两种玉凤花属植物的花结构和合蕊柱超微特征及其分类学意义
2024-03-10陶凯锋朱永王乐骋张颖铎李璐
陶凯锋 朱永 王乐骋 张颖铎 李璐


摘 要: 玉鳳花属植物的花形态变异多样,分类学问题复杂。为筛选出该属关键的分类学特征,该文以鹅毛玉凤花和粉叶玉凤花为研究对象,利用体式显微镜和扫描电镜,观察、比较了它们的花结构和合蕊柱超微特征。结果表明:(1)两种玉凤花的花结构差异显著,
尤其表现在侧花瓣裂片的有无、唇瓣3裂片形态、花距与子房的长度比。(2)合蕊柱超微特征在两个种间明显不同,表现在退化雄蕊、花药室外壁细胞、“花药室管”外壁细胞、“柱头枝”及其与蕊柱交界处的乳突等结构形态。(3)花粉小块外壁纹饰在种间不同,鹅毛玉凤花具网状花粉外壁纹饰,粉叶玉凤花则为棒状→网状的花粉外壁纹饰。该研究认为,侧花瓣裂片的有无、花距形态多变、花距与子房的长度比、花粉外壁纹饰类型等是玉凤花属物种鉴定的关键分类特征。在前人的研究基础上,该文补充并分析了粉叶玉凤花的花结构及合蕊柱的超微特征,不支持粉叶玉凤花独立于玉凤花属。
关键词: 兰科, 花结构, 合蕊柱, 柱头枝, 外壁纹饰, 花药室管
中图分类号: Q949.58
文献标识码: A
文章编号: 1000-3142(2024)01-0089-13
Floral structure and gynostemium ultrastructure of two
Habenaria species with their taxonomic implications
TAO Kaifeng, ZHU Yong, WANG Yuecheng, ZHANG Yingduo, LI Lu*
( College of Biodiversity Conservation, Southwest Forestry University, Kunming 650224, China )
Abstract: The genus Habenaria exhibits a highly diversity in floral morphology and has complex taxonomy problems. In order to select key taxonomic features of this genus, floral structure and gynostemium ultrastructure of H. dentata and H. glaucifolia were investigated using the stereo microscope and scanning electronic microscope (SEM). The results were as follows: (1) There were significant differences in floral features between species, especially in the presence or the absence of lateral lobes of petal, morphology of 3-lobed lip, and the length ratio of the spur to the ovary. (2) The ultrastructures of gynostemium were obviously different between the two species, which were shown in shapes of staminode, outer epidermal cells of anther wall, and stigma branch. (3) Exine sculptures of massula were distinguished in two species, the reticulate type in H. dentata, and the baculate → reticulate type in H. glaucifolia. It was suggested that some floral features were selected as potentially valuable for species delimitation, including the presence or absence of lateral petal lobes, variations in spur morphology, the length ratio of the spur to the ovary, and the types of pollen exine sculpture. Additional data on floral features and ultrastructures of gynostemium were reported as supplement for understanding of H. glaucifolia. Notably, it was not confirmed that H. glaucifolia should be treated as an independent genus from Habenaria.
Key words: Orchidaceae, floral structure, gynostemium, stigma branch, exine sculpture, anther sac channel
玉凤花属(Habenaria Willd.)是兰科(Orchidaceae)兰亚科(Orchidoideae)里最大的地生兰类群,约有876种,广泛分布于北半球温带地区以及新、旧世界的热带地区,非洲中部和南部及亚洲东部是该属的物种多样性中心(Dressler, 1993;Pridgeon et al., 2001)。中国有58种,包括22个特有种(金效华等,2019),集中分布于我国西南山区,特别是横断山脉地带(郎楷永等,1999)。玉凤花属为多年生草本植物,具发达的块茎;总状花序顶生,花型倒转,中萼片与两侧花瓣常靠合为兜状,花瓣2裂,花距明显或无;合蕊柱短,具2枚退化雄蕊;花药2室,有一对松散的花粉小块,药隔较宽,柱头凸起呈枝状,蕊喙发达且延伸成与花粉团柄靠合的“花药室管”(anther sac channel)(Dressler, 1993; 郎楷永等,1999; Pridgeon et al., 2001)。其中,玉凤花属的花瓣裂片和花距的有无,唇瓣裂片、药隔宽度、花粉小块、“花药室管”和枝状柱头等花形态特征常被用作该属及其近缘类群物种分类学依据(Dressler, 1981; Singer, 2001; Pedron et al., 2012; Suetsugu & Tanaka, 2014; Ikeuchi et al., 2015)。
然而,玉凤花属的地理分布广泛、物种多样、花结构复杂而变异丰富,其系统位置、属间界限及属内物种划分等争议较大,亟待解决(Dressler & Dodson, 1960; Bateman et al., 2003, 2009; Inda et al., 2012; Batista et al., 2013)。传统分类学上,玉凤花属被放置在玉凤花亚族(Habenariinae),该亚族与红门兰亚族(Orchidinae)共同构成红门兰族(Orchideae) (Dressler, 1993;Pridgeon et al., 2001)。这两个亚族的柱头结构差异显著,红门兰亚族为一个不分叉的凹陷柱头腔,而玉凤花亚族则延伸为2枚凸出的枝状柱头。不过,这样的分类学处理饱受争议,得不到其他花形态和分子证据的支持(Kurzweil & Weber, 1991, 1992; Pridgeon et al., 2001; Batista et al., 2013; Ngugi et al., 2020; Rewicz et al., 2022)。同時,枝状柱头这一分类学特征不仅出现在玉凤花属,还见于玉凤花亚族的其他类群,包括角盘兰属(Herminium Guett.)、阔蕊兰属(Peristylus Bl.)、手参属(Gymnadenia R. Br.)和兜被兰属(Neottianthe Schltr.)等。因此,很难用2枚枝状柱头这一特征来界定玉凤花属的分类界限(Kurzweil & Weber, 1992;Pridgeon et al., 2001)。目前,玉凤花属与其近缘类群之间的区别主要是根据2裂花瓣是否与其他花结构愈合、唇瓣是否深裂或缺乏胼胝体、柱头是否全缘等(Pridgeon et al., 2001; 郎楷永等,1999; Chen et al., 2009)。该属的部分物种曾被提升为独立的属,但后来都被处理为玉凤花属的异名(POWO, 2023),包括Podandriella Szlach.、Pseudoperistylus Szlach. & Olszewski、Centrostigma Schltr.、Platycoryne Rchb.f., Roeperocharis Rchb.f.等。因此,根据花形态特征,把它们处理为玉凤花属中形态较为独特的组(sectional rank)更合适(Kurzweil & Weber, 1992)。同时,曾被放在玉凤花属的一些类群被移出来独立为属,如舌唇兰属(Platanthera Rich.)和凹舌兰属(Coeloglossum Hartm.)(Dressler, 1993)。其中,椭圆叶玉凤花(Habenaria bakeriana)因其不形成“花药室管”且唇瓣舌状无裂片等特征而从玉凤花属移到舌唇兰属,并得到分子系统学的强烈支持(Bateman et al., 2009),被更名为滇藏舌唇兰(Platanthera bakeriana),被认为与玉凤花属关系密切(郎楷永,1994)。同样,根据花形态特征和合蕊柱器官发生模式,紫斑玉凤花(Habenaria purpureopunctata)曾被提升为紫斑兰属(Hemipiliopsis Y. B. Luo & S. C. Chen)(Luo & Chen, 2003; Luo et al., 2005),但后来的分子系统学支持把该种放置于舌喙兰属(Hemipilia Lindl.),将其修订为紫斑兰(H. purpureopunctata)(Jin et al., 2014)。现有分子系统学研究结果表明,玉凤花属与红门兰亚族的大部分属一样,并非单系,可以分成两个距离较远的分支,其他近缘属构成一支,成为红门兰亚族的姐妹群,关于该属的分类系统尚需更多证据来澄清(Batista et al., 2013; Jin et al., 2014; Ngugi et al., 2020; Rewicz et al., 2022)。
兰科的花结构形态及其超微特征在亚科、族和属间界限划分方面具有重要作用(Dressler, 1981, 1986; Freudenstein & Rasmussen, 1997, 1999; Bateman & Rudall, 2006; Singer et al., 2007),包括花瓣、唇瓣、合蕊柱、花药、花粉、花药帽和柱头等,见于拟兰亚科和杓兰亚科(Newton & Willams, 1978; Burns-balogh & Hesse, 1988),红门兰亚族(Kurzweil & Weber, 1991; Luo & Chen, 2003; Luo et al., 2005; Lumaga et al., 2006; Bell et al., 2009; Passarelli & Rolleri, 2010),石豆兰属(Bulbophyllum Thouars)(Nunes et al., 2014, 2017)和石斛属(Dendrobium Sw.)(王艳萍等,2021a, b)。为探究玉凤花亚族的花形态结构及其超微特征的分类学意义,早期的研究观察了非洲分布的红门兰亚族和玉凤花亚族下的属间花形态结构特征和合蕊柱的超微特征,认为花药直立和枝状柱头等特征在玉凤花属及近缘属间存在重叠,难以用于区分两个亚族(Kurzweil & Weber, 1991, 1992)。为探究花粉形态特征的分类学意义,有研究利用扫描电镜观察了南美产的4种玉凤花(Passarelli & Rolleri, 2010)和另两种玉凤花(Liu, 2015, 2016)的花粉小块超微特征,结果表明花粉小块形态及其外壁纹饰、四合花粉排列及外壁黏性丝状物等形态特征在种间差异显著,值得深入研究。
粉叶玉凤花(Habenaria glaucifolia)为中国特有的一种地生兰,仅分布于陕西、甘肃、四川、贵州、云南及西藏,生于海拔2 000~4 300 m的山坡林下、灌丛下或草地上(郎楷永等,1999)。其花形态特征较为独特,唇瓣3裂呈线条状并反卷,药隔较宽为马蹄形或U型,两枚枝状柱头较长,呈长披针形。据此,该种曾被处理为独立的属,即粉叶玉凤花属(Senghasiella Szlach.)(Szlachetko, 2001),但根据合蕊柱器官发育特征,粉叶玉凤花依旧被处理为玉凤花属的成员,其独特的花形态特征是该属系统演化方面的衍生类型(Luo et al., 2005)。鹅毛玉凤花(Habenaria dentata)是广布于亚洲东南部的热带和亚热带地区的一种地生兰,生于海拔190~2 300 m的山坡林下或沟边,在我国见于南方诸省(郎楷永等,1999)。其块茎富含植物化学活性成分,为我国传统中药材(陈谦海,2004)。该种曾被置于红门兰属(Orchis L.)(Grefing et al., 1800)和舌唇兰属中(Lindley, 1835),后又被重新归到玉凤花属(Govaerts, 2003)。考虑到亚洲分布的玉凤花属的花结构及其超微特征的分类学方面研究资料较少(Luo et al., 2005; Liu, 2015, 2016),本研究以中国产的鹅毛玉凤花和粉叶玉凤花为研究对象,利用显微镜和扫描电镜,比较分析它们的花结构、合蕊柱及花粉小块超微特征的差异,拟探讨以下问题:(1)通过观察两种玉凤花的花结构特征,分析该属关键的分类学特征;(2)玉凤花属的合蕊柱及花粉小块超微特征的分类学意义。
1 材料与方法
两种植物的实验材料均采自中国云南省,鹅毛玉凤花采自景东县无量山,粉叶玉凤花采自丽江市玉龙雪山,采集于2020—2021年。取部分新鲜材料在解剖镜(Leica M165 FC)下观察成熟花的形态解剖特征并拍照。其余新鲜材料固定于50%的乙醇,24 h后转到70%的乙醇常温保存。固定后的材料在解剖镜下分离出花药和柱头,经乙醇梯度脱水,后用液态二氧化碳进行临界点干燥。干燥后用解剖针刺破花药壁,以释放花粉。在体式镜下,将材料置于贴有导电胶的样品台上,随后将样品台放入离子溅射仪中喷镀铂金,最后置于扫描电镜(ZEISS Sigma 300)下观察并拍照。通过图像编辑软件Adobe Photoshop CC 2019进行图版制作。
兰科植物形态术语参考《中国兰科植物鉴别手册》(陈心启等,2009)。花粉超微特征描述参考《种子植物花粉电镜图志》(韦仲新,2003)。
2 结果与分析
2.1 两种玉凤花属植物的花结构特征比较
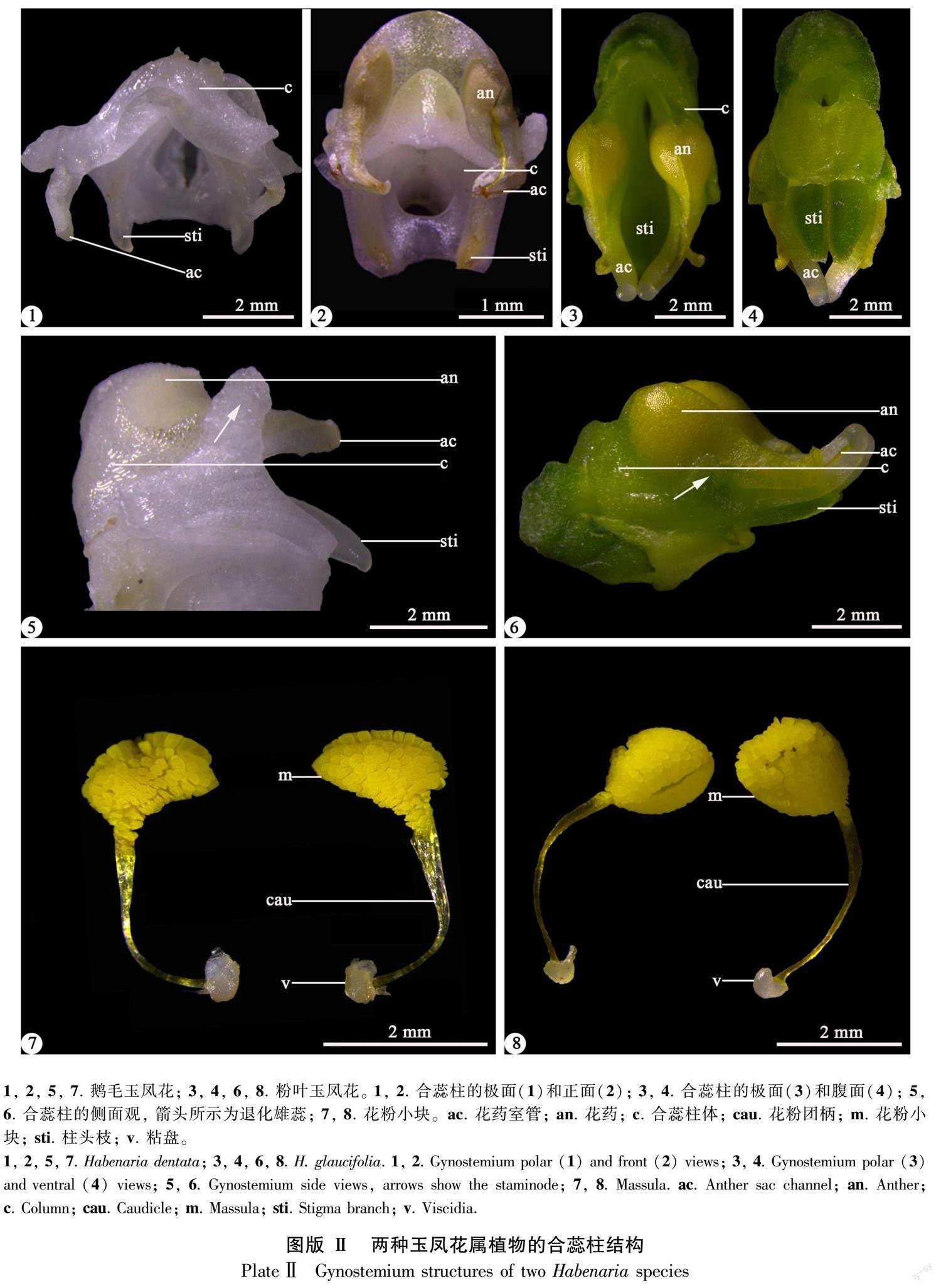
解剖镜下,鹅毛玉凤花的花序长5~12 cm,无毛,花白色(图版Ⅰ :1)。中萼片与花瓣靠合成兜状,侧萼片反折(图版Ⅰ :3)。侧花瓣不裂,呈披针形(图版Ⅰ :7)。唇瓣3浅裂,中裂片舌状披针形,侧裂片近菱形,边缘具锯齿(图版Ⅰ :5)。其花距明显长于子房(图版Ⅰ :9)。花药直立,两侧各具1枚退化雄蕊(图版Ⅱ:5)。花药室2,每个药室内各具1个粒粉质的花粉团,每个花粉团具1个狭长的花粉团柄和1个裸露的粘盘(图版Ⅱ:7)。药室基部延伸出包裹花粉团柄的狭窄结构——“花药室管”(图版Ⅱ:1,2,5)。该种具2个柱头且伸长成一对分离的“柱头枝”,位于合蕊柱的基部(图版Ⅱ:1,2)。
粉叶玉凤花的花序长5~20 cm,被短柔毛,花绿白色(图版Ⅰ :2)。中萼片也与花瓣靠合成兜状,侧萼片反折(图版Ⅰ :4)。侧花瓣2深裂,上裂片近等长于中萼片,匙状长圆形;下裂片披针形,远小于上裂片(图版Ⅰ :8)。唇瓣基部之上3深裂,中裂片线形,稍宽于侧裂片,侧裂片前部拳卷状(图版Ⅰ :6)。其花距与子房近等长(图版Ⅰ :10)。花药直立,两侧各具1枚退化雄蕊(图版Ⅱ :6)。药室2,每个药室内各具1个粒粉质的花粉团,每个花粉团具1个狭长的花粉团柄和1个裸露的粘盘(图版Ⅱ :8)。药室基部也延伸出包裹花粉团柄的“花药室管”。粉叶玉凤花也具2个柱头,位于蕊柱基部,但其所延伸出的“柱头枝”靠合在一起(图版Ⅱ :3,4)。
2.2 两种玉凤花属植物的合蕊柱超微特征比較
扫描电镜下,两种玉凤花的花药超微特征较为相似,表现在以下特征:花药室外侧均有一枚明显的耳状退化雄蕊;花药室外壁细胞排列紧密,并且相邻细胞间的界限较为明显;药室基部延伸成“花药室管”,包裹着花粉团柄,其外壁细胞均呈四边形,轮廓清楚、排列整齐(图版Ⅲ :5,6)。但是,退化雄蕊、花药室及“花药室管”外壁细胞形态具显著差异。第一,鹅毛玉凤花的退化雄蕊呈棒状(图版Ⅲ :1),而粉叶玉凤花的退化雄蕊则近三角形(图版Ⅲ :2)。第二,鹅毛玉凤花的花药室外壁细胞较粉叶玉凤花规则,前者表面具致密的波状条纹,而后者则表面具突起且相对光滑(图版Ⅲ :3,4)。第三,鹅毛玉凤花的“花药室管”外壁细胞表面具皱波状纹饰,而粉叶玉凤花的“花药室管”外壁细胞表面较光滑(图版Ⅲ :7,8)。
此外,两种玉凤花的柱头都延伸成“柱头枝”,在鹅毛玉凤花的柱头枝上观察到了可授性区域,位于柱头边缘增厚且光滑的区域(图版Ⅳ :1,3)。二者柱头与蕊柱交界处均具乳突(图版Ⅳ :5,6)。但是,两种玉凤花的乳突形态和大小存在差异,鹅毛玉凤花的乳突小于粉叶玉凤花,并且表面具排列紧密的波状条纹(图版Ⅳ :5,7)。粉叶玉凤花的乳突表面也具条纹,但较前者粗,呈脊状(图版Ⅳ :6,8)。
2.3 两种玉凤花属植物的花粉小块超微特征比较
两种玉凤花的花粉小块都近四棱形或扁平楔形(图版Ⅴ :1,2),花粉小块表面均具覆盖层或残存的覆盖层及部分丝状物(图版Ⅴ :3,4)。鹅毛玉凤花的每个花药室中央有条隔膜组织,在隔膜组织两侧有一层直立且排列紧密的花粉小块(图版Ⅴ :1)。在该种花粉小块的横切面上,还可见二细胞型花粉中的生殖细胞和营养细胞(图版Ⅴ :7)。粉叶玉凤花的花粉四分体排列方式较鹅毛玉凤花明显,多为正四面体形和十字交叉形。其中,还可见T形排列的四分体,但占比较少(图版Ⅴ :2)。
二者主要的差异在于花粉外壁纹饰类型。鹅毛玉凤花的花粉外壁纹饰呈不规则的网状,网脊明显,网眼形状不规则,网眼中具颗粒状雕纹(图版Ⅴ :5)。粉叶玉凤花具两种类型的花粉外壁纹饰:花粉小块两端的花粉外壁纹饰呈网状,网脊高,网眼大小不等,网眼中具颗粒雕纹;中央的花粉外壁纹饰则为短棒状(图版Ⅴ :6,8)。
3 讨论与结论
3.1 两种玉凤花属植物的花结构特征及分类学意义
侧花瓣、唇瓣、花距等花结构特征在玉凤花属的物种间差异明显,是编制该属物种检索表的关键特征(郎楷永等,1999;Pridgeion et al., 2001;Chen et al., 2009)。本研究的两种玉凤花具1对花粉块、较长的花粉团柄、裸露的粘盘及2个柱头等该属典型特征,但二者的唇瓣、侧花瓣及花距等结构差异显著。
第一, 侧花瓣开裂与否是该属的一个重要分类特征。玉凤花属大部分物种的花瓣具2裂且位于前部的裂片经常与蕊喙侧裂片及唇瓣愈合(Kurzweil & Weber, 1992)。根据侧花瓣裂片的有无,可分为2种,即侧花瓣全缘和侧花瓣2裂。侧花瓣全缘的类型出现在厚瓣玉凤花(Habenaria delavayi)和小巧玉凤花(H. diplonema)等种中,侧花瓣2裂见于落地金钱(H. aitchisonii)和卧龙玉凤花(H. wolongensis)等(郎楷永等,1999; Chen et al., 2009)。本研究中,鹅毛玉凤花的侧花瓣全缘,而粉叶玉凤花的侧花瓣2裂。可见,侧花瓣裂片的有无具有种间分类学意义。
第二,唇瓣裂片也可作为玉凤花属的物种鉴定依据。该属植物的唇瓣形状存在显著的差异,但3裂的唇瓣及丝状或披针状的深裂是最常见的类型,仅少数该属及其近缘的Platycoryne植物唇瓣全缘,4裂或5裂的唇瓣则更罕见,只见于与玉凤花属近缘的狗兰属(Cynorkis Thouars)中(Pridgeion et al., 2001; Chen et al., 2009)。岩生玉凤花(Habenaria idua)的植株和花部特征与丛叶玉凤花(H. tonkinensis)较为相似,但前者因唇瓣绿色且唇瓣侧裂片明显上举而区别于后者(张文柳等,2015)。本文中两种玉凤花的唇瓣均3裂,但唇瓣的裂片形态存在显著差异。鹅毛玉凤花的唇瓣中裂片舌状披针形,侧裂片近菱形,边缘具锯齿。粉叶玉凤花的唇瓣中裂片线形,侧裂片前部拳卷状。因此,唇瓣特征的分类学意义值得重视。
第三,玉凤花属的花距特征也呈现出多样性。在大多数玉凤花中,唇瓣基部与合蕊柱愈合且二者愈合的部位通常形成花距(Kurzweil & Weber, 1992)。该属的花距通常呈细长的丝状,一般较长,顶端渐细或膨大成棒状。后一性状在该属部分物种中演化到极致,如H.cataphysema的花距末端形成一个巨大的球形结构(Williamson, 1977)。在该属大多数物种中,花距直或略弯曲,但也有许多物种的花距呈不同程度的膝状弯曲(如H. tentaculigera)或螺旋状扭曲;而H. anaphysema的花距则更为特殊,其形成一个环(Kurzweil & Weber, 1992)。本文观察到两种玉凤花属植物均具近棒状的花距,但存在显著差异。鹅毛玉凤花的花距末端膨大,与距口近等宽;而粉叶玉凤花的花距中部弯曲且花距末端要窄于距口,不膨大。此外,花距与子房的长度比在该属不同的物种中区别显著。根据花距与子房的长度比,可把该属物种分为3种:花距明显长于子房;花距与子房近等长;花距短于子房。花距明显长于子房的类型出现在斧萼玉凤花(H. commelinifolia)和长距玉凤花(H. davidii)中,花距与子房近等长的类型见于卧龙玉凤花(H. wolongensis)和十字兰(H. schindleri),花距短于子房的类型出现在中泰玉凤花(H. siamensis)和剑叶玉凤花(H. pectinata)中(郎楷永等,1999; Chen et al., 2009)。在本研究中,鹅毛玉凤花的花距較子房长,而粉叶玉凤花的花距则与子房近等长。因此,花距特征也是该属重要的分类依据。
3.2 两种玉凤花属植物的合蕊柱超微特征及分类学意义
相较于具有两或三枚可育雄蕊并缺少蕊喙的拟兰亚科和杓兰亚科,兰亚科无疑是更为进化的类群,其有一枚可育雄蕊和两枚退化雄蕊,蕊喙也没有缺失(Dressler & Dodson, 1960)。
第一,根据解剖镜及扫描电镜观察结果,本研究的两种玉凤花属植物均只有一枚由可育雄蕊发育而来的花药,花药室外侧各具一枚退化雄蕊,药室基部延伸成包裹花粉团柄的“花药室管”,属于较进化的类群。但是,两种玉凤花的退化雄蕊形状及“花药室管”外壁细胞存在差异,鹅毛玉凤花具棒状退化雄蕊及皱波状“花药室管”外壁细胞,而粉叶玉凤花的退化雄蕊近三角形且“花药室管”外壁细胞表面较光滑。退化雄蕊形状及“花药室管”外壁细胞特征有待进一步研究。
第二,两种玉凤花属植物的两个柱头都伸长成“柱头枝”且具乳突和可授性区域。在兰科中,仅兰亚科红门兰亚族下的玉凤花属、舌唇兰属、角盘兰属(Herminium L.)等属具两个柱头,但玉凤花属的柱头延长成“柱头枝”,相对其他类群较明显(Dressler & Dodson, 1960; Dressler, 1993; Chase et al., 2015)。该属合蕊柱通常无毛,但也有例外,如Habenaria hirsutitrunci的柱头突起就密布毛(Williamson, 1980)。本研究中,鹅毛玉凤花的柱头枝离生,而粉叶玉凤花的柱头枝则联合成舌状,与罗毅波等通过扫描电镜所观察到的粉叶玉凤花的柱头枝相一致(Luo et al., 2005)。在上述两种玉凤花的合蕊柱上均未观察到明显的毛状结构,并且二者柱头枝上的乳突大小及表面纹饰均不同。
第三,本研究还在鹅毛玉凤花的柱头边缘增厚的部位发现了可授性区域,与前人在粉叶玉凤花及二叶舌唇兰(Platanthera chlorantha)上所观察到的可授性区域的位置相一致(Stpiczyn, 2003;Luo et al., 2005)。已有资料表明,可授性区域可为柱头枝上延长的扁平裂片,如Habenaria armatissima和H. mira(Kurzweil & Weber, 1992)。因此,这两种玉凤花的合蕊柱特征存在显著差异,并且合蕊柱特征对于该属的分类鉴定具有重要意义,值得深究。
3.3 两种玉凤花属植物的花粉外壁纹饰及分类学意义
由于授粉方式不同,植物会产生与功能相适应的花粉纹饰,风媒花粉表面一般光滑,虫媒花粉表面粗糙而有纹饰。玉凤花属植物为典型的虫媒花,其花结构高度特化,花粉外壁纹饰亦较复杂(Singer, 2001; Suetsugu & Tanaka, 2014; Xiong et al., 2015)。
结合现有文献资料,在分子系统学一致支持把兰科分为5个亚科的分类系统中(Chase et al., 2015),拟兰亚科的剑叶拟兰(Apostasia wallichii)和三蕊兰(Neuwiedia zollingeri var. singapureana)的花粉具网状外壁纹饰(Newton & Williams, 1978)。大多数杓兰亚科植物缺乏在其他亚科中发现的复杂外壁纹饰,杓兰属和兜兰属大多物种的花粉外壁光滑无任何纹饰(Williams & Broome, 1976)。树兰亚科的头蕊兰属(Cephalanthera Rich.)具3种纹饰,即光滑无任何纹饰、粗糙状纹饰和棒状纹饰;丛宝兰属(Limodorum L.)具粗糙状-棒状纹饰;无叶兰属(Aphyllorchis Blume)和火烧兰属(Epipactis Zinn)大多具网状纹饰(Ackerman & Williams, 1980)。兰亚科中各属的花粉外壁纹饰类型较为丰富,其中,倒距兰属(Anacamptis Rich.)具有2种花粉外壁纹饰,即光滑无任何纹饰-粗糙状纹饰和粗糙状-皱波状纹饰;掌裂兰属(Dactylorhiza Neck. ex Nevski)具3种花粉外壁纹饰,即光滑无任何纹饰、光滑无任何纹饰-粗糙状纹饰和网状纹饰;红门兰属具4种花粉外壁纹饰,即光滑无任何纹饰、网状纹饰、穿孔-皱波状纹饰和棒状纹饰(Lumaga et al., 2006)。
然而,玉凤花属植物的花粉外壁纹饰则更为多样,有光滑无任何纹饰、粗糙状、皱波状、棒状 [十字兰(Habenaria schindleri)]、疣状、网状等类型(Schill & Pfeiffer, 1977; Liu, 2015, 2016),其中还不乏各类型纹饰中的过渡类型(Hesse & Burns-Balogh, 1984; Lumaga et al., 2006)。现有研究表明,红门兰亚族植物的花粉外壁纹饰呈现出两种演化趋势:光滑无任何纹饰→皱波状或粗糙状纹饰 [由倒距兰屬、斑鸭兰属(Neotinea Rchb.f.)、蜂兰属(Ophrys L.)和长药兰属(Serapias L.)组成的分支]和光滑无任何纹饰→棒状纹饰→网状纹饰 [由矮麝兰属(Chamorchis Rich.)、掌裂兰属、手参属、红门兰属、舌唇兰属等所组成的分支](Lumaga et al., 2006)。本研究中,鹅毛玉凤花具网状花粉外壁纹饰,粉叶玉凤花的花粉外壁纹饰则为棒状→网状的过渡类型。根据红门兰亚族的花粉外壁纹饰演化趋势,鹅毛玉凤花的花粉外壁类型较粉叶玉凤花进化。
综上所述,玉凤花属侧花瓣裂片的有无、唇瓣3裂片及花距的形态、花距与子房的长度比、花粉外壁纹饰类型等具种级水平的可见差异,因而可作为属内物种划分的依据,应重视其在分类学上的应用。此外,本研究观察到鹅毛玉凤花和粉叶玉凤花的部分花结构及合蕊柱超微特征较为相似,如相互靠合的中萼片和侧花瓣、突起的柱头、柱头上的可授性区域位置、退化雄蕊的位置、粒粉质的花粉团、花粉外壁纹饰等,并且上述特征均符合玉凤花属的分类性状,所以不支持将粉叶玉凤花独立成属(Szlachetko, 2001; Luo et al., 2005)。
致谢 西南林业大学云南生物多样性研究院提供实验平台,古旭老师协助部分实验;中国科学院昆明植物研究所东亚植物与生物地理学重点实验室植物多维成像及多样性分析平台提供扫描电镜服务,谷志佳老师协助电镜扫描观察。在此一并谨致谢意。
参考文献:
AC KERMAN JD, WILLIAMS NH, 1980. Pollen morphology of the tribe Neottieae and its impact on the classification of the Orchidaceae [J]. Grana, 19(1): 7-18.
BATEMAN RM, HOLLINGSWORTH PM, PRESTON J, et al., 2003. Molecular phylogenetics and evolution of Orchidinae and selected Habenariinae (Orchidaceae) [J]. Bot J Linn Soc, 142(1): 1-40.
BATEMAN RM, RUDALL PJ, 2006. Evolutionary and morphometric implications of morphological variation among flowers within an inflorescence: a case-study using European orchids [J]. Ann Bot, 98(5): 975-993.
BATEMAN RM, JAMES KE, LUO YB, et al., 2009. Molecular phylogenetics and morphological reappraisal of the Platanthera clade (Orchidaceae: Orchidinae) prompts expansion of the generic limits of Galearis and Platanthera [J]. Ann Bot, 104(3): 431-445.
BATISTA JAN, BORGES KS, DE FARIA MWF, et al., 2013. Molecular phylogenetics of the species-rich genus Habenaria (Orchidaceae) in the New World based on nuclear and plastid DNA sequences [J]. Mol Phylogenet Evol, 67(1): 95-109.
BALOGH P, 1982. Generic redefinition in subtribe Spiranthinae (Orchidaceae) [J]. Amer J Bot, 69(7): 1119-1132.
BELL AK, ROBERTS DL, HAWKINS JA, et al., 2009. Comparative micromorphology of nectariferous and nectarless labellar spurs in selected clades of subtribe Orchidinae (Orchidaceae) [J]. Bot J Linn Soc, 160(4): 369-387.
BURNS-BALOGH P, HESSE M, 1988. Pollen morphology of the cypripedioid orchids [J]. Plant Syst Evol, 158(2): 165-182.
CHASE MW, CAMERON KM, et al., 2015. An updated classification of Orchidaceae [J]. Bot J Linn Soc, 177(2): 151-174.
CHEN SC,CRIBB, PJ, 2009. Habenaria Willdenow [M]//WU ZY, RAVEN PH, HONG DY, et al. Flora of China, Vol. 25. Beijing: Science Press; St. Louis: Missouri Botanical Garden Press: 144-160.
CHEN QH, 2004. Flora of Guizhou: Vol. 10 [M]. Guiyang: Guizhou Science and Technology Press: 389. [陳谦海, 2004. 贵州植物志: 第10卷 [M]. 贵阳: 贵州科技出版社: 389. ]
DRESSLER RL, DODSON CH, 1960. Classification and phylogeny in the Orchidacea [J]. Ann Mo Bot Gard, 47(1): 25-68.
DRESSLER RL, 1981. The orchids: natural history and classification [M]. Cambridge: Harvard University Press.
DRESSLER RL, 1986. Recent advances in orchid phylogeny [J]. Lindleyana, 1(1): 5-20.
DRESSLER RL, 1993. Phylogeny and classification of the orchid family [M]. Cambridge: Cambridge University Press.
ERDTMAN G, 1960. Pollen walls and angiosperm phylogeny [J]. Bot Notis, 113: 41-45.
FREUDENSTEIN JV, RASMUSSEN FN, 1997. Sectile pollinia and relationships in the Orchidaceae [J]. Plant Syst Evol, 205(3): 125-146.
FREUDENSTEIN JV, RASMUSSEN FN, 1999. What does morphology tell us about orchid relationships?—A cladistic analysis [J]. Amer J Bot, 86(2): 225-248.
GOVAERTS R, 2003. World checklist of monocotyledons database in ACCESS: 1-71827 [M]. Kew: The Board of Trustees of the Royal Botanic Gardens.
GREFING LL, HORR JL, LANGE JG, et al., 1800. Kungliga. Svenska vetenskapsakademiens handlingar [M]. Stockholm: L. L. Grefing.
HESSE M, BURNS-BALOGH P, 1984. Pollen and pollinarium morphology of Habenaria (Orchidaceae) [J]. Pollen et Spores, 26(3-4): 385-400.
IKEUCHI Y, SUETSUGU K, SUMIKAWA H, 2015. Diurnal skipper Pelopidas mathias (Lepidoptera: Hesperiidae) pollinates Habenaria radiata (Orchidaceae) [J]. Entomol News, 125(1): 7-11.
INDA LA, PIMENTEL M, CHASE MW, 2012. Phylogenetics of tribe Orchideae (Orchidaceae: Orchidoideae) based on combined DNA matrices: inferences regarding timing of diversification and evolution of pollination syndromes [J]. Ann Bot, 110(1): 71-90.
JIN WT, JIN XH, SCHUITEMAN A, et al., 2014. Molecular systematics of subtribe Orchidinae and Asian taxa of Habenariinae (Orchideae, Orchidaceae) based on plastid matK, rbcL and nuclear ITS [J]. Mol Phylogenet Evol, 77: 41-53.
JIN XH, LI JW, YE DP, 2019. Atlas of native Orchids in China [M]. Zhengzhou: Henan Science and Technology Press: 308-344. [金效華, 李剑武, 叶德平, 2019. 中国野生兰科植物原色图鉴 [M]. 郑州: 河南科学技术出版社: 308-344. ]
KRANZLIN F, 1897. Orchidacearum genera et species [M]. Berlin: Mayer & Miiller.
KURZWEIL H, WEBER A, 1991. Floral morphology of southern African Orchideae. I. Orchidinae [J]. Nord J Bot, 11(2): 155-178.
KURZWEIL H, WEBER A, 1992. Floral morphology of southern African Orchideae. II. Habenariinae [J]. Nord J Bot, 12(1): 39-61.
LANG KY, 1994. Studies on the distribution patterns of some significant genera in orchid flora [J]. J Syst Evol, 32(4): 328-339. [郎楷永, 1994. 兰科植物区系中一些有意义属的地理分布格局的研究 [J]. 植物分类学报, 32(4): 328-339.]
LANG KY, CHEN XQ, LUO YB,et al., 1999. Flora Reipublicae Popularis Sinicae: Vol. 17 [M]. Beijing: Science Press. [郎楷永, 陈心启, 罗毅波, 等, 1999. 中国植物志: 第17卷 [M]. 北京: 科学出版社. ]
LINDLEY J, 1835. The genera and species of orchidaceous plants [M]. London: Ridgways.
LIU L, 2015. Ultramicroscopic examination of mature massulae of Habenaria arinaria (Orchidaceae) [J]. Micron, 74: 1-7.
LIU L, 2016. Ultramicroscopic Characterization of Mature Pollen Grains of Habenaria sagittifera [J]. Hortic Plant J, 2(1): 50-54.
LUMAGA MRB, COZZOLINO S, KOCYAN A, 2006. Exine micromorphology of Orchidinae (Orchidoideae, Orchidaceae): phylogenetic constraints or ecological influences? [J]. Ann Bot, 98(1): 237-244.
LUO YB, CHEN SC, 2003. Hemipiliopsis, a new genus of Orchidaceae [J]. Novon, 13(4): 450-453.
LUO YB, ZHU GH, KURZWEIL H, 2005. The gynostemium of Hemipiliopsis purpureopunctata and Senghasiella glaucifolia, two taxonomically disputed species of Habenariinae (Orchidaceae) [J]. Bot J Linn Soc, 147(2): 191-196.
NEWTON GD, WILLIAMS NH, 1978. Pollen morphology of the Cypripedioideae and the Apostasioideae (Orchidaceae) [J]. Selbyana, 2(2/3): 169-182.
NGUGI G, LE PT, MARTOS F, et al, 2020. Phylogenetic relationships amongst the African genera of subtribe Orchidinae s. l. (Orchidaceae; Orchideae): implications for subtribal and generic delimitations [J]. Mol Phylogenet Evol, 153: 106946.
NUNES ELP, SMIDT EC, STTZEL T, et al., 2014. What do floral anatomy and micromorphology tell us about Neotropical Bulbophyllum section Didactyle (Orchidaceae: Bulbophyllinae)? [J]. Bot J Linn Soc, 175(3): 438-452.
NUNES ELP, MALDONADO PE, SMIDT EC, et al., 2017. Floral micromorphology and anatomy and its systematic application to Neotropical Bulbophyllum section Micranthae (Orchidaceae) [J]. Bot J Linn Soc, 183(2): 294-315.
PASSARELLI LM, ROLLERI CH, 2010. Pollen grains and massulae in pollinia of four south american palustrine species of Habenaria (Orchidaceae) [J]. Grana, 49(1): 47-55.
PEDRON M, BUZATTO CR, SINGER RB,et al., 2012. Pollination biology of four sympatric species of Habenaria (Orchidaceae: Orchidinae) from southern Brazil [J]. Bot J Linn Soc, 170(2): 141-156.
POWO, 2023. Plants of the world online [EB/OL]. Facilitated by Kew: the Royal Botanic Gardens. http: //www.plantsoftheworldonline.org//.
PRIDGEON AM, CRIBB PJ, CHASE MW, et al., 2001. Genera Orchidacearum: Vol. 2. Orchidoideae (Part one) [M]. New York: Oxford University Press: 1-416.
REWICZ A, KOLANOWSKA M, KRAS M, et al., 2022. Preliminary studies on variation in the micromorphology of the seed coat in Habenaria s. l. (Orchidaceae) and relatives [J]. Bot J Linn Soc, 200(1): 104-115.
SCHILL R, PFEIFFER W, 1977. Untersuchungen an Orchideenpollinien unterbesonderer Beruücksichtigung ihrer Feinskulpturen [J]. Pollen et Spores, 19: 5-118.
SINGER RB, 2001. Pollination biology of Hebenaria parviflora (Orchidaceae: Habenariinae) in southeastern Brazil [J]. Darwiniana, 39(3-4): 201-207.
SINGER RB, BREIER TB, FLACH A, et al., 2007. The Pollination Mechanism of Habenaria pleiophylla Hoehne & Schlechter (Orchidaceae: Orchidinae) [J]. Funct Ecosyst Comm, 1(1): 10-14.
STPICZYN M, 2003. Stigma receptivity during the life span of Platanthera chlorantha Custer (Rchb.) flowers [J]. Acta Biol Cracov Ser Bot, 45(1): 37-41.
SUETSUGU K, TANAKA K, 2014. Consumption of Habenaria sagittifera pollinia by juveniles of the katydid Ducetia japonica [J]. Entomol Sci, 17(1): 122-124.
SUMMERHAYES VS, 1960. African orchids: XXVII [J]. Kew Bull, 14(1): 126-159.
SZLACHETKO DL, 2001. Senghasiella, eine neue Gattung aus der Habenaria-Verwandtschaft [J]. J Orchid Freund, 8: 365-368.
SZLACHETKO DL, OLSZEWSKI TS, 1998. Orchidées 1[M]//SABATIE B, MORAT P. Flore du Cameroun. Yaoundé: Minrest: 102-236.
WANG YP, LI CR, LUO Y, et al., 2021a. Preliminary study on morphology and taxonomic significance of anther cap in nine Dendrobium species (Orchidaceae) from China [J]. Plant Sci J, 39(4): 367-378. [王艳萍, 李楚然, 罗艳, 等, 2021a. 中国9种石斛属植物的花药帽形态及其分类学意义初探 [J]. 植物科学学报, 39(4): 367-378.]
WANG YP, LI L, YANG CX, et al., 2021b. Pollinia morphology of 14 species in Dendrobium (Orchidaceae) and taxonomic significance [J]. Bull Bot Res, 41(1): 12-25. [王艷萍, 李璐, 杨晨璇, 等, 2021b. 14种石斛属(兰科)植物的花粉团形态及分类学意义 [J]. 植物研究, 41(1): 12-25.]
WEI ZX, 2003. Pollen flora of seed plants (all photos from scanning electron microscope) [M]. Kunming: Yunnan Science and Technology Press: 1-7. [韦仲新, 2003. 种子植物花粉电镜图志 [M]. 昆明: 云南科技出版社: 1-7. ]
WILLIAMS NH, BROOME CR, 1976. Scanning electron microscope studies of orchid pollen [J]. Bull Am Ceram Soc, 45: 699-707.
WILLIAMSON G, 1977. The orchids of south central Africa [M]. London: J. M. Dent & Sons, Ltd.
WILLIAMSON G, 1980. Contribution to the orchid flora of South Central Africa [J]. Plant Syst Evol, 134(1/2): 53-77.
XIONG YZ, LIU CQ, HUANG SQ, 2015. Mast fruiting in a hawkmoth-pollinated orchid Habenaria glaucifolia: an 8-year survey [J]. J Plant Ecol, 8(2): 136-141.
ZHANG WL, GAO JY, LIU Q, 2015. Habenaria vidua, a new recorded of Orchidaceae from Yunnan, China [J]. Guihaia, 35(1): 75-76. [张文柳, 高江云, 刘强, 2015. 中国兰科玉凤花属一新记录种——岩生玉凤花 [J]. 广西植物, 35(1): 75-76.]
( 责任编辑 周翠鸣 )
收稿日期: 2023-03-17
基金项目: 国家自然科学基金(32060049) 。
第一作者: 陶凯锋(1999-),硕士研究生,研究方向为资源利用与植物保护,(E-mail)17856109959@ 163.com。
* 通信作者: 李璐,博士,副研究员,研究方向为植物系统与进化、植物形态解剖学,(E-mail)lilusaraca@ foxmail.com, lilu@ swfu.edu.cn。
