. × poluanglica(S. aretioides、S. lilacina、S. media、S. poluniniana)共培育出34个品种;居第3位的杂交组合为S. × anglica (S. aretioides、S. lilacina、S. media),共育出25个品种(表3)。来自捷克共和国的Radvan HornAy'和Karel Mirko Webr发表的Porophyllum Saxifrages曾详细地介绍了该亚组3个亚系的种及杂交品种(Malcolm, 2008)。
虎耳草属的原生种作为亲本参与杂交培育的品种数如表4所示,其中涉及S. burseriana(1753年发表)的品种最多。14个参与杂交的原生种中除了近优越虎耳草(S. corymbosa)外,其余13个原生种在我国均不产,说明国外主要利用当地的种质资源进行新品种培育,我国的虎耳草属种质资源很少被欧美用于育种。研究表明,垫状组的自然杂交种分布于欧洲和高加索地区,喜马拉雅地区也可能广泛存在该组的自然杂交种(Malcolm, 2008)。
英国育种家Gornall(1994)在Some aspects of hybridisation in the genus Saxifraga一文中详细介绍了虎耳草属人工杂交育种方法的步骤。用酒精给镊子消毒,并在母本的花药成熟前去雄,使用空茶袋、薄纱袋甚至纸袋来套袋以避免去雄的花朵被污染;当母本的柱头变得潮湿或膨胀时,用镊子或清洗干净的毛笔取父本的成熟花粉在母本柱头表面轻轻摩擦,每次授粉前都须给镊子消毒;授粉后将母本重新套袋,挂上标签并在笔记本上记录日期,以及父本母本的信息等。虎耳草属植物的花粉产生后,会在几天或几周后失活,若花粉被弄湿,花粉活力仅可保持几个小时。一般来说双核花粉的花粉活力比三核花粉持续时间更长。虎耳草属中除山羊臭组为三核花粉外,其他均为双核花粉。因此,除山羊臭组外,其他虎耳草属物种的花粉均适合低温条件下保存以后使用。可在花药裂开前收集花药保存在干燥的小瓶中放入冰箱冷藏,这样花粉活力可保持一个月,冷冻起来可以保持更长时间,达一年,但不同种间会存在差异。此外,Richard Gornall还介绍了虎耳草属人工杂交的实验设计及衡量杂交是否成功的标准等。
2.3 虎耳草属品种群
《国际栽培植物命名法规》(第九版)规定:品种群是基于一定相似性的品种、植物个体或植物集合体的正式等级,品种群的品种可能来自直接选育,也可能来自杂交(向其柏和臧德奎,2016)。国际虎耳草协会数据库中共有来自68个品种群的296个品种,占总品种数的17.49%。品种数排在前三位的品种群分别为Saxifrage Mossy Group、Saxifrage Blues Group和Saxifrage London Pride Group,分别有138(46.62%)、24(8.11%)和22个品种(7.43%),其他品種群的品种总数均小于等于6个。Saxifrage Mossy Group由虎耳草组的大部分原种和杂交种组成,该栽培群植株低矮,叶片细小紧凑,形似苔藓植物,因此通常被称为苔藓虎耳草,该类品种花色丰富,有花色纯白的‘Fleece,白色花瓣上带明显浅绿脉络的‘Schwefelblüte等;花色深浅不一的红色系,如浅粉色的‘Knapton Pink、粉红的 ‘Glassels Crimson、玫红的‘Triumph、深红的‘Welsh Dragon等。这类虎耳草形态比较接近于生长在高山的部分种类,多数被应用于岩石园或盆栽,其花量大,花色鲜艳,少数当成地坛草花。Saxifrage Blues Group的叶莲座状着生,叶细小,紧凑排列在地面形成一个半球形,镶嵌着生粉色、橘红或玫红的花朵,花瓣圆形,其中‘Satchmo的花瓣还有浅波浪状缺刻。Saxifrage London Pride Group来自Sect. Gymnopera(除S. cuneifolia外),具长叶柄,叶片深绿或浅绿,圆形或匙形,具缘齿,椭圆形的白色花瓣上点缀着粉红色和黄色的斑点,雄蕊和花药为粉色。
2.4 虎耳草属品种的杰出育种者
欧洲的育种者对虎耳草属的育种贡献最大,其中德国的育种者Sündermann和Franz共同培育出115个品种,为育种者之最,所育品种数占德国的68.45%, 包括本国的首个虎耳草品种。这些品种中,有88个来源于杂交育种,26个来自选择育种,1个来自栽培群。品种数第二多的育种者为捷克共和国的Lang和Karel,两位在1990—2020年共同培育出100个品种,最近一次记载为2017年,该年培育出的品种有7个。这些品种中,通过栽培群选育的最多,有71个,来自杂交育种的有16个,源于选择育种的有13个。荷兰的Moerland和Bart是品种数量第三多的育种者,二者于2000—2020年培育出99个品种,占荷兰总数的82.50%。
3 问题与展望
3.1 虎耳草属的分类
作为研究高山植物系统演化、物种进化和多样性等领域的特征类群(邓家彬,2015),近二十年来,虎耳草属的分子系统学研究发展迅速,但该属的属下系统演化关系仍存在诸多问题(胡晶晶,2020),组的划分存在不确定性,并不能真实地反映虎耳草属属下等级的自然发生规律,有待整合形态和系统发育学手段开展系统而深入的研究。在资源调查和新物种描述方面,进入21世纪后,除了2001年发表15种最多外,此后每年发表的种类不超过4种,甚至这期间有7年无新种描述发表,说明21世纪全球虎耳草属的经典分类学研究几乎停滞不前。云南为我国虎耳草属种类分布最多的省份,2001年发表的15种均分布在该省,显然同该省具有丰富的生物多样性相关。21世纪我国发表虎耳草属种类29个,其中石荷叶组由原来7种增加到现在的15种,新增加的种类发现于广西、广东、湖南、江西和甘肃,因此推测该组还可能有新种待调查。此外,2001年发表的15种国产虎耳草属种类均来自从事虎耳草专属植物分类研究的同一学者,而此后发表的种为非虎耳草专属的不同学者参与的工作,说明国内现已缺少专门针对这一类群开展研究的分类学者,我国的虎耳草属家底尚未清楚,急需年轻分类学家针对该属开展系统的资源调查、分类及其修订工作,而面向全球开展虎耳草属的资源调查和分类研究可能面临更长期、更艰巨的挑战。
3.2 国际虎耳草属的育种趋势及展望
根据国际虎耳草协会的数据库数据统计,在有年份记载的品种数中,仅在1990—2022年培育的虎耳草属品种数占比66.19%,并且在将花卉业作为支柱产业的荷兰自1995年培育第1个虎耳草属品种以来,陆续培育出大量优良虎耳草属品种,说明了近半个世纪虎耳草属育种的兴起。但是,该属的育种大都局限于欧美国家,亚洲除日本外还处于落后阶段,目前其他洲的育种还是空白。现有记载的虎耳草属品种主要通过杂交育种和变异筛选的方式育成。杂交育种主要利用Subsect. Porophyllum 的3个系间或系内杂交,组间或属间杂交未见报道,组间或属间杂交可能成为未来的重要育种方向。当前,传统的育种方式还是主流,但将来一定会选择利用诱变、分子技术等进行辅助育种,而这些育种方法需要不断摸索。在育种材料选择上,国际交流的日趋频繁带动了种质资源的快速流通,为育种家们提供了亲本选择的便利。但是,在引进国外的优良原种和品种时,除了主要考虑其观赏价值以外,还应充分考虑到品种对本土气候和土壤条件的环境适应性。
3.3 中国虎耳草属的育种现状及展望
相比国际上的虎耳草属育种历程长和品种数量多,虽然中国开展该属的育种工作还很少且才刚开始不久,但也说明育种潜力巨大,因为中国的气候和资源多样性均很高。中国虎耳草属的育种存在明显不足:(1)育种起步晚,2013年才完成该属品种的首次国际登录,中国也才有了第一个虎耳草品种名(‘黑魁);(2)品种数量太少,截至2022年底仅有3个品种(其中1个已被英国虎耳草协会数据库收录);(3)育种材料及方式单一,品种仅通过野外变异筛选培育而成,杂交育种还没有起步。育种落后同我国丰富的虎耳草属种质资源形成了极大反差。系统全面地调查掌握我国虎耳草属的种质资源对该属的新品种选育至关重要(程金水,2010),也是开展活体资源收集和高效育种的前提。虎耳草属的石荷叶组(15~20个种)仅在东亚分布(Ebersbach et al., 2017;Magota et al., 2021),其中不乏观赏价值高的育种材料,如虎耳草、齿瓣虎耳草及近几年发表的部分石荷叶组新种,其中前两种的种内多样性很高,直接选育出品种的概率大。此外,被西方国家广泛应用于虎耳草属品种选育的垫状组在喜马拉雅山脉和中国西南部等地有分布,可作为驯化和杂交亲本的优良材料。因此,在引进国外优良虎耳草属育种材料作为亲本的同时,也应充分挖掘本国的丰富资源,开展选择驯化、杂交、理化诱变,甚至分子手段等综合育种方式,培育出适合中国不同区域气候特点的优良品种。
3.4 虎耳草属观赏资源的开发及应用
除了品种培育外,欧美国家还十分重视对观赏虎耳草属植物的开发和推广应用。例如,2009年4月,布拉格岩石园俱乐部(Prague Rock Garden Club)举办的虎耳草属春季展览会展出了大约700株虎耳草属植物及栽培品种,此次展览还包括了相关讲座和种子交换等活动。国际虎耳草协会专为虎耳草属育种者设立了塞尔吉奥·巴奇杯(Sergio Bacci Cup),以激励育种者培育出优秀新品种。由此可见,虎耳草属育种及推广已在欧美国家开展得很好。此外,国外很多植物园也重视虎耳草属植物资源的收集和展示,这些植物园通常建设有岩生植物专类园,其中就点缀着很多虎耳草属种类和品种,如英国的邱园(Kew Gardens)、美国的丹佛植物园(Denver Botanic Gardens)、新西兰的达尼丁植物园(Dunedin Botanic Garden)等,关于一些虎耳草属类群栽培方式的专著也有发行。对比国外,我国对虎耳草属植物的关注点大部分集中在少数种类的药理活性上(张慧等,2021),在本属的观赏价值的开发利用上存在较大差距。近年来,国内关于虎耳草在林下地被绿化、案头装饰、岩石园、墙垣、野趣园、垂直绿化中的阴面壁面绿化、城市立交垂直绿化方面和阴面垂直花坛上的应用已有报道(孔令亚等,2014;许红娟和陈之林,2015),但目前国内市场上的虎耳草属品系较少,多样性低,仅有来自日本石荷叶组和英国Saxifrage Mossy Group的少数品种。
3.5 虎耳草属新品种DUS测试指南创制
虎耳草属品种虽然通过国际虎耳草协会进行登录后使其名称合法化,便于生产和科研等活动交流,但同其他植物栽培品种一样,登录后的品种并不受法律保护。目前,国际植物新品种保护联盟(International Union for the Protection of New Varieties of Plants,UPOV)和任何一个国家都还没有创制出版《虎耳草属新品种DUS测试指南》,并且虎耳草属还不在我国植物新品种保护名录中。因此,新品种很难获得保护权,这将严重阻碍育种者的积极性。只有尽快将虎耳草属纳入国家新品种保护目录,及时创制出该属的新品种DUS测试指南并申请新品种权保护,才能切实保障虎耳草属植物育种者的权利,促进新品种培育及合理推广应用,有利于我国虎耳草园艺产业的健康发展。
参考文献:
AB BOTT RI, COMES HP, 2003. Evolution in the arctic: a phylogeographic analysis of the circumarctic plant, Saxifraga oppositifolia (purple Saxifraga) [J]. New Phytol, 161(1): 211-224.
BROCHMANN C, HAPNES A, 2001. Reproductive strategies in some arctic Saxifraga (Saxifragaceae), with emphasis on the narrow endemic S. svalbardensis and its parental species \[J\]. Bot J Linn Soc, 137(1): 31-49.
CHEN HL, XU WB, ZHAO WY, et al., 2022. Morphological and molecular identification for Saxifraga yangshuoensis (sect. Irregulares, Saxifragaceae), a new species from Guangxi, China [J]. Taiwania, 67(4): 603-608.
CHENG JS, 2010. Landscape plant genetic breeding [M]. Beijing: China Forestry Press: 1-468. [程金水, 2010. 園林植物遗传育种学 [M]. 北京: 中国林业出版社: 1-468.]
CONTI E, SOLTIS DE, HARDIG TM, et al., 1999. Phylogenetic relationships of the silver saxifrages (Saxifraga sect. Ligulatae Haworth): implications for the evolution of substrate specificity, life histories, and biogeography [J]. Mol Phylogenet Evol, 13(3): 536-555.
DECHAINE EG, ANDERSON SA, MCNEW JM, et al., 2013. On the evolutionary and biogeographic history of Saxifraga sect. Trachyphyllum (Gaud.) Koch (Saxifragaceae Juss.) [J]. PLoS ONE, 8(7): e69814.
DECHAINE EG, 2014. Introducing the spotted saxifrages: Saxifraga sect. Bronchiales, sect. nov. (Saxifragaceae) [J]. Rhodora, 116(965): 25-40.
DENG JB, DREW BT, MAVRODIEV EV, et al., 2015. Phylogeny, divergence times, and historical biogeography of the angiosperm family Saxifragaceae [J]. Mol Phylogenet Evol, 83(9): 86-98.
DENG JB, 2015. Phylogeny, divergence time, and biogeography of Saxifragaceae [D]. Yaan: Sichuan Agricultural University: 1-106. [鄧家彬, 2015. 虎耳草科的分子系统发育、时间分化及其生物地理学研究 [D]. 雅安: 四川农业大学: 1-106.]
EBERSBACH J, MUELLNER-RIEHL A, FAVRE A, et al., 2018. Driving forces behind evolutionary radiations: Saxifraga section Ciliatae (Saxifragaceae) in the region of the Qinghai-Tibet Plateau [J]. Bot J Linn Soc, 186(3): 304-320.
EBERSBACH J, MUELLNER-RIEHL AN, MICHALAK I, et al., 2016. In and out of the Qinghai-Tibet Plateau: divergence time estimation and historical biogeography of the large arctic-alpine genus Saxifraga L. [J]. J Biogeogr, 44(4): 900-910.
EBERSBACH J, SCHNITZLER J, FAVRE A, et al., 2017. Evolutionary radiations in the species-rich mountain genus Saxifraga L. [J]. BMC Evol Biol, 17(1): 199.
ELVEN R, MURRAY D, RAZZHIVIN VY, et al., 2011. Annotated checklist of the Panarctic Flora (PAF) vascular plants, version 1.0 [M]. Oslo: National Centre of Biosystematics, Natural History Museums and Botanic Garden, University of Oslo.
ENGLER A, IRMSCHER E, 1958. Saxifragaceae-Saxifraga [M]. Leipzig: Verlag von Wilhelm.
ENGLER A, IRMSCHER E, 1916—1919. Saxifragaceae: Saxifiraga [M]//Das Pflanzenrei ch. Leipzig: Wilhelm Engelmann: 1-709.
FUJII S, TOKUOKA Y, MAKI M, 2020. Leaf morphology of Saxifraga fortunei var. obtusocuneata and var. suwoensis (Saxifragaceae) [J]. Acta Phytotax. Geobot, 71(3): 231-242.
GAO QB, LI Y, GENGJI ZM, et al., 2017. Population genetic differentiation and taxonomy of three closely related species of Saxifraga (Saxifragaceae) from Southern Tibet and the Hengduan Mountains [J]. Front in Plant Sci, 8: 1325.
GAO QB, LI YH, GORNALL RJ, et al., 2015. Phylogeny and speciation in Saxifraga sect. Ciliatae (Saxifragaceae): Evidence from psbA-trnH, trnL-F and ITS sequences [J]. Taxon, 64(4): 703-713.
GAO Q, ZHANG Z, CHEN S, et al., 2018. A new species of Saxifraga in section Ciliatae subsection Gemmiparae (Saxifragaceae) from Sichuan Province, China [J]. Phytotaxa, 333(2): 228-234.
GORNALL R, 1994. Some aspects of hybridisation in the genus Saxifraga [J]. Saxifrage Mag Art. https://www.saxifraga.org/plants/saxifrage-magazine-articles/36-some-aspects-of-hybridisation.
GORNALL RJ, 1987. An outline of a revised classification of Saxifraga L. [J]. Bot J Linn Soc, 95: 273-292
HAWORTH AH, 1803. Miscellanea Naturalia: Sive Dissertationes Variae and Historiam Naturalem Spectantes [M]. Londini: Typis J. Taylor.
HU JJ, 2020. Studies on karyotypes and genome sizes of Saxifraga in the alpine belt from Hengduan Mountains [D]. Chongqing: Chongqing Normal University: 1-83. [胡晶晶, 2020. 橫断山高山带虎耳草属核型与基因组大小研究 [D]. 重庆: 重庆师范大学: 1-83.]
JRGENSEN MH, ELVEN R, TRIBSCH A, et al., 2006. Taxonomy and evolutionary relationships in the Saxifraga rivularis complex [J]. Syst Bot, 31(4): 702-729.
KAWAHARA T, ITO A, KISO A, et al., 2021. Inhibitory effect of strawberry geranium (Saxifraga stolonifera) on Toll-like receptor 2-mediated inflammatory response in human skin keratinocytes [J]. J Ethnopharmacol, 275 (2021): 114039.
KONG LY, OU GJ, WANG CH, 2014. Application of Saxifraga stolonifera Curt. to landscape architecture [J]. Shaanxi For Sci Technol, (2): 77-78. [孔令亚, 欧刚军, 王春辉, 2014. 虎耳草在园林中的应用 [J]. 陕西林业科技, (2): 77-78.]
Kunming Institute of Botany, Chinese Academy of Sciences. 2006. Flora of Yunnanica [M]. Beijing: Science Press: 1-227.
LINNAEUS C, 1753. Linnaeus C: Species Plantarum Stockholm [M]. Stockholm: Impensis Laurentii Salvii: 398-405.
LIU J, TAO JC, WU M, 2016. Saxifrage fortunei var. incisolobata [J]. Flower Plant & Penjing (Bonsai Stone Appreciation), (1): 78-79. [刘嘉, 陶隽超, 吴鸣, 2016. 大文字草 [J]. 花木盆景(盆景赏石), (1): 78-79.]
LI YL, 2011. Studies on the chemical constituents and activity of Saxifraga tangutica Engl. [D]. Lanzhou: Lanzhou University of Technology: 1-49. [李玉兰, 2011. 藏药甘青虎耳草的化学成分及药理作用研究 [D]. 兰州: 兰州理工大学: 1-49.]
MALCOLM MG, 2008. Saxifrages: The definitive guide to 2000 species, hybrids & cultivars [M]. London: Timber Press: 1-384.
MAGOTA K, SAKAGUCHI S, AKAI K, et al., 2018. Diversity of Saxifraga acerifolia and S. fortunei based on nuclear and chloroplast microsatellite markers [J]. Bull Nat Mus Nat Sci, Ser B, 44(2): 85-96.
MAGOTA K, SAKAGUCHI S, HIROTA SK, et al., 2021. Comparative analysis of spatial genetic structures in sympatric populations of two riparian plants, Saxifraga acerifolia and Saxifraga fortunei [J]. Am J Bot, 108(4): 680-693.
MAGOTA K, SAKAGUCHI S, LEE JS et al., 2021. Phylogeographic analysis of Saxifraga fortunei complex (Saxifragaceae) reveals multiple origins of morphological and ecological variations in the Japanese Archipelago [J]. Mol Phylogenet Evol, 163(2021): 107230.
NYVOLL K, 2019. Arctic cuisine-indigenous use [J]. Saxifrage Mag Art. https://www.saxifraga.org/plants/saxifrage-magazine-articles/54-arctic-cuisine-indigenous-use
PAN JT, 1992. Flora Reipublicae Popularis Sinicae: Tomus 34(2) [M]. Beijing: Science Press: 35-231. [潘锦堂, 1992. 中国植物志: 第三十四卷, 第二分册 [D]. 北京: 科学出版社: 35-231.]
PAN JT, GORNALL RJ, OHBA H, 2001. Saxifragaceae, in Flora of China [M]. Beijing: Science Press; St. Louis: Missouri Botanical Garden Press, (8): 280-344.
PAN JT, MEI LJ, CHEN SL, et al., 2006. Saxifraga banmaensis and S. dingqingensis, two new species of the Saxifragaceae from China [J]. Acta Phytotaxon Sin, 44(4): 443-446.
PAN JT, MEI LJ, ZHANG DJ, et al., 2007. Two news pecies of Saxifraga (Saxifragaceae) from southwestern China [J]. J Bot Nomenclature, 17(4): 512-515.
SELLARS D, 2019. The joy of Sax [J]. Saxifrage Mag Art. https://www.saxifraga.org/plants/saxifrage-magazine-articles/57-the-joy-of-sax.
TAKEDA Y, MURATA T, JAMSRANSUREN D, et al., 2020. Saxifraga spinulosa-derived components rapidly inactivate multiple viruses including SARS-CoV-2 [J].Viruses, 12(7): 699.
TAO JC, LIU J, WU M, 2016. Beautiful Saxifraga stolonifera [J]. Flower Plant Penjing (Bonsai Stone Appreciation), (6): 68-69. [陶隽超, 刘嘉, 吴鸣, 2016. 缤纷“虎耳草” [J]. 花木盆景(盆景赏石), (6): 68-69.]
TIAN DK, 2014. Saxifraga stolonifera ‘Hei kui [J]. Saxifrage Mag, (22): 48-49
TIAN DK, 2015.‘Hei kui, the first new cultivar of Saxifraga in China [J]. Chin Flower Bonsai, (5): 4-5. [田代科, 2015. 中國首个虎耳草属国际登录新品种——‘黑魁 [J]. 中国花卉盆景, (5): 4-5.]
TIAN DK, 2022. Three international registered cultivars of Saxifraga L. in China [J]. Flower Plant Penjing, (5): 36-39. [田代科, 2022. 我国虎耳草国际登录的三个品种 [J]. 花木盆景(花卉园艺), 2022(5): 36-39.]
TKACH N, RSERM, MIEHE G, et al., 2015. Molecular phylogenetics, morphology and a revised classification of the complex genus Saxifraga (Saxifragaceae) [J]. Taxon, 64(6): 1159-1187.
VARGAS PA, 2000. Phylogenetic study of Saxifraga sect. Saxifraga (Saxifragaceae) based on nrDNA ITS sequences [J]. Plant Syst Evol, 223(1): 59-70.
WANG FG, YU SY, XING FW, 2008. Saxifraga daqiaoensis (Saxifragaceae), a new species from Guangdong, China [J]. Ann Bot Fenn, 45(3): 237-239.
WEBB DA, GORNALL RJ, 1989. Saxifrages of Europe: with notes on African, American and some Asiatic species [M]. London: Helm.
WINKLER M, TRIBSCH A, SCHNEEWEISS GM, et al., 2012. Tales of the unexpected: Phylogeography of the arctic-alpine model plant Saxifraga oppositifolia (Saxifragaceae) revisited [J]. Mol Ecol, 21(18): 4618-30.
XIANG CL, GITZENDANNER MA, SOLTIS DE, et al., 2012. Phylogenetic placement of the enigmatic and critically endangered genus Saniculiphyllum ( Saxifragaceae ) inferred from combined analysis of plastid and nuclear DNA sequences [J]. Mol Phylogenet Evol, 64(2): 357-367.
XIANG QB, ZANG DK, 2014. International Code of Botanical Nomenclature (ICBN) [M]. Beijing: China Forestry Press: 1-206. [向其柏, 臧德奎, 2016. 國际栽培植物命名法规. 北京: 中国林业出版社: 1-206]
XU HJ, CHEN ZL, 2015. Application of Saxifraga stolonifera in landscape vertical greening [J]. Heilongjiang Agric Sci, (10): 126-128. [许红娟, 陈之林, 2015. 虎耳草在园林垂直绿化中应用初探 [J]. 黑龙江农业科学, (10): 126-128.]
YU SL, 2009. Potted maintenance technology of Saxifraga stolonifera, a health flower [J]. Chin Hortic Abstr, 25(11): 154. [于淑玲, 2009. 保健花卉——虎耳草的盆栽养护技术 [J]. 中国园艺文摘, 25(11): 154.]
ZHANG DJ, CHEN SY, GAO QB, et al., 2008. Circumscription and phylogeny of Saxifraga sect. Ciliatae: Evidence from nrDNA ITS sequences [J]. J Syst Evol, 46(5): 667-675. [张得钧, 陈生云, 高庆波, 等, 2008. 虎耳草属山羊臭组的界定和系统发育: 核糖体DNA ITS序列证据 [J]. 植物分类学报, 46(5): 667-675.]
ZHANG H, LI QY, HE SW, et al., 2021. Research progress on chemical constituents and pharmacological activities of Saxifraga stolonifera [J]. Prog Vet Med, 42(1): 94-99. [张慧, 李秋月, 贺尚文, 等, 2021. 虎耳草化学成分及药理活性研究进展 [J]. 动物医学进展, 42(1): 94-99.]
ZHANG JJ, ZHAO WY, MENG KK, et al., 2019. Saxifraga shennongii, a new species of Saxifragaceae from Hunan Province [J]. Phytotaxa, 418(1): 79-88.
ZHANG MH, 2018. Phylogenetic and biogeographic evolution of Saxifraga sect. Irregulares (Saxifragaceae) [D]. Jishou: Jishou University. [张梦华, 2018. 虎耳草属石荷叶组系统发育与地理演化研究 [D]. 吉首: 吉首大学: 1-70.]
ZHANG MH, WANG CY, ZHANG C, et al., 2020. Phylogenetic relationships and biogeographic history of the unique Saxifraga sect. Irregulares (Saxifragaceae) from eastern Asia [J]. J Syst Evol, 58(6): 958-971.
ZHANG MH, ZHANG XS, NIE ZL, et al., 2017. Saxifraga kegangii (Saxifragaceae), a new species from Hunan province of central China [J]. Phytotaxa, 309(2): 159-165.
ZHANG XJ, FU QS, CHEN JT, et al., 2021. Saxifraga viridiflora (Saxifragaceae), an unusual new species from Guangxi, China [J]. PhytoKeys, 184(184): 19-26.
ZHANG XJ, KUANG TH, CHEN JT, et al., 2022. Saxifraga sunhangiana (Saxifragaceae), a new species from Gansu, China [J]. Taiwania, 67(2): 195-200.
ZHANG XJ, LIU ZC, MENG KK, et al., 2018. Saxifraga luoxiaoensis (Saxifragaceae), a new species from Hunan and Jiangxi, China [J]. Phytotaxa, 350(3): 291-296.
ZHANG Z, 2016. Saxifraga stolonifera ‘Hei Kui of Chenshan won the most potential new cultivar [J]. For Sci Technol Commun, (9): 43. [張哲, 2016. 辰山‘黑魁虎耳草获评最具潜力新品种 [J]. 林业科技通讯, (9): 43.]
ZHANG ZX, XIA N, GORNALL RJ, 2015. Leaf venation patterns in the genus Saxifraga (Saxifragaceae) [J]. Phytotaxa, 221(2): 123-136.
ZHAO WY, MENG KK, FAN Q, et al., 2019. Saxifraga damingshanensis (S. sect. Irregulares, Saxifragaceae), a new species from Guangxi, China [J]. PhytoKeys, 133: 95-103.
ZHMYLEV PY, 2001. Pollination and sex forms in rockfoils in connection with genus Saxifraga L. (Saxifragaceae) evolution \[M\]. Moscow: Byulleten′ Moskovskogo Obshchestva Ispytatelei Prirody Otdel Biologicheskii, 106: 30-38.
ZUO GY, LI ZQ, CHEN LR, et al., 2007. Gallic acid esters of bergenin from Saxifraga melanocentra(Saxifragaceae)and their inhibition against HCV NS3 protease [J]. Acta Bot Yunnan, 29(4): 486-488. [左国营, 李正全, 陈丽蓉, 等, 2007. 黑蕊虎耳草中岩白菜素没食子酸酯类及其对丙型肝炎丝氨酸蛋白酶的抑制作用 [J]. 云南植物研究, 29(4): 486-488.]
( 责任编辑 李 莉 )
收稿日期: 2023-05-14
基金项目: 上海市绿化和市容管理局科研专项(G182412)。
第一作者: 唐世梅(1998-),硕士,研究方向为国产虎耳草属的多样性及观赏价值评价,(E-mail)1648943359@qq.com。
* 通信作者: 田代科,博士,研究员,博士生导师,研究方向为植物分类及多样性评价、植物迁地保育、观赏植物种质创新及利用等,(E-mail)dktian@cemps.ac.cn。
