不同温度下PEG模拟干旱对濒危植物狭叶坡垒种子萌发的影响
2024-03-10梁惠子杨继生覃毅潘韦虎肖玉菲黄荣林王仁杰蒋燚刘雄盛
梁惠子 杨继生 覃毅 潘韦虎 肖玉菲 黄荣林 王仁杰 蒋燚 刘雄盛

摘 要: 狹叶坡垒(Hopea chinensis)种子为典型的顽拗性种子。为探究种子萌发对温度和水分的适应性,该研究在人工气候培养箱内设置3种温度(15、20、25 ℃),采用6个聚乙二醇(PEG-6000)浓度(0%、5%、10%、15%、25%和35%)模拟干旱胁迫处理,研究狭叶坡垒种子的萌发特性。结果表明:(1)温度对种子萌发具有显著影响。同一干旱胁迫浓度下,种子的萌发率、萌发势、萌发指数、胚根长、芽长和活力指数随着温度升高呈上升趋势,萌发时滞随温度升高而降低,萌发历期随温度升高而波动性上升。(2)干旱胁迫对种子萌发具有显著影响。同一温度下,种子的萌发率、萌发势、萌发指数、萌发历期、胚根长、胚芽长和活力指数随着干旱胁迫程度加重呈降低的趋势,萌发时滞随干旱胁迫程度加重而升高。(3)温度和干旱胁迫的交互作用对种子萌发具有显著影响。干旱胁迫下,不同温度条件下种子萌发的表现不同。在35% PEG-6000胁迫下,20 ℃和25 ℃时,种子萌发率分别为8.89%和15.55%,显著高于15 ℃(0%)。综上所述,适宜种子萌发的温度为20 ℃和25 ℃,适宜幼苗早期生长最佳温度为25 ℃;干旱程度越大,种子萌发受到的抑制作用越强,干旱胁迫对种子萌发的影响高于温度因素,适当增温可缓解干旱胁迫对种子萌发的抑制作用。
关键词: 濒危植物, 狭叶坡垒, 温度, 干旱胁迫, 种子萌发
中图分类号: Q945
文献标识码: A
文章编号: 1000-3142(2024)01-0068-09
Effects of simulated drought by PEG-6000 on the
germination of Hopea chinensis seeds under
different temperature conditions
LIANG Huizi1, YANG Jisheng1, QIN Yi2, PAN Weihu2, XIAO Yufei1,
HUANG Ronglin1, WANG Renjie1, JIANG Yi1, LIU Xiongsheng1*
( 1. Guangxi Key Laboratory of Superior Trees Resource Cultivation, Guangxi Forestry Research Institute, Nanning 530002, China;
2. Guangxi Fangcheng Golden Camellias National Nature Reserve, Fangchenggang 538021, Guangxi, China )
Abstract: Hopea chinensis is distributed in China and its seeds are typically reclacitrant. In order to investigate the germination adaptability of seeds to temperature and moisture, we carried out an experiment with three different constant temperatures (15, 20, 25 ℃) controlled by artificial climate incubators, and used six polyethylene glycol (PEG-6000) concentrations (0%, 5%, 10%, 15%, 25% and 35%) to simulate drought stress. The germination characteristics of these seeds were carefully examined. The results were as follows: (1) Temperature exerted a significant influence on seed germination. Regardless of the level of drought stress applied, higher temperatures positively affected the germination percentage, germination energy, germination index, radicle length, sprout length and vitality index of seeds, higher temperatures led to shorter germination delay and increased fluctuations in the germination duration. (2) Drought stress had a significant impact on seed germination. Under identical temperature conditions, increasing levels of drought stress resulted in decreasing of germination percentage, germination energy, germination index, germination duration, radicle length, sprout length and vitality index of seeds. Moreover, higher levels of drought stress led to longer germination delay of seeds. (3) The interaction between temperature and drought stress had a significant influence on seed germination. Under drought stress, seed germination exhibited different responses under different temperature conditions. Specifically, under conditions where a PEG-6000 concentration of 35% was applied as drought stress treatment, the germination percentage at temperatures of 20 ℃ and 25 ℃ were significantly higher than those observed at 15 ℃ (8.89% and 15.55% compared to lower percentage at 15 ℃). However, it was important to note that when subjected to both 35% PEG-6000 and a temperature of 15 ℃, the seeds no longer germinate. In summary, our results indicate that the suitable temperatures for seed germination are identified as being at 20 ℃ and 25 ℃. It is determined that the most favorable temperature for early seedling growth is 25 ℃. As the drought stress intensifies, the inhibitory effect on seed germination become stronger, the effect of drought stress on seed germination is higher than that of temperature. Additionally, moderate warming can help alleviate the inhibitory effect of drought stress on seed germination.
Key words: Endangered plant, Hopea chinensis, temperature, drought stress, seed germination
种子萌发是植物种群自然更新的重要阶段,对外界环境压力的反应最为敏感(李小双等,2007)。种子能否萌发、出苗受到多种生态因子的综合影响(鱼小军等,2006),它对环境变化的适应能力是植物适应环境的重要体现(雷春英等,2020),与植物本身的特性和适应环境的生态策略有关(鱼小军等,2006;李小双等,2007)。不同植物种子萌发所需的条件不同,对环境的适应性表现不同。因此,探究生态因子对种子萌发的影响及其机制一直都是研究热点(雷春英等,2020;范翠枝等,2021;关正等,2022)。
影响种子萌发的重要生态因子包括温度和水分。有的植物种子在变温条件下利于萌发(关正等,2022),有的则为恒温、高温(鱼小军等,2004)或低温(盛海燕等,2004)。不同植物种子在不同水分胁迫条件下的萌发行为存在差异,众多学者通过PEG-6000水溶液模拟干旱胁迫对植物种子萌发期的抗旱性进行研究(李志萍等,2013;闫兴富等,2016)。低浓度干旱胁迫可促进种子萌发(汪建军等,2016),如5%~10%浓度的PEG-6000溶液可促进水杉(Metasequoia glyptostroboides)种子发芽和出苗整齐(吴漫玲等,2020),5%~10% PEG-6000处理促使小溪洞杜鹃(Rhododendron xiaoxidongense)种子打破休眠(李丹丹等,2022)。随着PEG-6000溶液浓度增加,种子萌发受到抑制作用增强(汪建军等,2016;陈士超等,2017),种子发芽率、发芽指数等萌发指标随之下降(刘雪松等,2019)。温度和水分的交互作用对种子萌发具有综合影响,如适当增温可缓解较高浓度PEG溶液模拟干旱胁迫对歪头菜(Vicia unijuga)种子萌发的抑制作用(唐伟和南志标,2019),在较低温度下柠条锦鸡儿(Caragana korshinskii)种子对干旱胁迫的耐受性最强(闫兴富等,2016)。研究种子萌发所需的温度和水分条件,尤其是濒危植物种子,有助于了解其萌发特性,掌握种群动态、调控与更新、生态适应性等(Steven, 1991),可为揭示其濒危机制、制定种群保护与恢复策略提供理论依据(张俊杰等,2018)。
狭叶坡垒(Hopea chinensis)是龙脑香科(Dipterocarpaceae)坡垒属(Hopea)常绿乔木,是我国二级重点保护野生植物,全国极小种群物种,被世界自然保护联盟(International Union For Conservation of Nature)列為极危(CR)物种。目前,对狭叶坡垒的研究集中在种群生态(黄仕训等,2008a)、光合特性(莫凌等,2009)、幼苗生理特性(周太久等,2013)、传粉生物学(卢清彪等,2020)、野外种子分布格局及种子萌发特性(唐文秀等,2009)和种子生理特性(黄宁等,2022)等方面,以上研究对狭叶坡垒的种群生态学和生物学特性有了进一步的了解。狭叶坡垒种子生物学特性研究表明,种子含水量高,无休眠期,在湿热环境中易萌发(黄仕训等,2008b)。在自然生境中,成熟散落于岩石上的种子因风干快速失水而失去活力(唐文秀等,2009)。黄宁等(2022)研究表明,自然脱水第8天时,种子萌发率仅为51.67%,接近半致死,干旱可能是影响种子萌发的重要因素之一。已有研究发现,种子在保湿条件12~13 ℃下贮藏,延迟至25 d后播种,发芽率低,仅为37%(黄仕训等,2008b),并且狭叶坡垒种子于12月至翌年1月成熟,此时正值广西西南地区的冬季,温度较低,低温可能是影响种子萌发的另一个重要因素。狭叶坡垒区系具有热带性质,萌发环境需湿热条件,然而,目前温度和水分条件对狭叶坡垒种子萌发的影响尚不清楚。鉴于此,本研究以狭叶坡垒种子为研究对象,采用不同温度和不同浓度PEG-6000溶液模拟干旱胁迫处理的方法,对种子的萌发特性进行分析,拟探讨以下问题:(1)狭叶坡垒种子在不同温度和不同干旱胁迫处理下的适应机制;(2)种子萌发对不同温度和不同干旱胁迫的响应策略;(3)不同温度与不同干旱胁迫处理及二者交互作用下,种子萌发的差异性。以期探究种子萌发所需的适宜生态条件,为狭叶坡垒种质资源保存和苗木繁育提供理论依据。
1 材料与方法
1.1 试验材料
试验种子于2020年1月采自广西壮族自治区防城金花茶国家级自然保护区南山保护站(108°02′33″ E、21°43′34″ N,海拔196 m,坡向朝南,坡度10°),供试种子千粒重为470.73 g,含水量为42.57%。所有种子均为母树上自然脱落期间而采集的成熟种子。采集后将种子混合均匀,立即带回广西壮族自治区林业科学研究院广西优质用材林资源培育重点实验室,去除杂质后立即开展萌发试验。所有采集活动均获得当地管理部门和主管部门许可。
1.2 试验方法
1.2.1 不同温度下PEG-6000模拟干旱胁迫对狭叶坡垒种子萌发影响试验[HTSS] 参照陈士超等(2017)的方法,分别配置PEG-6000浓度为0%(蒸馏水)、5%、10%、15%、25%和35%6个模拟干旱胁迫处理,每个处理设置3个重复,每个重复选择30粒去除果翅的健康种子,用0.2%高锰酸钾溶液浸泡消毒5 min,流水冲洗后置于蒸馏水中浸泡24 h。之后将种子均匀播于铺有2 层滤纸的120 mm培养皿中,分别加入PEG-6000处理液10 mL,根据狭叶坡垒原生境的气候条件及预实验结果,分别置于15、20、25 ℃,3 000 lx 12 h·d-1,80% 相对湿度(relative humidity,RH)的培养箱内。每2 d更换1次滤纸和PEG-6000溶液。
[HTK]1.2.2 狭叶坡垒种子活力和萌发特性试验[HTSS] 每天定时观察记录种子萌发情况,以胚根生长到种子长1/2时视为发芽。20 d试验结束,测定胚根长(redicle length,Lr)和芽长(sprout length,Ls)。种子萌发指标包括萌发率(gemination percentage,GP)、萌发指数(germination index,GI,从试验开始培养到第一颗种子萌发所需的时间)、萌发时滞(germination delay,GD)、萌发历期(germination duration,D,种子萌发过程所需的时间)、萌发势(germination energy,GE)以及活力指数(vitality index,VI),计算公式如下(闫兴富等,2016;雷春英等,2020):
GP=(发芽种子总数/供试种子数)×100%[JY](1)
GI=∑(时间t天萌发的种子数/对应的萌发天数)[JY](2)
GE=(7 d内正常发芽种子数/供试种子数)×100%[JY](3)
VI=GI×(Lr+Ls)[JY](4)
式中, Lr和Ls分别为萌发试验结束时胚根长(cm)和芽长(cm)(本研究中幼苗根长为胚根及胚轴的总长度)。
1.3 数据分析
用SPSS 19.0对不同处理下狭叶坡垒种子各萌发指标进行方差分析(ANOVA)和多重比较(采用邓肯氏新复极差法),采用双因素方差分析探究温度和干旱胁迫交互作用对种子萌发的影响(P<0.05)。以上统计分析显著性水平均设定为α= 0.05。
2 结果与分析
2.1 不同温度对种子萌发的影响
由表1可知,0% PEG-6000浓度处理下,20 ℃和25 ℃时种子萌发率、萌发势、萌发指数、胚根长均显著高于15 ℃,萌发时滞均显著低于15 ℃,其中萌发率分别比15 ℃时显著高59%和56%,萌发势比15 ℃时显著高110%和120%,萌发指数比15 ℃时显著低87%和99%,萌发时滞比15 ℃时显著低29%和50%,胚根长比15 ℃时显著高21%和46%,活力指数比15 ℃时显著高344%和495%。25 ℃时种子萌发历期、胚根长、芽长和活力指数均最高,分别为14.3 d、6.28 cm、6.56 cm、27.62,其中萌发历期、胚根长和活力指数显著大于其余2种温度。
5% PEG-6000胁迫下,20 ℃和25 ℃时种子萌发率、萌发势均显著高于15 ℃,萌发率分别比15 ℃时显著高42%和46%,萌发势分别比15 ℃时显著高100%和111%;25 ℃时种子萌发指数、萌发历期、胚根長、芽长和活力指数均最高,其中萌发指数、萌发历期、胚根长和活力指数显著高于其余2种温度。10% PEG-6000胁迫下,20 ℃和25 ℃时种子萌发率、萌发势、萌发指数均显著高于15 ℃,萌发率分别比15 ℃时显著高48%和33%,萌发势分别比15 ℃时显著高175%和200%,萌发指数分别比15 ℃时显著高77%和75%,萌发时滞均比15 ℃时显著低35%,萌发历期比15 ℃时显著高44%和12%;25 ℃时种子胚根长、芽长和活力指数均最高,其中胚根长和活力指数显著高于其余2种温度。
15% PEG-6000胁迫下,3种温度处理下种子萌发率均低于50%,种子达半致死率。25 ℃时种子萌发势、胚根长、芽长和活力指数均最高。25% PEG-6000胁迫下,25 ℃时种子萌发率、萌发指数、胚根长和活力指数最高,种子萌发时滞最低,25 ℃萌发时滞比20 ℃和15 ℃时显著低23%和28%,胚根长比20 ℃和15 ℃时显著高21%和35%,活力指数比15 ℃显著增高142%。35% PEG-6000胁迫下,3种温度间的萌发率与其他浓度的干旱胁迫相较最低,仅在20 ℃和25 ℃条件下种子萌发且萌发率分别为8.89%和15.55%,萌发受到严重抑制。
2.2 不同PEG-6000浓度模拟干旱胁迫对种子萌发的影响
不同干旱程度对狭叶坡垒种子萌发的影响见表1。15 ℃时0%和5%胁迫下萌发率、萌发势、萌发指数、胚根长和活力指数显著高于其余PEG-6000浓度处理;0%、5%、10%胁迫下种子萌发时滞显著低于其余3个胁迫浓度;0%、5%、10%、15%胁迫浓度处理下种子萌发历期显著高于其余2个胁迫浓度。
20 ℃时,0%浓度处理下种子萌发率、萌发指数和活力指数均显著高于其余胁迫浓度;0%、5%、10%胁迫下种子萌发时滞显著低于其余3个胁迫浓度;0%和5%胁迫下萌发历期显著低于10% PEG-6000胁迫,随着干旱程度增加呈先升后降的趋势。
25 ℃时,0%和5%胁迫下萌发率、萌发指数、胚根长、芽长和活力指数显著高于其余胁迫浓度,萌发时滞显著低于其余胁迫浓度;0%浓度处理下种子萌发势、萌发历期均显著高于其余胁迫浓度。
2.3 温度和PEG-6000模拟干旱胁迫对种子萌发的相互作用
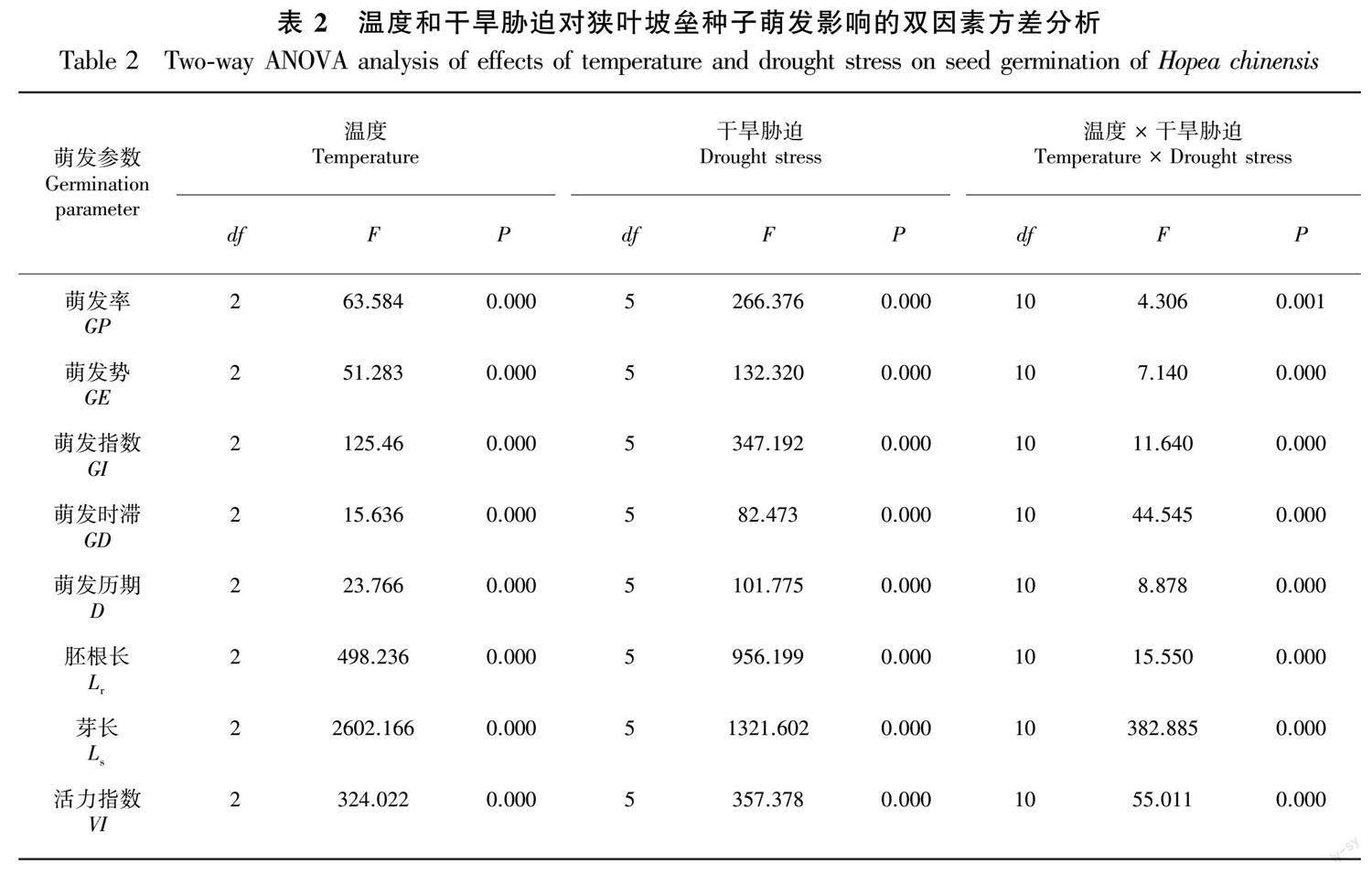
由表2可知,温度、PEG-6000模拟干旱胁迫及两者间的交互作用对狭叶坡垒种子8个萌发指标均有极显著影响(P<0.01)。不同温度条件下,种子萌发对PEG-6000胁迫所表现的差异反映了温度和PEG-6000模拟干旱胁迫对狭叶坡垒种子萌发的交互影响。例如,5% PEG-6000胁迫下,20 ℃和15 ℃种子萌发率分别为82.22%和57.78%,两者相差24.44%;10% PEG-6000胁迫下,20 ℃和15 ℃种子萌发率分别为68.89%和46.67%,两者相差22.22%,较5% PEG-6000胁迫下两者差异降低(表1)。
3 讨论与结论
种子萌发需要适宜的生态条件, 各个生态因
子的相互作用影响着种子活力(鱼小军等,2006)。本研究结果发现,20 ℃和25 ℃时,狭叶坡垒种子萌发率、萌发势、萌发指数、活力指数较高,胚根和胚芽较长,萌发时滞较短;而15 ℃时,萌发率、萌发势、萌发指数和活力指数最低,胚芽不生长,萌发受到抑制,以上萌发指标表明20~25 ℃是狭叶坡垒种子萌发的适宜温度条件。其中,20 ℃时种子萌发历期显著低于25 ℃,萌发时间最短,表明20 ℃时全部种子萌发时间加快,25 ℃时种子于第2天开始萌发,胚根和胚芽最长,说明25 ℃为早期幼苗最佳生长温度。适宜的温度保证种子体内酶具有较高活性,使得酶促反应速度、呼吸作用和种子内部贮藏的营养物质分解加快,满足种子萌发所需的物质和能量(秦爱丽等,2020)。若外界温度过高或过低,会改变种子体内水解酶性质,影响膜透性和膜结合蛋白的活性,从而抑制种子萌发(宋兆伟等,2010)。不同植物的最适萌发温度不同,与其原生境密不可分,适时萌发的机制是植物对原生境适应性的体现(李阳等,2021)。狭叶坡垒在15 ℃时萌发受到抑制,与望天树(Shorea wantianshuea)、番龙眼(Pometia pinnata)等热带顽拗性种子不耐低于15 ℃的低温结果相一致(文彬等,2002;闫兴富和曹敏,2006),可能是因为温度较低会造成低温损伤,说明其不适合在温度较低的地区生长。黄仕训等(2008b)研究表明,狭叶坡垒仅分布在广西十万大山海拔低于600 m的区域,北热带季风气候区的气温为其提供了生长所需条件,也限制了分布范围。种子成熟于冬季,在分布区的自然生境中,偏低的温度会使种子面临难以形成幼苗的风险,这可能是影响狭叶坡垒种群自然更新的重要因素。此外,狭叶坡垒种子在湿润低温条件下仍可萌发,表明这是一种适应自然生境形成的生存策略,在低温条件下,若遇到降水种子及时萌发形成幼苗以提高存活的机会。
种子萌发离不开足量的水分(秦爱丽等,2020)。在10% PEG-6000胁迫下,种子萌发受到抑制,在15%胁迫时种子萌发已达半致死率,在35%胁迫下严重抑制了种子萌发和幼苗生长,与干旱胁迫下胀果甘草种子(Glycyrrhiza inflata)(史薇等,2010)、紫花苜蓿(Medicago sativa)和高粱(Sorghum bicolor)种子(李文娆等,2009)等研究结果类似,可能因为干旱胁迫抑制种子保护酶活性和体内渗透调节物质作用,从而抑制正常的生理代谢(李志萍等,2013)。狭叶坡垒种子萌发对干旱胁迫反应敏感,可能与其原生境和繁育习性有关。狭叶坡垒是热带季节性雨林的代表树种,种子成熟期正值最干旱的季节,若不具备萌发所需的湿润条件,萌发受到抑制,可能是目前狭叶坡垒只在沟谷两侧的茂密森林和溪水岸边分布的重要原因(黄仕训等,2008b)。
温度和干旱胁迫对种子萌发有交互影响。20 ℃时,种子萌发历期随干旱胁迫增加呈先增后降的趋势,说明种子在适宜温度下调整萌发行为,通过延长萌发时间来抵御不稳定的环境变化,减少死亡风险,以确保种子能在适宜条件下进行萌发(王慧慧等,2016;Zhang et al., 2022)。在5% PEG-6000胁迫条件下,20 ℃和25 ℃时种子萌发率在分别为82.22%和84.45%,表明在低浓度干旱胁迫下,种子仍具有较高的萌发率,可能因为植物通过增加渗透调节物质和提高保护性酶的活性等措施来提高抗旱性(麦苗苗等,2009;张中峰等,2012)。在35% PEG-6000胁迫条件下,20 ℃和25 ℃时种子萌发率分别为8.89%和15.35%,胚根长分别为1.63 cm和2.04 cm,具有一定的生长活力,表明温度升高可缓解干旱胁迫对种子萌发的抑制作用(唐伟和南志标,2019)。此时,胚根仍在生长,利于吸收水分,表明在35% PEG-6000胁迫下种子表现出一定的活力,可能是种子在长期生活环境中形成的萌发策略,具有较强的环境适应性(唐伟和南志标,2019)。应当注意,在35% PEG-6000胁迫下,15 ℃时种子不再萌发,表明同时遭遇低温和重度干旱胁迫,这样会对狭叶坡垒种子萌发具有致命的打击。
综上所述,温度和干旱胁迫均会影响狭叶坡垒种子的萌发进程。15 ℃时种子萌发受到抑制,20~25 ℃为种子萌发的适宜温度,其中25 ℃为幼苗早期生长最佳温度,种子对水分要求高,随着干旱胁迫程度增强,种子萌发受到的抑制作用增强,20~25 ℃可缓解35% PEG-6000干旱胁迫带来的抑制作用,低温和干旱可能是影响狭叶坡垒幼苗分布和自然更新的重要因素。人工培育狭叶坡垒,在保持水分充足情况下可以采用20~25 ℃促进种子萌发,种子萌发后保持25 ℃左右环境温度有利于幼苗生长。
参考文献:
CH EN SC, WANG M, WANG J, et al., 2017. Response of seed germination and seedling physiological characteristics of Medicago sativa to the simulated osmotic potential of PEG6000 [J]. Chin J Appl Ecol, 28(9): 2923-2931. [陳士超, 王猛, 汪季, 等, 2017. 紫花苜蓿种子萌发及幼苗生理特性PEG-6000模拟渗透势的响应 [J]. 应用生态学报, 28(9): 2923-2931.]
FAN CZ, WU XY, GUAN X, et al., 2021. Concentration effects and its physiological mechanism of soaking seeds with brassinolide on tomato seed germination under salt stress [J]. Acta Ecol Sin, 41(5): 1857-1867. [范翠枝, 吴馨怡, 关欣, 等, 2021. 油菜素内酯浸种对盐胁迫番茄种子萌发的影响及其生理机制 [J]. 生态学报, 41(5): 1857-1867.]
GUANG Z, WANG LJ, DUAN L, et al., 2022. Effects of PEG simulated drought stress on seed germination of Abutilon theophrasti medicus [J]. Seed, 41(8): 66-70. [关正, 王丽君, 段玲, 等, 2022. PEG模拟干旱胁迫对苘麻种子萌发的影响 [J]. 种子, 41(8): 66-70.]
HUANG N, LIU XS, LIAO NY, et al., 2022. Study on desiccation sensitivity of seeds of extremely endangered plant Hopea chinensis [J]. Guangxi For Sci, 51(5): 634-640. [黄宁, 刘雄盛, 廖南燕, 等, 2022. 极度濒危植物狭叶坡垒种子脱水敏感性研究 [J]. 广西林业科学, 51(5): 634-640.]
HUANG SX, CHEN H, PAN B, et al., 2008a. Characteristics of Hopea chinensis community, an endemic and endangered species in Guangxi [J]. Acta Bot Boreal-Occident Sin, 28(1): 164-170. [黄仕训, 陈泓, 盘波, 等, 2008a. 广西特有濒危植物狭叶坡垒群落特征研究 [J]. 西北植物学报, 28(1): 164-170.]
HUANG SX, CHEN H, TANG WX, et al., 2008b. Biological and ecological characteristics of Hopea chinensis, a plant endemic to Guangxi [J]. Biodivers Sci, 16(1): 15-23. [黄仕训, 陈泓, 唐文秀, 等, 2008b, 狭叶坡垒生物生态学特征及致濒原因研究 [J]. 生物多样性, 16(1): 15-23.]
LEI CY, ZHANG H, ZHANG DD, et al., 2020. Effects of temperature, salinity and light on seed germination of Betula halophila [J]. Chin Wild Plant Resour, 39(11): 39-43. [雷春英, 张浩, 张丹丹, 等, 2020. 温度、盐分和光照对濒危植物盐桦(Betula halophila)种子萌发特性的影响 [J]. 中国野生植物资源, 39(11): 39-43.]
LI DD, LI XH, LIU J, et al., 2022. Effects of single treatments of gibberellin and drought stress on seed germination of rare and endangered plant Rhododendron xiaoxidongense [J]. J Plant Resour Environ, 31(4): 57-64. [李丹丹, 李晓花, 刘杰, 等, 2022, 赤霉素和干旱胁迫单一处理对珍稀濒危植物小溪洞杜鹃种子萌发的影响 [J]. 植物资源与环境学报, 31(4): 57-64.]
LI WR, ZHANG SQ, SHAN L, 2009. Seeds germination characteristics and drought tolerance of alfalfa and sorghum seedling under water stress [J]. Acta Ecol Sin, 29(6): 3066-3074. [李文嬈, 张岁岐, 山仑, 2009. 水分胁迫下紫花苜蓿和高粱种子萌发特性及幼苗耐旱性 [J]. 生态学报, 29(6): 3066-3074.]
LI XS, PENG MC, DANG CL, 2007. Research progress on natural regeneration of plants [J]. Chin J Ecol, 26(12): 2081-2088. [李小双, 彭明春, 党承林, 2007. 植物自然更新研究进展[J]. 生态学杂志, 26(12): 2081-2088.]
LI Y, LI X, DING FB, et al., 2021. Responses of Potentilla rupestris seeds germination to environmental factors [J]. Seed, 40(4): 85-89. [李阳, 李想, 丁芳兵, 等, 2021. 石生委陵菜种子萌发对环境因素的响应 [J]. 种子, 40(4): 85-89. ]
LI ZP, ZHANG WH, CUI YC, 2013. Effects of PEG simulated drought stress on seed germination and growth physiology of Quercus variabilis [J]. Acta Bot Boreal-Occident Sin, 33(10): 2043-2049. [李志萍, 张文辉, 崔豫川, 2013. PEG模拟干旱胁迫对栓皮栎种子萌发及生长生理的影响 [J]. 西北植物学报, 33(10): 2043-2049.]
LIU XS, 2019. Effects of temperature, light, and PEG on seed germination in different ecotypes of Achnatherum inebrians [J]. Pratac Sci, 36(6): 1600-1607. [刘雪松, 2019. 温度、光照及PEG胁迫对不同生态型醉马草种子萌发的影响 [J]. 草业科学, 36(6): 1600-1607.]
LU QB, ZHU XZ, LIU CQ, et al., 2020. Pollination biology of Hopea chinensis [J]. Guihaia, 40(11): 1628-1637. [卢清彪, 朱晓珍, 刘长秋, 等, 2020. 狭叶坡垒传粉生物学初探 [J]. 广西植物, 40(11): 1628-1637.]
MAI MM, SHI DX, WANG ML, et al., 2009. Seed germination and seedling growth of Cercidiphyllum japonicum with PEG treatment [J]. Sci Silv Sin, 45(10):94-99. [麦苗苗, 石大兴, 王米力, 等, 2009. PEG处理对连香树种子萌发与芽苗生长的影响 [J]. 林业科学, 45(10): 94-99.]
MO L, TANG WX, MAO SZ, et al., 2009. Photosynthesis characteristics of rare and endangered plant Hopea chinensis [J]. J Fujian Coll For, 29(4): 357-361. [莫凌, 唐文秀, 毛世忠, 等, 2009. 珍稀濒危植物狭叶坡垒的光合特性 [J]. 福建林学院学报, 29(4): 357-361.]
QIN AL, GUO QS, MA FQ, et al., 2020. Effects of temperature, light and water conditions on seed germination of Thuja sutchuenensis Franch. [J]. Seed, 39(2): 15-20. [秦爱丽, 郭泉水, 马凡强, 等, 2020. 温度、光照和水分对珍稀濒危树种崖柏种子萌发的影响 [J]. 种子, 39(2): 15-20.]
SHENG HY, GE Y, CHANG J, et al., 2004. Influence of environmental factors on seed germination of two species in Umbellaceae [J]. Acta Ecol Sin, 24(2): 221-226. [盛海燕, 葛滢, 常杰, 等, 2004. 环境因素对伞形科两种植物种子萌发的影响 [J]. 生态学报, 24(2): 221-226.]
SHI W, XU HL, ZHAO XF, et al., 2010. Physiological and biochemical responses to drought stress during seed germination of Glycyrrhiza inflata [J]. Acta Ecol Sin, 30(8): 2112-2117. [史薇, 徐海量, 赵新风, 等, 2010. 胀果甘草种子萌发对干旱胁迫的生理响应 [J]. 生态学报, 30(8): 2112-2117.]
SONG ZW, HAO LZ, HUANG ZY, et al., 2010. Effects of light and temperature on the germination of Pugionium cornutum (L.) Gaertn. and Pugionium dolabratum Maxim. seeds [J]. Acta Ecol Sin, 30(10): 2562-2568. [宋兆偉, 郝丽珍, 黄振英, 等, 2010. 光照和温度对沙芥和斧翅沙芥植物种子萌发的影响 [J]. 生态学报, 30(10): 2562-2568.]
STEVEN DD, 1991. Experiments on mechanisms of establishment in old-field succession: seedling emergence [J]. Ecology, 72 (3): 1066-1075.
TANG W, NAN ZB, 2019. Effects of osmotic stress by PEG-6000 on germination of Vicia unijuga seeds under different temperature conditions [J]. Pratac Sci, 36(5): 1323-1332. [唐伟, 南志标, 2019. 不同温度下PEG-6000渗透胁迫对歪头菜种子发芽的影响 [J]. 草业科学, 36(5): 1323-1332.]
TANG WX, MAO SZ, PAN B, et al., 2009. Spatial distribution pattern of seed rain and seed germination characteristics of endangered plant Hopea chinensis [J]. J Fujian Coll For, 29(2): 149-154. [唐文秀, 毛世忠, 盘波, 等, 2009. 狭叶坡垒种子雨的空间格局及种子萌发特性 [J]. 福建林学院学报, 29(2): 149-154.]
WANG HH, WANG PC, ZHAO G, et al., 2016. Seed size and germination strategy of Sophora davidii under drought stress [J]. Acta Ecol Sin, 36(2): 335-341. [王慧慧, 王普昶, 趙钢, 等, 2016. 干旱胁迫下白刺花种子大小与萌发对策 [J]. 生态学报, 36(2): 335-341.]
WANG JJ, MA AW, WANG ZG, et al., 2016. Effects of different temperature and moisture conditions on seed germination of Festuca sinensis [J]. Acta Pratac Sin, 25(4): 73-80. [汪建军,麻安卫, 汪治刚, 等, 2016. 不同温度和PEG处理对中华羊茅种子萌发的影响 [J]. 草业学报, 25(4): 73-80.]
WEN B, YIN SH, LAN QY, et al., 2002. Ecological characteristics of seed germination of Pometia tomentosa [J]. Guihaia, 22(5): 408-412. [文彬, 殷寿华, 兰芹英, 等, 2002. 绒毛番龙眼种子萌发生态特性的研究 [J]. 广西植物, 22(5): 408-412. ]
WU ML, ZHU J, AI XR, et al., 2020. Influences of PEG simulating drought stress on seed germination of Metasequoia glyptostroboides under different temperatures [J]. Guihaia, 40(11): 1691-1698. [吴漫玲, 朱江, 艾训儒, 等, 2020. 不同温度条件下PEG模拟干旱胁迫对水杉种子萌发的影响 [J]. 广西植物, 40(11): 1691-1698.]
YAN XF, CAO M, 2006. Influence of light and temperature on the germination of Shorea wantianshuea (Dipterocarpaceae) seeds [J]. Chin Bull Bot, 23(6): 642-650. [闫兴富, 曹敏, 2006. 光照和温度对望天树种子萌发的影响 [J]. 植物学通报, 23(6): 642-650. ]
YAN XF, ZHOU LB, SI BB, et al., 2016. Stress effects of simulated drought by polyethylene glycol on the germination of Caragana korshinskii Kom. seeds under different temperature conditions [J]. Acta Ecol Sin, 36(7): 1989-1996. [闫兴富, 周立彪, 思彬彬, 等, 2016. 不同温度下PEG-6000模拟干旱对柠条锦鸡儿种子萌发的胁迫效应 [J]. 生态学报, 36(7): 1989-1996.]
YU XJ, SHI SL, LONG RJ, et al., 2006. Research progress on effects of ecological factors on seed germination [J]. Pratac Sci, 23(10): 44-49. [鱼小军, 师尚礼, 龙瑞军, 等, 2006. 生态条件对种子萌发影响研究进展 [J]. 草业科学, 23(10): 44-49.]
YU XJ, WANG YR, ZENG YJ, et al., 2004. Effects of temperature and osmotic potential on seed germination of Cleistogenes songorica and Plantago lessingii [J]. Acta Ecol Sin, 24(5): 883-887. [鱼小军, 王彦荣, 曾彦军, 等, 2004. 温度和水分对无芒隐子草和条叶车前种子萌发的影响 [J]. 生态学报, 24(5): 883-887. ]
ZHANG JJ, CHAI SF, WEI X, et al., 2018. Germination characteristics of the seed of a rare and endangered plant,Garcinia paucinervis [J]. Sci Silv Sin, 54(4): 174-185.[张俊杰, 柴胜丰, 韦霄, 等, 2018. 珍稀濒危植物金丝李种子的萌发特性 [J]. 林业科学, 54(4): 174-185.]
ZHANG R, CHEN DL, LIU HZ, et al., 2022. Effect of temperature and water potential on the germination of seeds from three different populations of Bidens pilosa as a potential Cd hyperaccumulator [J]. BMC Plant Biol, 22(1): 1-13.
ZHANG ZF, YOU YM, HUANG YQ, et al., 2012. Effects of drought stress on Cyclobalanopsis glauca seedlings under simulating karst environment condition [J]. Acta Ecol Sin, 32(20): 6318-6325. [张中峰, 尤业明, 黄玉清, 等, 2012. 模拟喀斯特生境条件下干旱胁迫对青冈栎苗木的影响 [J]. 生态学报, 32(20): 6318-6325.]
ZHOU TJ, DENG T, TANG WX, et al., 2013. Physiological responses of seedlings of Hopea chinensis to PEG simulated drought stress [J]. Hubei Agric Sci, 52(24): 6079-6083. [周太久, 邓涛, 唐文秀, 等, 2013. 狭叶坡垒幼苗对PEG模拟干旱胁迫的生理响应 [J]. 湖北农业科学, 52(24): 6079-6083.]
( 责任编辑 李 莉 )
收稿日期: 2023-08-11
基金项目: 国家自然科学基金(32301464); 广西青年科学基金(2020GXNSFBA297043)。
第一作者: 梁惠子(1992-),硕士,工程师,主要从事环境生态学方面的研究,(E-mail)289631276@qq.com。
* 通信作者: 刘雄盛,硕士,副研究员,主要从事林木遗传育种研究,(E-mail)517261654@qq.com。
