外周组织胶质细胞发育与功能的研究进展
2024-03-08余悦邓灏刘思忞戚智慧叶芷铭刘梦洁姚茂金
余悦 ,邓灏 ,刘思忞 ,戚智慧 ,叶芷铭 ,刘梦洁 ,姚茂金
1广州医科大学附属第一医院/广州呼吸健康研究院/呼吸疾病全国重点实验室 广州市,510120
2中山大学附属第一医院麻醉科 广州市,510080
胶质细胞发挥运输营养物质,回收神经递质,支持和修复神经元等作用。虽然胶质细胞不直接参与神经冲动的产生和神经电活动的传递,但它们对维持神经元周围局部微环境的稳定至关重要,为神经元发挥正常功能所依赖。研究发现胶质细胞参与调节突触可塑性,影响学习记忆等多种重要的脑功能。胶质细胞与众多神经系统疾病的发生发展有紧密的联系,例如帕金森病,阿兹海默病等。越来越多的研究表明,靶向胶质细胞为治疗神经系统疾病提供了新的策略。除大量研究证实中枢神经系统胶质细胞功能外,零星研究发现,外周肠胶质细胞可调节肠神经系统稳态、肠道蠕动、肠道干细胞稳态、形成大肠结肠迁移运动复合体[1],并通过与神经元和免疫细胞的互作促进肠自主神经可塑性进而调节肠免疫稳态。说明外周组织胶质细胞对器官发挥生理功能具有十分重要的作用,但其他外周组织的胶质细胞研究进展仍十分有限。因外周胶质细胞起源于外胚层,故本综述将主要针对神经外胚层起源的胶质细胞的起源进行简要回顾,并对参与其生理病理功能的细胞分子深入讨论。
1 中枢胶质细胞
中枢胶质细胞广泛分布于中枢神经系统(central nervous system,CNS),在CNS 中数量约是神经元的10 倍。CNS 胶质细胞主要包括星形胶质细胞、少突胶质细胞和穆勒胶质细胞。因小胶质细胞起源于中胚层[2],非神经外胚层起源,故在本综述中不做讨论。
1.1 起源
在第七天(embryonic day 7,E7),小鼠胚胎形成原肠胚并产生3 个胚层,至E8 发展为早期神经管。放射状胶质细胞来自CNS 的神经上皮细胞,并在脑室下区(subventricular zone,SVZ) 通过不对称分裂生成中间祖细胞(intermediate progenitar cell,IPCs),这些IPCs 能分化为神经元和不同胶质细胞[3]。星形胶质细胞起源于脑室区和SVZ 的祖细胞。E14 后,大部分放射状胶质细胞逐渐失去增殖能力。约E15,初步的星形胶质细胞在大脑皮层中开始增多,为后续的皮层提供了主要的星形胶质细胞来源[4]。在整个中枢神经系统中,少突胶质细胞前体细胞 (oligodendrocyte precursor cells,OPCs) 一般来自胚胎的早期心室区 (ventricular zone,VZ) 的神经上皮细胞,但放射状胶质细胞也可以产生OPCs。随着胚胎发育,OPCs 分化并形成多种少突胶质细胞[5-7](图1)。

图1 中枢胶质细胞发生
1.2 星形胶质细胞
星形胶质细胞是CNS 最常见的胶质细胞,有纤维状和原生质状两种。纤维状主要分布在CNS白质,原生质状主要在大脑灰质。这些细胞产生并释放多种神经营养因子,如GDNF、BDNF 和NGF。GDNF 支持多巴胺能神经元和其他神经元的存活,BDNF 增强突触可塑性并促进海马体神经发生,NGF 维持胆碱能神经元的密度并保护神经元。
星形胶质细胞在血脑屏障(blood-brain barrier,BBB) 中起到关键作用,与其它细胞共同维持BBB 完整性[8]。它们通过释放特定因子来维持血脑屏障的稳定性,确保有毒物质不能进入大脑,同时确保营养物质能进入大脑。但BBB 也限制了一些治疗药物进入大脑,影响治疗效果。
星形胶质细胞与多种神经退行性疾病关系密切。载脂蛋白E (apolipoprotein E,ApoE) 是一个关键蛋白,主要由星形胶质细胞合成,APOE4 基因突变与阿尔兹海默症风险增加相关[9-10]。蛛网膜下腔出血会损伤BBB,但星形胶质细胞可以帮助BBB 重建[11]。此外,星形胶质细胞在调节神经递质和保护神经元免受伤害中也起到重要作用,例如它们摄取并转化谷氨酸以供神经元再利用。星形胶质细胞的某些功能障碍与疾病(如癫痫) 有关[12]。此外,星形胶质细胞通过摄取和释放特定化合物来抵抗氧化应激。
1.3 少突胶质细胞
少突胶质细胞在CNS 中形成髓鞘结构,助力电信号传递并保护神经元。在髓鞘生成高峰,这些细胞需要保持高代谢活性以生成髓鞘所需的多层脂质膜[13],而这增加了其对氧化性物质的敏感性[12]。少突胶质细胞可为神经元提供能量,通过转运乳酸等能量底物。在成年小鼠中,它们也参与髓鞘的维护和再生。
肌萎缩侧索硬化症与少突胶质细胞损伤有紧密关联,这种疾病因神经元丧失而导致肌无力等症状。当少突胶质细胞受损并凋亡,会引起小胶质细胞的聚集和活化。实验研究显示,星形胶质细胞在髓鞘再生时具有反应性,并与髓鞘恢复过程相关[14-15]。活化的胶质细胞会释放促炎因子和活性氧,加重神经损伤,进一步影响少突胶质细胞,形成一个恶性循环导致神经元损伤[5]。
1.4 穆勒胶质细胞
穆勒胶质细胞是视网膜的主要胶质细胞,起源于室管膜细胞。它们构成视网膜的支持组织,通过释放神经营养因子、降解兴奋性毒素等,为视网膜提供保护、营养和维持外部环境稳态,还参与血管生成与视网膜血流调节。
斑马鱼视网膜研究揭示,视网膜损伤后,穆勒胶质细胞会表达生长因子等基因如HBEGFα和TNFα,刺激祖细胞形成与增殖。Ascl1a基因在损伤中起关键作用,它调控细胞分化和增殖,与lin28 和let-7 miRNA 有关。lin28 与干细胞更新有关;let-7 miRNA 与细胞分化相关。Ascl1a还影响Wnt 信号和穆勒胶质细胞的重编程。胰岛素瘤相关蛋白1 (Insm1a) 也受Ascl1a调控,它影响细胞重编程和祖细胞周期[16]。
但是,穆勒胶质细胞的过度增生会干扰视网膜稳态,引起视网膜功能异常[17]。病变视网膜中的穆勒胶质细胞功能失调可能导致谷氨酸代谢与离子稳态紊乱,进而引起视网膜水肿和神经元死亡[18]。此外,穆勒胶质细胞与小胶质细胞之间的互作可加重视网膜炎症反应,如在实验性青光眼中。因此,控制这两种细胞的互作可能是预防青光眼中神经节细胞凋亡的有效策略[19]。
2 外周组织胶质细胞
一般而言,外周组织胶质细胞主要包括施旺细胞、卫星胶质细胞和肠胶质细胞,但最新研究进展发现,肺部和心脏组织也存在胶质细胞。
2.1 起源
背根和腹根的部分施旺细胞起源于神经嵴来源的边界帽细胞(一类位于胚胎运动出口点和背根进入区域的祖细胞的短暂群体),大多数施旺细胞起源于神经嵴来源的施旺前体细胞。施旺前体细胞亦起源于神经嵴,维持了神经干细胞的部分多能性。施旺细胞和施旺前体细胞具有发育可塑性,可作为多种胶质和非胶质类型细胞的多能祖细胞,如成牙本质细胞、黑色素细胞、自主神经元、软骨细胞、神经内膜成纤维细胞、嗜铬细胞和肠胶质细胞[20]。成熟卫星胶质细胞亦来源于神经嵴细胞。
在肠神经系统发育过程中发现了两种类型的神经祖细胞: 巢蛋白(Nestin) 阳性细胞可产生神经元和胶质细胞;而巢蛋白阴性细胞仅产生神经元。形成肠神经系统的神经元和胶质细胞均起源于神经嵴细胞,这些细胞从神经轴背侧的迷走神经和骶区迁移而来。在脊椎动物中,大多数肠神经元和胶质细胞来源于一小群表达Sox10 的迷走神经嵴细胞,这些细胞侵入胚胎前肠,后称为肠内神经嵴细胞,在广泛增殖后定植于整个肠道。不断扩大的肠内神经嵴细胞群体的分化导致大量Sox10 神经元亚型和Sox10 胶质细胞的嵌合,这些细胞被组织成肌肠丛和粘膜下丛。
多能神经嵴细胞的发育潜能受其所在组织的微环境限制,具有各种中间表型的细胞伴随其所在组织发育,最终分化为胶质细胞[21]。研究人员最近发现的一种心脏胶质细胞——心脏连接胶质细胞(cardiac nexus glia) 正是由神经嵴细胞发育分化而成,随后迁移到心脏,并主要集中在心脏流入和流出道结构中。
2.2 施旺细胞
施旺细胞具有形成髓鞘和支持轴突的功能,可确保中枢神经系统和外周之间快速、准确的交流。施旺细胞分为成熟和未成熟两种。转录因子Sox10是施旺细胞谱系命运决定最关键的转录因子,其表达决定施旺细胞向髓鞘细胞分化。Notch 信号传导参与施旺细胞前体到未成熟施旺细胞的转变。在胚胎晚期和围产期神经中,持续的轴突信号决定施旺细胞前体分化为未成熟施旺细胞。未成熟的施旺细胞将选择大直径轴突与之形成1∶1 的共生关系,此时,未成熟的施旺细胞分化为前施旺细胞,并被自身基板包围。最终,来自轴突和细胞外基质的持续信号引导成熟髓鞘施旺细胞的形成,这是最常见的外周胶质细胞亚型[22]。
施旺细胞决定啮齿动物中周围神经的再生能力。周围神经损伤后,施旺细胞通常会由分化的髓鞘型转变为修复施旺细胞。周围神经损伤后的急性期变化涉及施旺细胞重编程、轴突变性和髓鞘碎片清除。此后,轴突再生,修复施旺细胞再次分化为髓鞘施旺细胞。有研究表明,将施旺细胞移植入脊髓损伤模型可起神经保护作用,促进轴突和髓鞘的再生,以改善损伤后功能,但其在体内与宿主细胞整合能力差。在一项Ⅰ期临床实验中,研究人员将施旺细胞移植入完全性胸段脊髓损伤的损伤中心后发现ASIA 分级有所改善,损伤水平下神经电生理检查可见活动增加[23]。
皮肤损伤修复过程中,施旺细胞会富集于瘢痕疙瘩(keloid)。研究发现瘢痕疙瘩施旺细胞中肿瘤坏死因子α 诱导蛋白6 (tumor necrosis factor alpha-induced protein 6,TNFAIP6) 和细胞通讯网络因子3 (cellular communication network factor 3,CCN3) 表达上调,以募集巨噬细胞清除髓鞘碎片中的病变并重塑细胞外基质(ECM) 以促进神经再生。TNFAIP6 参与影响M2 巨噬细胞(与纤维化和瘢痕形成有关) 极化,并抑制炎症发生,CCN3则是一种可促进巨噬细胞募集和分化为M2 表型的生长因子[24]。施旺细胞也与周围神经鞘瘤的形成密切相关。周围神经鞘瘤是外周神经源性肿瘤中最常见的类型。
有研究通过体内外实验发现施旺细胞可调控肺癌细胞的迁移和侵袭,并协助形成转移灶[25]。在癌细胞灶中,施旺细胞诱导癌细胞在其方向上形成突起并在癌细胞之间插层,导致癌细胞分散。施旺细胞与癌细胞接触时会表达神经细胞黏附分子1(neural cell adhesion molecule 1,NCAM1),并诱导其更长的丝状伪足形成,促进癌细胞向神经周围侵袭。NCAM1 编码的细胞黏附蛋白是免疫球蛋白超家族的一员,在神经系统的发育中发挥作用,其表达参与局灶粘附组装的抑制和癌细胞迁移的增加,且已被报道与多种癌症标本中神经周围浸润(perineural invade,PNI) 的存在相关[26](图2)。
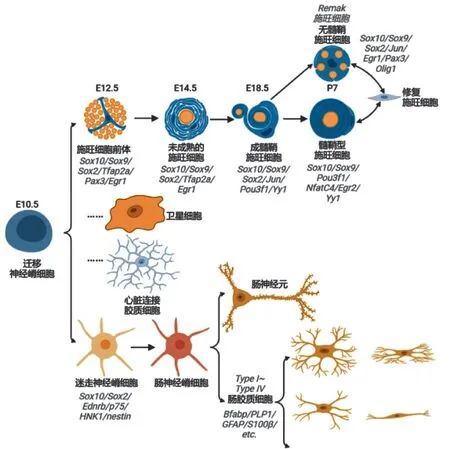
图2 外周胶质细胞起源与发育关键基因
2.3 卫星胶质细胞
卫星胶质细胞(satellite glial cells,SGC) 呈扁平状,围绕并覆盖在外周组织中的运动和感觉神经元胞体周围。研究认为,这种结构使得卫星胶质细胞能像血脑屏障一样调节外周神经节周围微环境,具有支持、保护和营养等功能。目前卫星胶质细胞的研究大多基于感觉神经节中的卫星胶质细胞,而对自主神经节中的卫星胶质细胞研究较少。此外,交感神经节内的卫星胶质细胞可支持交感神经元的生理功能,并通过释放胶质递质影响交感神经元的突触活动[27]。卫星细胞上也有着丰富的神经递质和其余神经活性物质的受体,能与其周围的神经细胞及其邻近的卫星胶质细胞密切相互作用。
当神经损伤时,SGC 能够上调脂质代谢相关基因以响应神经损伤。SGC 中富集的基因之一是脂肪酸合成酶(FASN) 基因,其控制内源性脂肪酸合成的特定步骤。脂质是细胞双层膜的重要组成部分。研究发现当FASN基因被敲除时,小鼠受损轴突长度变短,再生能力降低,但尚未发现会造成神经元形态缺陷。此外,FASN 在神经损伤后产生PPARα,FASN 依赖的PPARα 激活有助于促进轴突再生。过氧化物酶体增殖物激活受体(PPAR)具有结合脂质信号分子和转导来自代谢环境的适当信号以控制基因表达的独特能力,它有3 种亚型,PPARα 为其一。一些研究表明,PPARα 还能调控其靶基因APOE发挥作用[28],ApoE 参与激活水解脂肪的酶类,参与免疫调节及神经组织的再生。
研究发现卫星胶质细胞在进行单侧迷走神经切除术后的疼痛模型中被激活,表现为卫星胶质细胞趋于肥大,并产生大量溶酶体。损伤或炎症状态下,卫星胶质细胞激活导致促炎细胞因子释放,从而破坏神经元正常放电,导致神经性疼痛,该过程通过损伤后上调间隙连接蛋白,在感觉神经节神经元去极化中发挥作用。
2.4 肠胶质细胞
肠道中存在至少6 种主要类型的肠胶质细胞:包括与肌肠和黏膜下神经丛神经元胞体相关的胶质细胞(分别为ⅠMP型和ⅠSMP型),连接肌肠神经节的神经纤维束内的胶质细胞(Ⅱ型),与肌肠和黏膜下神经丛水平的神经纤维相关的神经节外胶质细胞(ⅢMP型或ⅢSMP型) 或肠黏膜(Ⅲmucosa型),以及平滑肌层中与神经纤维相关的胶质细胞 (Ⅳ型)[1]。
肠胶质细胞发育的关键因子之一是RET 信号通路。RET 是一种由神经嵴来源细胞(enteric neural crest-derived cells,ENCCs) 表达的酪氨酸激酶跨膜受体,在结合胶质源性神经营养因子GDNF后,ENCCs 会被磷酸化,并激活下游胞内磷脂酰肌醇激酶PI3K 通路,此通路可介导GDNF 诱导的神经嵴来源细胞存活、增殖和迁移。RET 表达需要转录因子Sox10 和Phox2B,两者的基因敲除表型均会导致全肠段无胶质细胞。内皮素受体B(endothelin receptor B,EDNRB) 信号转导也能促进ENCCs 的增殖,并抑制其分化为神经元[29]。肠胶质细胞的发育还受到骨形态发生蛋白如BMP4 通过ErbB3 受体传递信号的调控[30]。
肠神经系统是一种广泛的神经元网络,肠胶质细胞群是肠神经系统的重要组成部分,它们嵌入肠壁并控制肠道稳态。肠胶质细胞也通过神经递质受体信号监测多种形式的神经活动。肠胶质细胞表达中间丝胶质纤维酸性蛋白GFAP、静脉蛋白、钙结合蛋白S100、谷氨酰胺合成酶和胶质细胞表面抗原Ran-2。在单个胶质细胞受机械或化学刺激后,钙波在肠胶质细胞之间以波状方式传播,这一发现支持了功能性胶质网络存在的假说,该网络可能协调胶质细胞的协同活动。此外,研究报道肠胶质细胞对多种神经配体具有Ca2+瞬态反应,与星形胶质细胞相似,肠胶质细胞的兴奋性主要由细胞内钙离子浓度的短时间内迅速升高所介导,这表明它们存在神经配体-信号转导系统。
肠胶质细胞是连接肠道神经系统和免疫系统的纽带,并在维持黏膜屏障的完整性和肠道内稳态中发挥重要作用。多项在小鼠中的独立研究均证明肠道胶质细胞消融会导致严重的肠道炎症,这很可能是由于肠道神经丛结构的破坏或者血管上皮通透性的变化。这说明了肠胶质细胞参与维持肠道完整性。肠胶质细胞功能障碍早期病变的表现是黏膜和血管病变,这提示肠胶质细胞具有调节上皮和内皮细胞通透性的潜能。正在进行的研究表明,肠胶质细胞消融影响黏膜屏障功能不一定由肠神经元介导,而可能是肠胶质细胞和上皮细胞之间的直接相互作用[31]。证据表明,肠胶质细胞是胃肠道黏膜通透性和屏障功能的重要调节因子[32]。GFAP 的表达间接反映了肠胶质细胞的成熟度和胶质细胞对炎症刺激的反应性,如在克罗恩病 (Crohn’s disease,CD) 和溃疡性结肠炎 (ulcerative colitis,UC) 患者的肠道炎症区域均发现GFAP 表达增加。然而在非炎症区域,UC 中发现更高水平的GFAP表达,在CD 中却没有,这可能与CD 中持续炎症引发的胶质细胞损伤有关[33]。
综上,肠胶质细胞在特定情况下可驱动神经炎症,并通过与神经元和免疫细胞的互作增强神经可塑性、调节胃肠免疫稳态。因此靶向肠胶质细胞(如神经胶质递质释放、促进神经胶质增生的信号通路) 的疗法可能对常见胃肠道疾病的治疗具有潜在价值。此外,还有报道表明肠胶质细胞和癌症干细胞之间的相互作用的调节可能与十二指肠胃泌素瘤与结肠癌的发生相关[34-35]。
2.5 心脏胶质细胞
心脏连接胶质细胞是位于心脏流出道的分化成熟的细胞,可调节交感神经系统和副交感神经系统的活性,继而影响心脏的节律变化,如抑制室性心动过速和室颤。研究发现,敲除调控胶质细胞发育的METRN基因后,心脏连接胶质细胞的数量会显著减少,同时引发心脏节律异常。此外,研究人员在心脏发育早期应用甲硝唑和激光消融等方法清除心脏内连接胶质细胞后,观察到胚胎心脏中的神经发育速度加快,这表明心脏连接胶质细胞在发育早期起到抑制或延迟神经元神经支配的作用[36]。
心脏胶质细胞释放的S100β 是房颤消融术中急性心内神经损伤的标志。S100β 的释放促进了局部神经元的生长,同时降低了神经元的电活动。临床上接受导管消融的房颤患者可发现血浆中S100β水平升高[27]。
3 胶质细胞研究常用工具鼠
鉴于各类中枢和外周组织胶质细胞的重要功能,目前也已经开发了多品系工具小鼠用于对胶质细胞的研究,各品系工具小鼠靶向的基因。细胞种类及特点见表1。
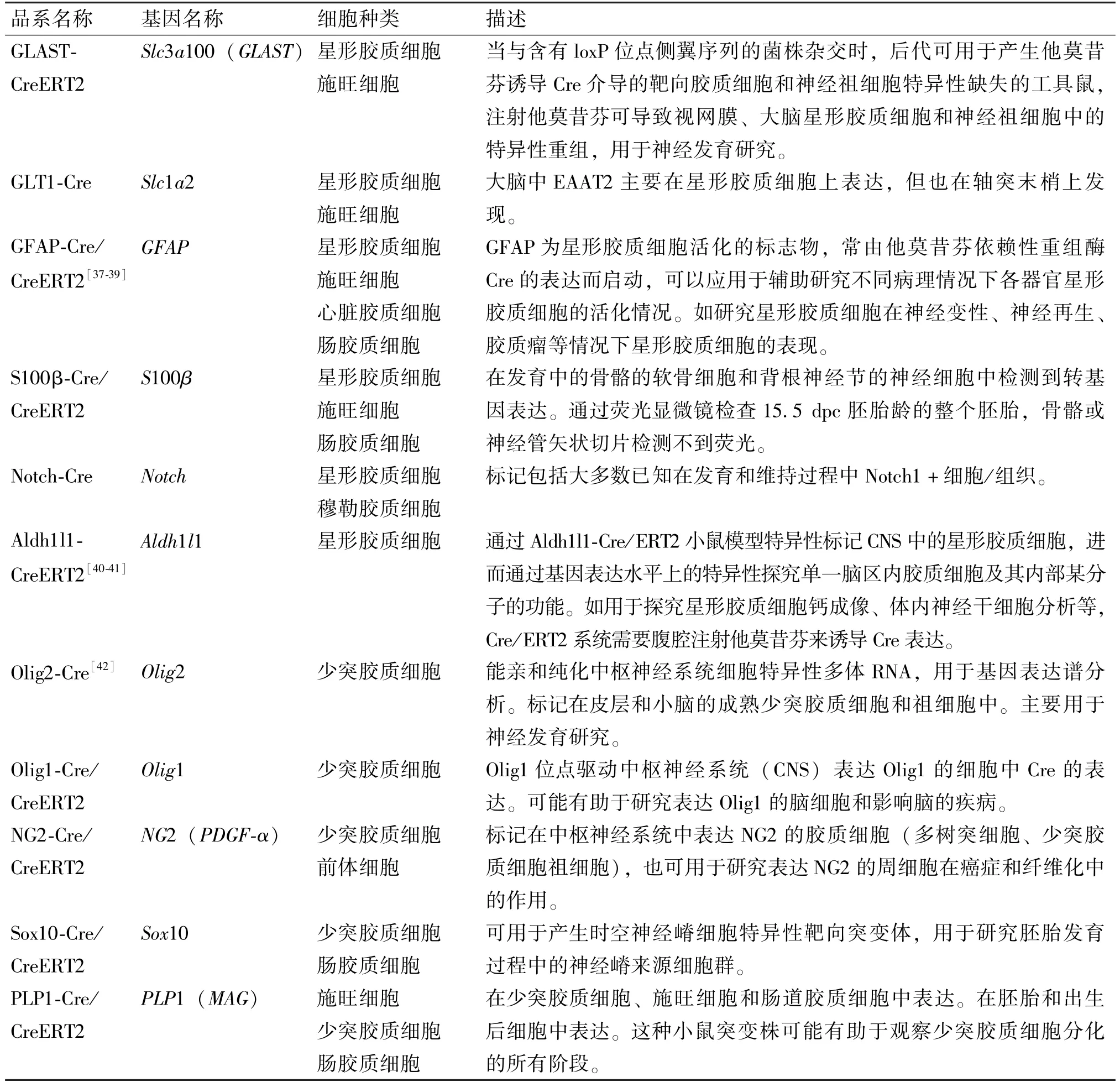
表1 胶质细胞研究常用工具鼠
4 展望
过去,胶质细胞一直被视为仅仅对神经元提供支持和营养的“配角”。但如今,随着研究的深入,这种认知已经发生了根本性的变化。胶质细胞不再是简单的背景填充,而是神经系统中功能丰富、种类繁多的关键角色。
在神经生物学中,胶质细胞的重要性已逐渐受到认可。不同类型的胶质细胞在起源、组织构成、形态和解剖学特征上均有所不同,它们在各自所处的器官中都发挥着独特且不可或缺的功能。但遗憾的是,目前学界对于胶质细胞在各种组织中的精确分布、功能以及交互关系尚未进行全面系统的描述。我们仍然需要进一步探索并鉴定。
中枢与外周的胶质细胞之间存在明显的差异,包括其分型、亚群分类以及在不同生理状态下的功能异质性。单细胞测序和空间转录组分析等前沿技术为我们提供了探索这些差异的有力工具。这些技术不仅能揭示同一来源的胶质细胞如何根据其所在器官的特性进行适应性演化,还能帮助我们解答外周胶质细胞的发育起源是否具有共性,以及其在表观遗传学层面的调控机制是否存在一致性。
当前,尽管我们已经对外周胶质细胞有了一定的了解,但关于它们的诸多疑问仍待解答。加速外周胶质细胞的研究不仅会揭示与众多疾病相关的新机制,亦为疾病治疗开辟新的路径,还将丰富我们对生命复杂性的理解。
