云南火焰兰的花形态和花药发育的胚胎学特征
2024-03-10王乐骋陶凯锋朱永张锦罗艳李璐
王乐骋 陶凯锋 朱永 张锦 罗艳 李璐
摘 要: 關于珍稀濒危植物火焰兰属的研究资料较少。该文以云南火焰兰为研究对象,通过显微镜和石蜡切片技术观察了云南火焰兰的花形态和花药发育胚胎学特征,并结合现有资料探讨了其分类学意义。结果表明:(1)具有分类学意义的花形态特征为唇瓣三裂且花距不明显,合蕊柱圆柱形,花药帽紫色具黄条纹,花粉团一对且由粘盘及粘盘柄连接。(2)花药原基分化出一对侧生花药室,每个花药室的小孢子囊中央分化出一条偏离中轴的不育隔膜组织,发育为一对不等大的次生小孢子囊;花药成熟时,因不育隔膜组织降解而形成2个深裂花粉团。(3)发育完整的花药壁有5~6层,为“多层型”花药壁,包括表皮、2~3层药室内壁、中层和绒毡层;绒毡层细胞为单核、腺质型。花药成熟时,药室内壁发生纤维状加厚,花药室在远轴端开裂散粉。(4)小孢子母细胞经同时型胞质分裂,形成正四面体和左右对称排列的小孢子四分体,小孢子保持在四分体内,经有丝分裂发育为2-细胞型的四合花粉,排列紧密,形成坚固的花粉团;在花药发育过程中未观察到花粉败育现象。该研究结果为火焰兰属的分类学和保护生物学提供了新资料。
关键词: 火焰兰属, 形态学, 花粉团发育, 多层型花药壁, 小孢子发生, 雄配子体发育
中图分类号: Q944. 58
文献标识码: A
文章编号: 1000-3142(2024)01-0077-12
Floral morphology and embryological features of
anther development in Renanthera imschootiana
WANG Yuecheng1, TAO Kaifeng1, ZHU Yong1, ZHANG Jin1, LUO Yan2, LI Lu1*
( 1. Department of Biodiversity Conservation, Southwest Forestry University, Kunming 650224, China; 2. Department of Horticulture
and Gardening, Xishuangbanna Tropical Botanical Garden, Chinese Academy of Sciences, Mengla 666300, Yunnan, China )
Abstract: There are few data on the rare and endangered orchids of Renanthera Lour. Floral morphology and embryological features of anther development in R. imschootiana were investigated, and the taxonomic significance of R. imschootiana was analyzed by comparing with existing data. The results were as follows: (1) Flowers were characterized by such taxonomic features as tri-lobed labium with an unconspicous spur, cylindrical column, purple anther cap with yellow stripes, and a pair of deeply cleft pollinia connected by a well-developed stipe and a viscidium. (2) Anther primordium was differentiated into two lateral thecae containing two densely dyed microsporangia, and then, a sterile septum was formed longitudinally and unequally which divided each microsporangium into two jointly sub-microsporangium. The septum became absorbed during microsporogenesis and collapsed when anther ripened, resulting in a pair of deeply cleft pollinia. (3) The developed anther wall comprised 5-6 layers, including an epidermis, 2-3 endothecium, a middle layer, and a tapetum, which conformed to the massive type. Tapetal cell was uni-nucleated and glandular. (4) Microspore mother cells underwent a simultaneous cytokinesis during meiosis resulting in the tetrahedral or isobilateral microspore tetrads. Finally, each microspore retained in tetrads and produced 2-celled pollen via mitosis. Pollen tetrads were compactly arranged, leading to a hard cleft pollinia in each thecae. No pollen abortion was observed during anther development. The results provide new reference for taxonomy and conservation of Renanthera.
Key words: Renanthera, morphology, pollinium development, anther wall of massive tape, microsporogenesis, male gametophyte development
火焰兰属(Renanthera)隶属于兰科树兰亚科(Epidenroideae)万代兰族(Vandeae),物种数目相对较少,仅20种,主要分布于东南亚和热带喜马拉雅地区(吉占和等,1999)。中国有3种,地理分布较为狭窄(金效华等,2019)。其中,火焰兰(R. coccinea)分布略广,见于中国的云南南部、广西和海南,以及缅甸、泰国、老挝和越南(陈心启等,1999; 吴征镒,2003; 金效华等,2019)。云南火焰兰(R. imschootiana)为中国云南特有种,仅见于模式标本产地越南和中国的云南南部元江(吉占和等,1999)。近年发表的中华火焰兰(R. sinica)为中国特有种,仅分布于中国云南文山(刘仲健等,2003)。火焰兰属是一类大型附生兰,茎长、具攀附性,叶片革质,圆锥花序或总状花序,具多朵花,花大、花色鲜红或橘黄或带斑点,形似火焰,具有较高的观赏价值(陈心启等,1999),现已出现一些人工杂交园艺品种(罗剑飘等,2012; 曹雯静等,2014)。
火焰兰属地理分布狭窄、野生居群和标本馆馆藏稀少,不仅被《濒危野生动植物种国际贸易公约—附录I》收录(http: //www. cites.org.cn),还被列入《中国生物多样性红色名录—高等植物》的极危种名单(杨明森,2013)、《国家重点保护野生植物名录》(国家林业和草原局农业农村部,2021)和《云南省极小种群保护名录(2021版)》(孙卫邦,2021)。目前,关于火焰兰属的地理分布、居群结构、生物学和繁殖生态学等资料相对匮乏,亟待补充(李娟等,2018)。李娟等(2018)在2014 — 2018年间,经野外调查发现,云南火焰兰在云南元江有2个居群点,位于海拔1 100~1 200 m范围的常绿阔叶和落叶混交林中,附生在栓皮栎(Quercus variabilis)和毛枝青冈(Cyclobalanopsis helferiana)的树干上,共有12株,其中仅有1株更新幼苗,为衰退型种群,认为开花而坐果率低是其濒危的直接原因。因此,有必要开展云南火焰兰的生殖生物学和传粉生态学研究。
花形態和花药发育是兰科传粉生物学、繁育系统和胚胎发育的主要内容,包括传粉综合征(pollination syndrome)(Pansarin & Maciel, 2017; 黄双全和郭友好,2000; Zhang et al., 2022)、花药成熟时散粉单元(pollen dispersal unit)的分化(Singer et al., 2008)和花药发育过程中小孢子发生和雄配子体发育的胚胎学特征(Swamy, 1949; Rao, 1967; Sood, 1989; Kant & Hossain, 2010; 张锦等,2019;谭庆琴等,2020;李璐等,2020)。这些特征在兰科的亚科、族、亚族和属及其种间的物种分类和系统演化方面发挥着重要作用(Dressler, 1986, 1993; Freudenstein et al., 2002; Pridgeon et al., 2005; Freudenstein & Chase, 2015; Mosquera-Mosquera et al., 2019; 李璐,2023)。国产火焰兰属的3种植物在植株形态和花型花色方面极为相似,野外调查时极易混淆(陈心启等,1999; 吴征镒,2003; 金效华等,2019)。刘仲健等(2003)在首次描述中华火焰兰的形态特征时,重点比较分析了国产3个种的花形态特征,认为花序、花色与唇瓣裂片是火焰兰属物种鉴定的关键特征。Rusdi和Najwa(2020)观察了产自马来西亚的贝拉火焰兰(Renanthera bella)在人工栽培条件下的生长发育过程,为火焰兰属的人工繁育提供了新资料。目前,关于珍稀濒危植物火焰兰属的研究资料较少,考虑到花形态学和胚胎学的重要性,本文以云南火焰兰为研究对象,通过解剖镜和石蜡切片技术观察其花形态和花药发育的胚胎学特征,并结合现有资料比较分析了其分类学意义,为该属的物种鉴定和保护生物学提供新资料。
1 材料与方法
云南火焰兰引种栽培于西南林业大学科研教学实践基地(云南丰春坊生物科技有限公司兰花种质资源圃),正常生长开花,花期为每年的4—5月。2017—2020年,连续4年采集不同发育时期的花蕾和开放花,固定于50%酒精中,常温保存。取新鲜开放花于体式显微镜(Leica M165 FC)下解剖观察,记录唇瓣、合蕊柱、花药帽、花粉团、粘盘和粘盘柄等花结构的形态、颜色和大小等,并进行拍照处理。取固定后的花蕾材料,按形态大小分为5个发育阶段,每个阶段取5~10朵,经酒精系列脱水、二甲苯透明、浸蜡包埋,常规石蜡切片(Leica RM 2235),切片厚度为6~8 μm;用铁矾—苏木精染色、中性树脂胶封片(李正理,2006),烘干后在显微镜下观察、拍照,并使用图像编辑软件Adobe Photoshop 2020排版。
2 结果与分析
2.1 花形态结构
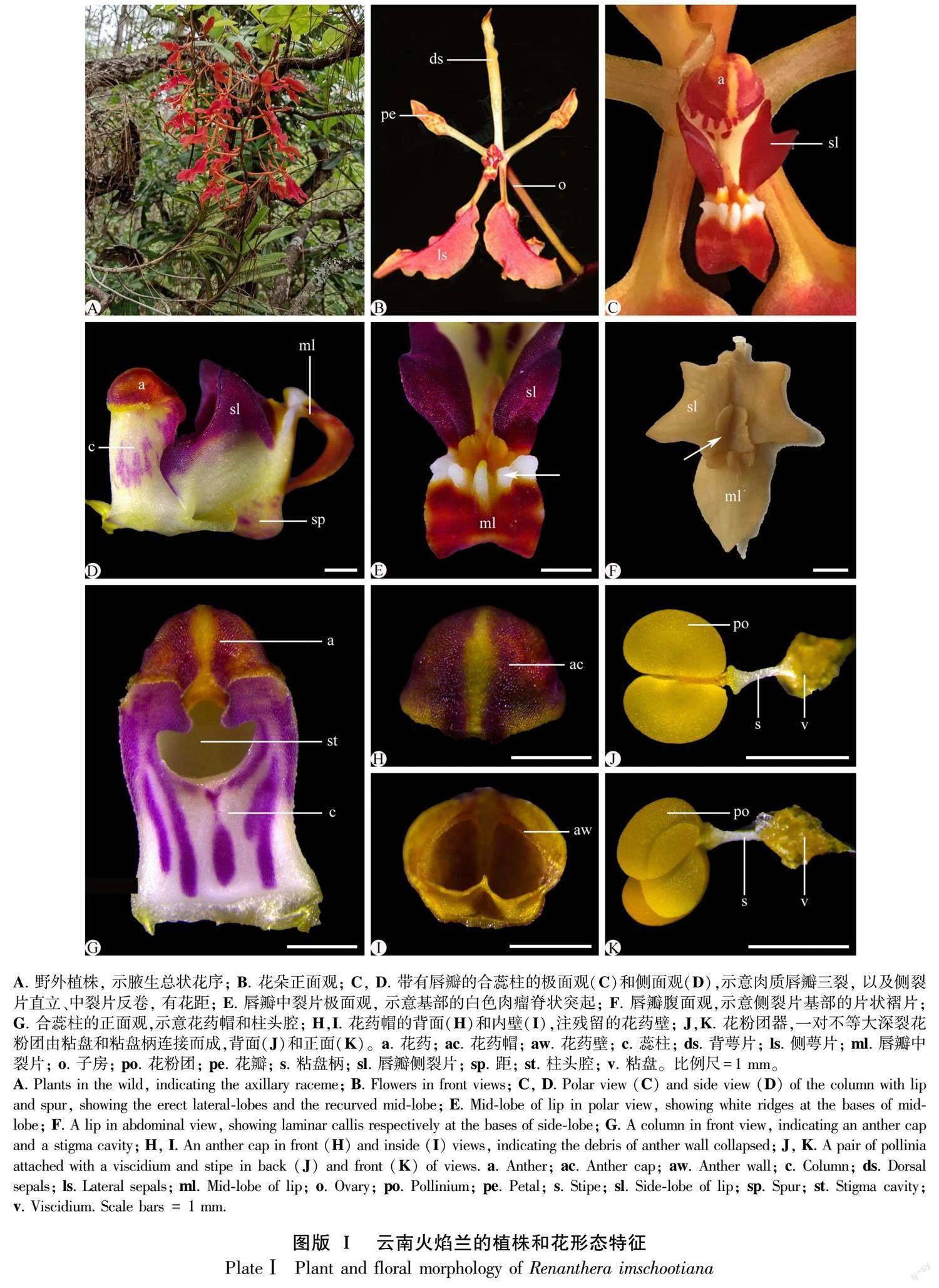
云南火焰兰为大型附生草本,开花时具有典型的腋生总状花序,花序轴纤细,多分枝,有5~30朵花(图版Ⅰ:A)。花两侧对称,中萼片和侧花瓣倒披针形,均为浅黄色;两枚侧萼片狭匙形,橘红色(图版1:B)。肉质唇瓣小而三裂,侧裂片深紫,三角形、先端锐尖,直立、高于合蕊柱,基部具2枚边缘不规则的黄色褶片(图版Ⅰ:C-E);中裂片卵形,反卷,先端深红色,基部具白色突起瘤状物(图版Ⅰ:E,F),花距不明显,囊状突起,长约2 mm。合蕊柱短,长4~5 mm,圆柱形、白底带紫红斑块,柱头腔深陷,下半部呈半圆形,上部开口处有深凹裂缝,构成向内合拢的蕊柱翅(图版Ⅰ:D,G)。花药帽盔状、半圆形,外部浅紫色,内壁明显分割为2室,在花药开裂花粉团释放后花药壁残留组织(图版Ⅰ:H,I)。每个药室有1枚花粉团,不等大深裂,远轴面较大,近轴面较小,由白色的细圆柱形粘盘柄和黄色的粘盘相连接,构成带有附属结构的花粉块或花粉团器(a pollinarium)(图版Ⅰ[KG-0.8mm]: J,K)。
2.2 一对深裂花粉团的发育模式
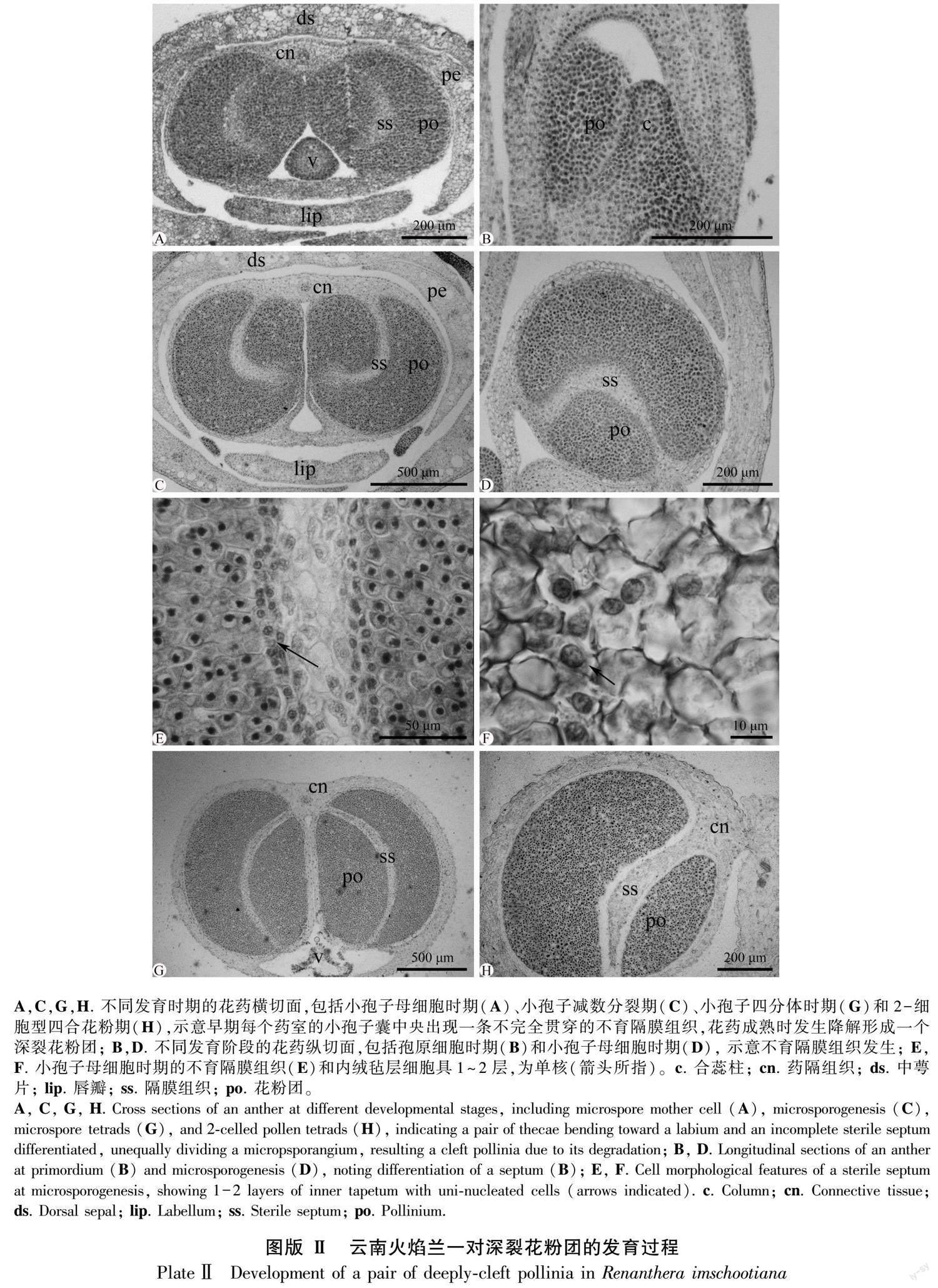
早期的花药原基分化出面向唇瓣弯曲生长的一对侧生并列花药室,每个花药室分化出一个小孢子囊,由核质比大、形态大小均匀的孢原细胞组成(图版Ⅱ:A,B),继而分化为一团染色较深的小孢子囊和一条染色较浅的不育隔膜组织(图版Ⅱ:A,C,D)。不育隔膜组织由染色较浅的薄壁细胞组成,明显区别于细胞分裂活动旺盛、染色较深的小孢子母细胞(图版Ⅱ:E)。它从近药隔组织端的小孢子囊中部出发,先沿花药室纵轴方向延伸生长至小孢子囊中部,再横折向药室内侧生长,止于近花药壁的5~6层小孢子母细胞处,分化为一条横折弯曲、不完全贯穿小孢子囊的组织(图版Ⅱ:C),分割出一对不等大、深裂的次生小孢子囊(图版Ⅱ:C)。在小孢子母细胞减数分裂期间,不育隔膜组织靠近小孢子母细胞处的细胞分化为1~2层形态较小、近圆球形单细胞核的内绒毡层(图版Ⅱ:E,F)。从小孢子四分体到雄配子体发育阶段,每个花药室的内绒毡层和不育隔膜组织细胞开始降解,形成空隙,把由小孢子囊发育而来的花粉囊分裂为一个不等大深裂的花粉团(图版Ⅱ:G, H)。因此,云南火焰兰的成熟花药里,产生了一对深裂的花粉团,这与花形态解剖观察结果一致。
2.3 多层型花药壁发育
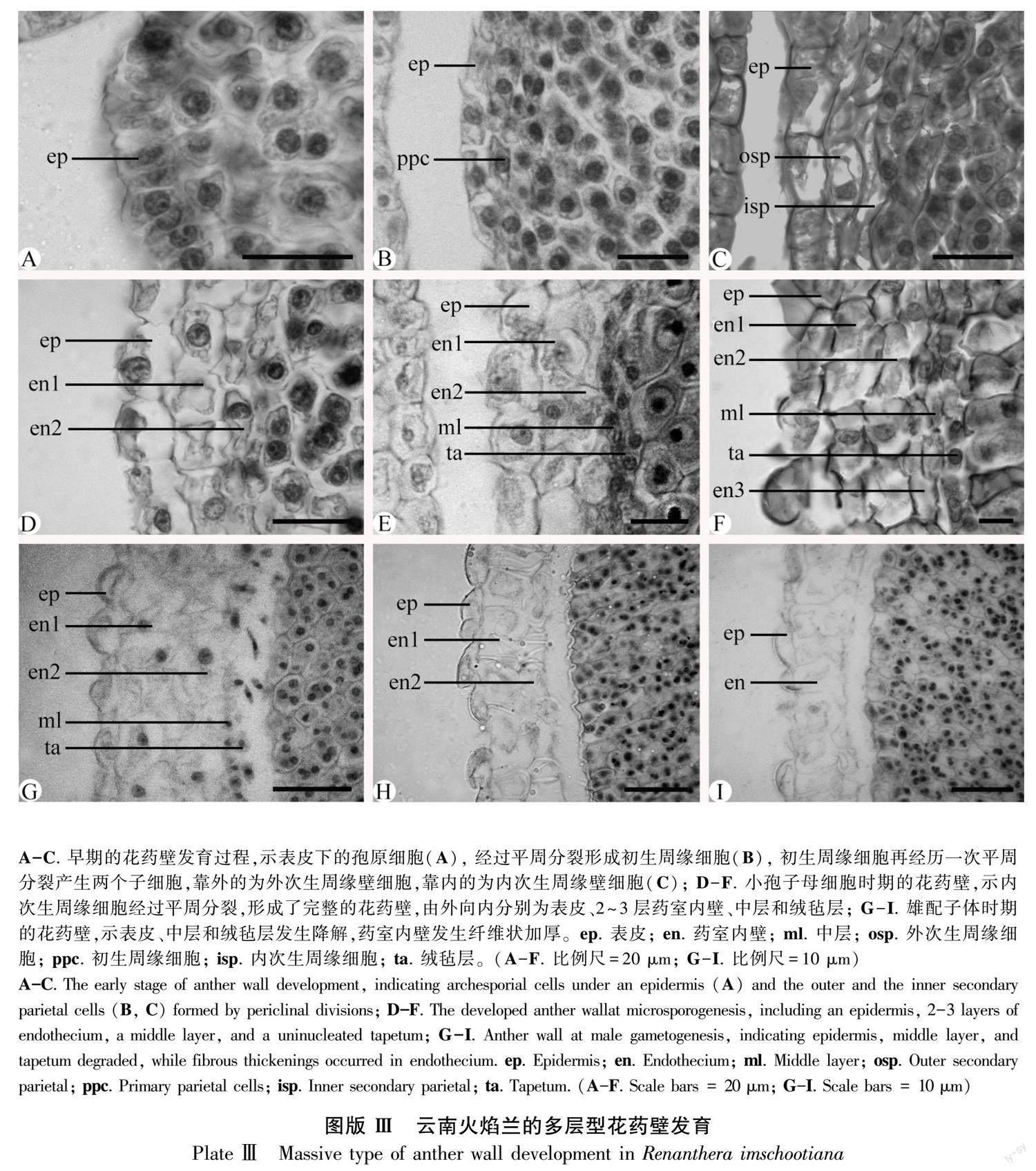
早期的花药原基由表皮下的一团染色较深的孢原细胞形成(圖版Ⅲ:A)。表皮下的孢原细胞分化为初生周缘壁细胞(图版Ⅲ:B)。初生周缘壁细胞通过2~3次平周分裂,相继发育为内、外次生周缘壁细胞(图版Ⅲ:C)。到小孢子母细胞时期,发育为完整的花药壁,由外至内分别为表皮、2~3层药室内壁、中层和绒毡层(图版Ⅲ:D-F)。因此,花药壁发育类型为“多层型”(massive tape)(Sood, 1989)。此时,表皮细胞和药室内壁细胞增大,呈方廓形;中层细长;绒毡层近圆球形、具单核、为腺质型(图版Ⅲ:E,F)。在小孢子母细胞减数分裂期间,中层和绒毡层出现降解,表皮细胞发生退化,呈不规则锯齿状(图版Ⅲ:G-I)。在雄配子体发育至2-细胞花粉时期,表皮逐步降解,有明显残迹,中层和绒毡层降解完全,出现明显空隙,2层药室内壁细胞出现明显环状加厚(图版Ⅲ:H,I),为药室开裂散粉提供了基础。
2.4 小孢子发生及雄配子体发育
首先,早期的花药原基表皮下包围着一团染色较深的孢原细胞,经数次有丝分裂后分化为排列紧密、核质比大的小孢子母细胞(图版Ⅳ:A)。小孢子母细胞的核质比增大,明显呈多边形,进入减数分裂Ⅰ阶段(图版Ⅳ:B),产生了没有子细胞壁的2个子核(图版Ⅳ:C)。随着发育的进行,2个子核同时进行下一步分裂,构成了保持在同一个细胞壁内、排列为正四面体或左右对称的4个子核(图版Ⅳ:D)。然后,4个子核间出现缢痕,同时各分化出子细胞壁,构成小孢子四分体,排列方式多为正四面体(图版Ⅳ:E),偶见左右对称型(图版Ⅳ:F),未见其他排列类型。在雄配子体发育阶段,小孢子四分体没有从胼胝质中释放出来,继续通过有丝分裂,形成一个居中的大营养细胞和一个靠边的小生殖细胞(图版Ⅳ:G)。最后,生殖细胞被吸入营养细胞内,构成了2-细胞型花粉(图版Ⅳ:H)。4个成熟花粉保持在同一胼胝质体内,形成了四合花粉,排列紧密且形成了坚硬的成熟花粉团。值得注意的是,本文选取约50朵花蕾作为研究材料,观察了100个花药室的发育,均未观察到花粉败育现象。
3 讨论与结论
本研究通过解剖镜和石蜡切片技术,从花形态的结构和花药发育的细胞组织切片水平,揭示了云南火焰兰的花形态特征和花药发育的胚胎学特征,证实了一对深裂花粉团的形成与每个花药室小孢子囊中央的一条不完全贯穿的不育隔膜组织的分化和降解有关,结合现有资料,分析了它们的分类学及保护生物学意义。
3.1 可用于火焰兰属物种鉴定和分类的花形态特征
首先,本研究支持前人观点,认为总状花序的着生位置、花色和唇瓣裂片特征是国产3种火焰兰属植物的物种鉴定依据(吉占和等,1999; 刘仲健等,2003; Chen et al., 2009; 金效华等,2019)。腋生总状花序在云南火焰兰和中华火焰兰出现,对生总状花序见于火焰兰。花色为橘红色系见于云南火焰兰和火焰兰,花色浅黄色系带紫红色斑点在中华火焰兰出现。唇瓣在3种中均为3裂,但中裂片和侧裂片特征在种间差异明显。中裂片上半部囊状球形见于中华火焰兰,其余2种均无囊状结构。侧裂片为直立的三角形且基部具一对平行高褶片在云南火焰兰出现;方形或近圆形见于火焰兰;卵圆形或披针形在中华火焰兰出现(刘仲健等,2003)。本研究观察到云南火焰兰的唇瓣中裂片和侧裂片基部的各有3枚白色肉质隆起的脊状物和褶片状胼胝体,在其余2种中均未有详细描述,值得进一步研究。
其次,本研究认为花粉团数目及形态、粘盘与粘盘柄特征在火焰兰属和种间变化明显,具有重要的分类学意义。火焰兰属所在的树兰亚科具有高度愈合的花粉团及花粉团附属物,包括花药帽、粘盘、粘盘柄和花粉团柄等(Dressler, 1993; Singer et al., 2008; Mosquera-Mosquera et al., 2019)。然而,关于这些花结构特征的研究在火焰兰属资料中很少见(吉占和等,1999; 刘仲健等,2003; Chen et al., 2009; 金效华等,2019)。本研究利用解剖镜观察了云南火焰兰的成熟花药结构,发现花药开裂时,露出2个不等大、深裂的花粉团,由近圆形且厚实的粘盘粘结,并且该特征在组织切片上已得到验证。显然,这与现有资料将火焰兰属描述为4个近等大的花粉团,基部有弹丝,粘盘柄稍长而宽、粘盘厚、近圆形的特征明显不同(吉占和等,1999)。同时,中华火焰兰作为新种发表时,虽没有详细描述它的花粉团数目和形态,但线描图记录了其具一对不等大的花粉团且粘盘为马鞍形(刘仲健等,2003)。火焰兰的花粉团形态和数目不详,粘盘柄被描述为中部屈膝状(吉占和等,1999),但其他2种的粘盘柄均为平直(刘仲健等,2003)。因此,花粉团形态和粘盘及粘盘柄等细节特征在火焰兰属具有重要的分类学意义,需要对其花形态特征进行准确的描述。
3.2 花药发育有助于确定指甲兰亚族多样的花粉团类型
长期以来,关于花粉团形态和数目在树兰亚科,尤其是指甲兰亚族不同类群的变化及其分类学意义颇有争议(Dressler, 1993; Freudestein et al., 2002; Chase et al., 2015)。这主要是因一些物种的花粉团特征(孔裂,或浅裂,或深裂)的位置较为隐蔽,往往藏在2个花药室内壁连接处,在解剖镜下也不容易观察清楚而造成的“误判”。例如,巾唇兰(Pennilabium yunnanensis)在解剖镜下为一对全缘花粉团,但组织切片却揭示其为一对细孔裂花粉团(李璐等,2020)。Freudenstein和Rasmussen(1996)通过石蜡切片技术,观察了树兰亚科具不同花粉团类型的10属代表类群的花药发育过程,揭示了2个、4个或8个花粉团是早期花药原基的小孢子囊内分化出不同数目(2,4)且纵向或横向排列的不育隔膜组织将其分割为次生小孢子囊,在小孢子发生和雄配子体阶段被吸收而降解出现空隙,把成熟花粉囊分割为不同数目的花粉团。类似研究在其他树兰亚科成员里已得到了证实。例如,具有一对孔裂或浅裂的花粉团是在花药早期分化出一条不完全贯穿小孢子囊的不育隔膜组织(An incomplete sterile septum)在花药成熟时因发生降解形成空隙而造成,见于美冠兰(Eulophia hormusjii)(Bhanwra et al., 2003)、纹瓣兰(Cymbidium aloifolium)(Kant et al., 2013)和大花万代兰(Vanda coerulea)(张锦等,2019)。竹叶兰(Arundina graminifolia)的8个花粉团是花药发育早期,每个药室的小孢子囊分化出的一对十字交叉的不育隔膜组织,在花药成熟时发生降解形成空隙,因将花粉囊分隔为成4枚棒状花粉团而形成的(谭庆琴等,2020)。本研究从形态结构和组织切片水平揭示了云南火焰兰的花粉团数目为2个,呈一对深裂花粉团,而非如资料所记载的火焰兰属具4个花粉团(吉占合等,1999)。因此,有必要通过组织切片观察兰科花药发育过程中不育隔膜组织分化的数目及其排列位置,以及它们在成熟花药阶段因发生降解而导致的花粉团形态和数目的关系(李璐等,2020)。这对于根据花粉团数目和形态特征来探讨万代兰族—指甲兰亚族的分类学和系统学具有重要启示(Freustein et al., 2002)。
3.3 兰科“多层型”花药壁发育模式与花粉团特征密切相关
本研究观察到云南火焰兰发育完整的花药壁有5~6层,属于“多层型”发育类型(massive type)(Sood, 1989),具有重要的分类系统学和传粉生态学意义。通常,被子植物的花药壁层数均为5层以下,可归为4种花药壁发育类型,包括基本型、单子叶型、双子叶型以及简化型(Johri et al., 1992; 胡适宜,2005)。然而,兰科的花药壁层数在亚科和属间变化较大,不仅有4层的单子叶型(monocotyledonous type),也有6~7层的多层型,值得收集更多资料分析其分类学意义。在拟兰亚科(Kocyan & Endress, 2001)、杓兰亚科(Swamy, 1949; Ghimire et al., 2020)和兰亚科(Sood, 1986; Sood, 1988; Kant & Goel, 2013)的一些物种里,花药壁层数多为4层,属于单子叶型。然而,在树兰亚科,花药壁层数既有4层的单子叶型,也有多达6~7层的多层型。例如,单子叶型花药壁见于原沼兰属 (Malaxis)(Sood, 1992)、石斛属(Dendrobium)(Gurudeva, 2016)以及云南巾唇蘭(李璐等,2020)。多层型花药壁首次命名于羊耳蒜属(Liparis)(Sood, 1989),后来陆续见于兰属(Cymbidium)(Kant et al., 2013)、苞舌兰(Spathoglottis plicata)(Sriyot et al., 2015)、美冠兰属(Bhanwra et al., 2003)等。其中,大花万代兰(Vanda coerulea)的花药壁超过7层,高达9层(张锦等,2019),这在现有资料里较为少见。关于多层型花药壁,除了兰科的树兰亚科外,在广义夹竹桃科(Apocynaceae s. l.)也有报道(Johri et al., 1992; 艾静等,2021),例如马铃果属(Voacanga)的花药壁普遍为8~9层,有的多达12~16层(Maheshwari, 1971)。因此,本研究认为多层型花药壁的出现与花粉团的形成在兰科和萝藦科分类系统与传粉生态适应性方面具有一定相关性(Endress, 2016),值得重视。
3.4 小孢子减数分裂的胞质分裂方式和小孢子四分体排列多样化的分类系统意义
本研究观察到云南火焰兰的小孢子母细胞经同时型胞质分裂形成正四面体和左右对称的小孢子四分体,接着通过有丝分裂形成2-细胞型四合花粉,构成坚固致密的花粉团,这与兰科现有研究结果一致(Johri et al., 1992)。在兰科的花药发育过程中,同时型胞质分裂方式比较普遍,见于大部分兰科类群,如杓兰亚科的杓兰属(Sood & Rao, 1986),树兰亚科的天麻(Gastrodia alata)(梁汉兴,1983)、苞舌兰(Sriyot et al., 2015)、巾唇兰(李璐等,2020)和竹叶兰(谭庆琴等,2020)等。相反,连续型胞质分裂方式较少,出现在兰亚科的眉兰属(Ophrys)(Aybeke, 2012)和绶草属(Spiranthes)(Kant et al., 2013),树兰亚科的大花万代兰(张锦等,2019)等。关于胞质分裂方式的系统演化意义,已有研究认为同时型胞质分裂较为常见,是被子植物的祖征,而连续型较为少见,属于衍征(Furness et al., 2002)。这在兰科所在的单子叶植物纲或天门冬目(Asparagales)均有类似的观点(Rudall et al., 1997; Furness & Rudall, 1999)。因此,本研究支持在兰科系统演化中,同时型胞质分裂方式较为常见,且属于祖征。值得一提的是,同时型胞质分裂方式也见于杓兰亚科(Swamy,1949; Sood & Rao, 1986; Ghimire et al., 2020)。
本研究觀察到云南火焰兰的小孢子四分体排列形式有2种,即正四面体和左右对称,这与大部分兰科的特征一致。现有资料报道了兰科的小孢子四分体排列方式多样,通常有2~3种,多达5种,包括正四面体、左右对称、十字交叉、“T”型和线型(胡适宜,1982; Johri et al., 1992)。小孢子四分体的排列方式基本与花药成熟时四合花粉的排列方式保持一致,这在具有花粉小块的兰亚科(Kant & Arabaci, 2010; Kant et al., 2013)和具坚固花粉团的树兰亚科(张锦等,2019; 谭庆琴等,2020; 李璐等,2020)的花药发育过程中得到证实。结合现有资料,本研究认为,尽管每种兰花的小孢子四分体排列形式多样,但正四面体和左右对称最常见,这也是兰科小孢子发生的普遍特征之一。
3.5 云南火焰兰花形态及花药发育的保护生物学意义
本研究发现在花药发育过程中,云南火焰兰的小孢子发生和雄配子体发育均未出现花粉败育现象。因此,前人提到过云南火焰兰野外居群个体数量少(李娟等,2018)的原因与花粉发育无关,但有必要从胚珠发育、大孢子发生、雌配子体形成以及双受精后的胚胎和种子发育等方面开展生殖生物学研究,为理解火焰兰属的繁殖生物学和濒危机制提供新证据。
致谢 感谢云南丰春坊生物科技有限公司的王晓云女士和徐志锋先生提供实验材料;玉溪市林业和草原局的吴建勇先生提供部分图片; 西南林业大学云南生物多样性研究院提供实验平台。
参考文献:
AI J, LI L, WANG YP, et al., 2021. Sporogenesis and gametogenesis in Chinese endemic plant of Dregea sinensis Hemsl. var. sinensis (Apocynaceae sensu lato) and their taxonomic insights [J]. Guihaia, 41(12): 2014-2023. [艾静, 李璐, 王艳萍, 等, 2021. 中国特有种苦绳(广义夹竹桃科)的大小孢子发生和雌雄配子体发育及其分类学意义 [J]. 广西植物, 41(12): 2014-2023.]
AYBEKE M, 2012. Anther wall and pollen development in Ophrys mammosa L. (Orchidaceae) [J]. Plant Syst Evol, 298(6): 1015-1023.
BHANWRA RK, VIJ SP, 2003. The development of anther and male gametophyte in Eulophia hormusjii Duthie (Orchidaceae) [J]. Orchid Soc Ind, 17(1/2): 87-91.
CAO WJ, LI HX, TANG YJ, et al., 2014. Karyotype analysis of four intragenus and intergenus hybridized combinations F1 generation of Renanthera [J]. Subtrop Plant Sci, 43(1): 19-23. [曹雯静, 李虹侠, 唐源江, 等, 2014. 4个火焰兰属内及属间杂交新组合F1代核型分析 [J]. 亚热带植物科学, 43(1): 19-23.]
CHASE MW, CAMERON KM, FREUDENSTEIN, et al., 2015. An updated classification of Orchidaceae [J]. Bot J Linn Soc, 177(2): 151-174.
CHEN XQ, JI ZH, LUO YB, 1999. Native orchids of China in colour [M]. Beijing: Science Press: 318. [陈心启, 吉占和, 罗毅波, 1999. 中国野生兰科植物彩色图版鉴 [M]. 北京: 科学出版社: 318.]
CHEN XQ, LIU ZJ, ZHU GH, et al., 2009. Flora of China: Vol. 25 [M]. Beijing: Science Press; St. Louis: Missouri Botanical Garden Press: 451-452.
DRESSLER RL, 1986. Features of pollinaria and orchid classification [J]. Lindleyana, 1(2): 125-130.
DRESSLER RL, 1993. Phylogeny and classification of the orchid family [M]. Cambridge: Cambridge University Press.
ENDRESS PK, 2016. Development and evolution of extreme synorganization in angiosperm flowers and diversity: a comparison of Apocynaceae and Orchidaceae [J]. Ann Bot, 117(5): 749-767.
FREUDENSTEIN JV, RASMUSSEN FN, 1996. Pollinium development and number in the Orchidaceae [J]. Am J Bot, 83(7): 813-824.
FREUDENSTEIN JV, HARRIS EM, RASMUSSEN FN, 2002. The evolution of anther morphology in orchids: incumbent anthers, superposed pollinia, and the vandoid complex [J]. Am J Bot, 89(11): 1747-1755.
FREUDENSTEIN JV, CHASE MW, 2015. Phylogenetic relationships in Epidendroideae (Orchidaceae), one of the great flowering plant radiations: progressive specialization and diversification [J]. Ann Bot, 115(4): 665-681.
FURNESS CA, RUDALL PJ, 1999. Microsporogenesis in monocotyledons [J]. Ann Bot, 84(4): 475-499.
FURNESS CA, RUDALL PJ, SAMPSON FB, 2002. Evolution of microsporogenesis in angiosperms [J]. Int J Plant Sci, 163(2): 235-260.
GHIMIRE B, SON S, KIM JH, et al., 2020. Gametophyte and embryonic ontogeny: understanding the reproductive calendar of Cypripedium japonicum Thunb. (Cypripedoideae, Orchidaceae), a lady s slipper orchid endemic to East Asia [J]. BMC Plant Biol, 20(1): 426.
GURUDEVA MR, 2016. Development of male and female gametophytes in Dendrobium ovatum (L.) Kraenzl. (Orchidaceae) [J]. J Orchid Soc Ind, 30: 75-87.
HU SY, 1982. Embryology of angiosperms [M]. Beijing: Higher Education Press. [胡适宜, 1982. 被子植物胚胎学 [M]. 北京: 高等教育出版社.]
HU SY, 2005. Reproductive biology of angiosperms [M]. Beijing: Higher Education Press: 29-92. [胡适宜, 2005. 被子植物生殖生物学 [M]. 北京: 高等教育出版社: 25-92.]
HUANG SQ, GUO YH, 2000. Research progress of pollination biology [J]. Chin Sci Bull, 45(3): 225-237. [黄双全, 郭友好, 2000. 传粉生物学的研究进展 [J]. 科学通报, 45(3): 225-237.]
JI ZH, CHEN XQ, LUO YB, et al., 1999. Flora Republicae Popularis Sinicae: Vol. 19 [M]. Beijing: Science Press: 291-294. [吉占和, 陈心启, 罗毅波, 等, 1999. 中国植物志: 第19卷 [M]. 北京: 科学出版社: 291-294.]
JIN XH, LI JW, YE DP, 2019. Atlas of native orchids in China [M]. Zhengzhou: Henan Science and Technology Press, 1036-1039. [金效華, 李剑武, 叶德平, 2019. 中国野生兰科植物原色图版鉴 [M]. 郑州: 河南科学技术出版社, 1036-1039.]
JOHRI BM, AMBEGAOKAR KB, SRIWASTAVA PS, 1992. Comparative embryology of angiosperms [M]. Springer Berlin Heidelberg.
KANT R, ARABACI G, 2010. Development of anther in relation to sectile pollinium in Zeuxine strateumatica (Lindl.) Schltr [J]. J Biol Life Sci, 1(1): 5-12.
KANT R, HOSSAIN MM, 2010. Development of pollinium in Malaxis muscifera (Lindl.) Kuntze [J]. Bangladesh J Botany, 39(2): 193-198.
KANT R, GOEL S, 2013. Development of anther in Goodyera repens (L.) R. Br.: A massulate orchid [J]. Vegetos, 26(1): 9-18.
KANT R, HOSSAIN MM, ATTRI LK, 2013. Pollinium development in Spiranthes sinensis (Pers.) Ames. and Cymbidium pendulum SW: A comparative study [J]. Bangladesh J Bot, 42(2): 307-314.
KOCYAN A, ENDRESS PK, 2001. Floral structure and development of Apostasia and Neuwiedia (Apostasioideae) and their relationships to other Orchidaceae [J]. Int J Plant Sci, 162(4): 847-867.
LI J, SHI M, L Y, et al., 2018. Discovery of an extremely endangered species Renanthera imschootiana and its population situation [J]. For Res, 31(3): 9-14. [李娟, 石明, 呂亚媚, 等, 2018. 极危植物云南火焰兰的发现及种群现状 [J]. 林业科学研究, 31(3): 9-14.]
LI L, 2023. Research progress on diversity of androecium development of Orchidaceae [J]. Guihaia, 43(8): 1537-1552. [李璐,2023. 兰科雄蕊发育多样性研究进展 [J]. 广西植物, 43(8): 1537-1552.]
LI L, ZHANG J, WANG YP, et al., 2020. Pollinia development of in a new species of Pennilabium yunnanensis (Orchidaceae) with systematic implications [J]. Bull Bot Res, 40(2): 161-171. [李璐, 张锦, 王艳萍, 等, 2020. 兰科新种巾唇兰的花粉团发育及分类学意义 [J]. 植物研究, 40(2): 161-171.]
LI ZL, 2006. Plant tissue production science [M]. Beijing: Peking University Press. [李正理, 2006. 植物组织制片学 [M]. 北京: 北京大学出版社.]
LIAN HX, 1981. Sporogenesis and the formation of male and female gametophyte of Gastrodiaelata [J]. J Integr Plant Biol, 3: 186-191. [梁汉兴, 1981. 天麻大小孢子的发生和雌雄配子体的形成 [J]. 植物学报, 3: 186-191.]
LIU ZJ, CHEN XQ, ZHANG JY, 2003. Renanthera sinica, A new species of Orchidaceae from Yunnan [J]. J Wuhan Bot Res, 21(1): 37-39. [刘仲健, 陈心启, 张建勇, 2003. 云南兰科一新种——中华火焰兰 [J]. 武汉植物学研究, 21(1): 37-39.]
LUO JP, WU KL, WENG SF, 2012. Research progress on Renanthera [J]. Guangdong Agric Sci, 39(10): 69-72. [罗剑飘, 吴坤林, 翁殊斐, 2012. 火焰兰属植物研究进展 [J]. 广东农业科学, 39(10): 69-72.]
MAHESHWARI DH, 1971. Embryology of Apocynaceae I. Plumiereae [J]. J Ind Bot Soc, (50): 74-85.
Ministry of Agriculture and Rural Affairs of State Forestry and grassland administration, 2021. List of National Key Protected Wild Plants in China [R]. Beijing: Ministry of Agriculture and Rural Affairs of State Forestry and Grassland Administration. [国家林业和草原局农业农村部, 2021. 国家重点保护野生植物名录 [R]. 北京: 国家林业和草原局农业农村部.]
MOSQUERA-MOSQUERA HR, VALENCIABARRERA RM, ACEDO C, et al., 2019. Variation and evolutionary transformation of some characters of the pollinarium and pistil in Epidendroideae(Orchidaceae) [J]. Plant Syst Evol, 305(5): 353-374.
PANSARIN ER, MACIEL AA, 2017. Evolution of pollination systems involving edible trichomes in orchids [J]. Aob Plants, 9(4): plx033.
PRIDGEON AM, CRIBB PJ, CHASE MW, et al., 2005. Genera Orchidacearum: Vol. 4. Epidendroideae(Part one) [M]. New York: Oxford University Press.
RAO AN, 1967. Flower and seed development in Arundina graminifolia [J]. Phytomorphology, (17): 291-300.
RUSDI NA, NAJWA A, 2020. Scanning electron microscopy analysis of early floral development in Renanthera bella J. J. Wood, an endemic orchid from Sabah [J]. Pertanika J Trop Agric Sci, 43(3): 377-389 .
SINGER RB, BARBARA G, CROSS H, et al., 2008. The use of orchid pollinia or pollinaria for taxonomic identification [J]. Selbyana, 29(1): 6-19.
SOOD SK, 1986. Gametogenesis, integuments initiation and embryogeny in three species of Habenaria ( Orchidaceae, Orchideae) [J]. Proc Ind Acad Sci (Plant Sci), 96(6): 487-494.
SOOD SK, RAO PRM, 1986. Gametophytes, embryogeny and pericarp of Microstylis wallichii Lindl. [J]. Bot Mag Tokyo, 99: 351-359.
SOOD SK, 1988. Development of gametophytes, embryogeny and pericarp in Goodyera repens (Orchidaceae, Neottieae) [J]. Proc Indian Acad Sci (Plant Sci), 98(2): 149-156.
SOOD SK, 1989. Embryology and systematic position of Liparis(Orchidaceae) [J]. Plant Sys Evol, 166: 1-9.
SOOD SK, 1992. Embryology of Malaxis saprophyta, with comments on the systematic position of Malaxis (Orchidaceae) [J]. Plant Syst Evol, 179(1/2): 95-105.
SRIYOT N, THAMMATHAWORN A, THEERAKULPISUT P, 2015. Embryology of Spathoglottis plicata Blume: A reinvestigation and additional data [J]. Trop Natural Hist, 15(2): 97-115.
SUN WB, 2021. List of Yunnan protected plant species with extremely small populations (2021) [M]. Kunming: Yunnan Science and Technology Press. [孙卫邦, 2021. 《云南省极小种群野生植物保护名录(2021版)》 [M]. 昆明: 云南科技出版社.]
SWAMY B, 1949. Embryological studies in the Orchidaceae. II. Embryogeny [J]. Am Midl Nat, 41(1): 184-201.
TAN QQ, LI L, ZHANG J, et al., 2020. Pollinia development in Arundina graminifolia (Orchidaceae) with taxonomic implications [J]. Guihaia, 40(1): 83-94. [谭庆琴, 李璐, 张锦, 等, 2020. 传统傣药竹叶兰的花粉团发育及分类学意义 [J]. 广西植物, 40(1): 83-94.]
WU ZY, 2003. Flora of Yunnan: Vol. 14 [M]. Beijing: Science Press: 737. [吴征镒, 2003. 云南植物志, 第14卷 [M]. 北京: 科学出版社: 737.]
YANG MS, 2013. China red list of higher plants—evaluation s report [M]. Beijing: Chinas Environmental Yearbook: 646. [杨明森, 2013. 中国生物多样性红色名录——高等植物卷 [M]. 北京: 中国环境年鉴社: 646.]
ZHANG HP, TAO ZB, TRUNSCHKE J, et al., 2022. Reproductive isolation among three nocturnal moth-pollinated sympatric Habenaria species (Orchidaceae) [J]. Fr Plant Sci, (13): 908852.
ZHANG J, ZHANG H, LI L, et al., 2019. Pollinia development in rare orchid of Vanda coerulea with its taxonomic implications [J]. Acta Bot Boreal-Occident Sin, 39(7): 1143-1153. [张锦, 张辉, 李璐, 等, 2019. 珍稀濒危植物大花万代兰的花粉团发育及其分类学意义 [J]. 西北植物学报, 39(7): 1143-1153.]
( 责任编辑 蒋巧媛 )
收稿日期: 2023-04-05
基金项目: 国家自然科学基金(32060049); 中国科学院西双版纳热带植物园园林园艺中心研究基金(E2ZK291B05)。
第一作者: 王乐骋(1998-),硕士研究生,研究方向为资源利用与植物保护,(E-mail)wangyuechengwyb@163.com。
* 通信作者: 李璐,博士,副研究員,研究方向为植物系统与进化、植物形态解剖学,(E-mail)lilu@swfu.edu.com。
