亚热带-温带过渡区秦岭落叶阔叶林幼苗存活机制
2024-03-08何念军朱文婷谢峰淋党海山
何念军 ,朱文婷 ,谢峰淋 ,李 涛 ,党海山
(1 陕西佛坪国家级自然保护区管理局,陕西汉中 723400;2 中国科学院 武汉植物园,武汉 430074;3 西藏大学 理学院,拉萨 850000;4 武汉市科籁恩科技发展有限公司,武汉 430074)
植物幼苗在生活史周期中是最脆弱和敏感的[1],其存活和生长与各种生物和非生物因素密切相关。对幼苗影响较大的因素包括林分条件、植物邻体、食草动物、微生物等生物因子[2],还有光照、水分、地形、土壤等环境因子[3]。植物幼苗的存活状态和生长速度影响着种群数量和群落组成,因此探究导致群落幼苗动态变化的因素对理解群落多样性维持及物种共存机制具有重要意义。
目前解释群落共存机制的2个重要理论分别为生态位分化和负密度制约效应。生态位分化理论认为不同的物种占有不同的生态位,影响物种共存的主要是生境因子,适宜某一生境的物种存活下来,反之则被淘汰[4-5]。负密度制约效应则认为同种物种由于资源竞争和有害生物侵害导致种群死亡率增加,从而为其他物种的生存提供资源和空间,促进物种共存[6-8]。因此探究幼苗存活是受到生物还是非生物因素的影响,即是探究物种共存的潜在机制。目前有许多研究证实负密度制约效应的存在,在温带和热带森林进行的大量研究发现,较高的同种邻体密度及距母树较近的幼苗死亡率较高[9-10]。也有研究以环境因子作为自变量对幼苗存活进行分析,证明生态位分化的重要性[11-12]。目前关于这2个理论的研究还没有定论。
亚热带-温带秦岭落叶阔叶林位于南北气候分界线处,拥有丰富而独特的植物资源,是进行植物多样性及物种共存机制研究的重要地点[13]。目前在此地进行的研究大多集中在物种组成、群落结构、物种多样性、幼苗组成及数量动态、种间关联等方面[14-16],而综合探讨生物和非生物等多种因素对幼苗存活影响的研究较少。文章以秦岭落叶阔叶林25 hm2动态监测样地为研究平台,对2015—2019年收集的植物数据和环境因子数据进行分析,探究生物及非生物因素对幼苗存活的影响,讨论群落物种共存的机制。旨在探明生物因素与非生物因素对幼苗存活的影响以及二者相对重要性的变化特征,并在群落水平上揭示不同苗龄阶段生物因素和非生物因素影响的差异,以期为亚热带-温带过渡区森林物种共存维持机制的研究提供理论依据。
1 材料和方法
1.1 研究区概况
研究区位于中国秦岭中段南坡的陕西省佛坪国家级自然保护区内,地理坐标为33°33′—33°46′N、107°41′—107°55′E,总面积约29 240 hm2。保护区的海拔在980~2 904 m 之间[17],地貌以山、沟、谷为主。研究区气候为北亚热带向暖温带过渡的山地暖温带气候,保护区内森林面积占总面积的98.98%,存在明显的山地森林小气候特征。年均温为11.5℃,春秋季均温在6~12℃之间,夏季均温在15℃以上,冬季均温低于0℃;夏秋季湿润,9月降水较多,冬季干燥少雨,年均降雨量980 mm[18]。植被和土壤均具有垂直地带性,土壤主要包括棕壤、黄棕壤、暗棕壤等[19]。
1.2 样地建设与调查
2015年参照CTFS(centre for tropical forest science)大样地建设标准[20],在保护区内建立1块25 hm2(500 m×500 m)的动态监测样地。以西南角为坐标原点,利用全站仪将样地分为625个20 m×20 m 的大样方,每个大样方又划分为16个5 m×5 m 的小样方,对每个小样方内所有胸径(DBH)≥1 cm 的木本植物进行普查,记录其种名、胸径、坐标及生长状态并挂牌,调查以“S形”顺序进行。
1.3 幼苗监测样方设置及调查
在秦岭样地中根据实际地形情况,设置135个种子雨收集器(图1),用于监测种子雨动态。以种子雨收集器为中心,在避开道路的另外3个方向分别设置1个1 m×1 m 的幼苗小样方,小样方与种子雨收集器的距离为2 m,分别编号为1-3,共405个小样方,每个种子雨收集器和其周围的3个小样方共同构成1个样站,用于长期监测幼苗动态(图2)。本研究将小样方内所有胸径<1 cm 的木本植物定义为幼苗,2015年7月对小样方内所有的幼苗进行清查编号并挂牌,记录幼苗的种名、叶片数、主干高度等指标。2015—2019 年,每年分别在6—7月和8—9月进行幼苗复查并记录更新情况,期间年累计监测到幼苗11 408 株,2019年仍存活的有4 477株。

图1 25 hm2 样地中种子雨收集器分布Fig.1 Distribution of seed traps in the 25 hm2 permanent plot

图2 幼苗监测样站示意图Fig.2 The components of a census station
研究将2018—2019 年记录的新生幼苗作为1~2年生幼苗;2016-2017年记录的新生幼苗作为3~4年生幼苗;将2015年记录的所有幼苗作为≥5年生幼苗。
1.4 环境因子测量
在建立秦岭大样地时使用全站仪获得676组高程数据,通过计算得到每个20 m×20 m 大样方内的坡度、坡向、海拔及凹凸度的平均值。海拔为每个样方4个顶点海拔的平均值[21];凹凸度是样方与其周围所有相邻样方平均值的差值;坡度是样方的4个顶点中随机选择3个,连接起来组成的4个不同平面与水平面夹角的平均值;坡向为上述4个不同平面与正北方向夹角的平均值[22]。
用五点取样法采集土壤样品[23],在20 m×20 m 的样方内,隔1个样方进行1次采样,共采集土壤样品313个。用直径为5 cm 的土钻钻取0-10 cm的表层土壤,再用四分法取100 g土样充分混匀后装入密封袋中。将土样风干、去除杂质后过筛,测量p H 值、容重、含水量、有机碳、全氮和全磷6种成分含量。
1.5 数据分析
将2015-2019年收集的幼苗数据进行整理,计算每株幼苗的大树邻体与幼苗邻体,幼苗邻体是指与目标幼苗在同一样方内的同种或异种幼苗的个体数;大树邻体是以目标幼苗所在的样站为圆心,半径为10 m 范围内的所有胸径≥1 cm 的同种或异种大树胸高断面积除以其与目标幼苗的距离之和。用Akaike's information criterion(AIC)对数据进行预分析,发现半径为10 m 时AIC值最小,即模型的拟合优度最高,所以选择以10 m 为半径计算大树变量,公式为:
式中:SBAi为第i棵大树的胸高断面积;D i为第i棵大树与目标幼苗的距离。
根据收集的土壤数据,对其中所有变量进行主成分分析(PCA),选取对幼苗存活解释度较大的前2位作为土壤因子的变量。按照计算得出的相关系数,选取前2个主成分,第1主成分中有机碳、全氮和全磷所占比重较高;第2主成分中含水量和p H所占比重较高。主成分分析的结果如表1所示。

表1 土壤变量的主成分分析Table 1 Soil variables loadings on the two PCAs
利用广义线性混合效应模型(generalized linear mixed-effect model,GLMMs)分析幼苗存活状态与各影响因素之间的关系。模型的因变量为幼苗存活状态:0(死亡)和1(存活);固定效应包括同种大树邻体(acon)和异种大树邻体(ahet)的胸高断面积、同种幼苗邻体(scon)、异种幼苗邻体(shet)、海拔(elevation)、凹凸度(convexity)、坡度(slope)、坡向(aspect)、土壤主成分1(comp.1)和土壤主成分2(comp.2);随机效应包括样站(site)和物种(Sp.)。本研究采用3种候选模型:(1)生物模型,包括随机效应(site+Sp.)和生物因子(scon+shet+acon+ahet);(2)非生物模型,包括随机效应和生境因子(elevation+convexity+slope+aspect+comp.1+comp.2);(3)全模型,包括随机效应、生物因子和生境因子(scon+shet+acon+ahet+elevation+convexity+slope+aspect+comp.1+comp.2)。分别在群落水平和不同年龄阶段(1~2年生,3~4年生,≥5年生)这2个水平检验各因素对幼苗存活影响的贡献。
利用AIC对模型进行筛选,AIC值最小的模型具有较好的拟合度,不同模型间AIC 的差值<2被认为具有同等拟合优度[24]。为了更好地比较各参数对幼苗存活的贡献,计算出各参数的比值比(也称优势比率),当优势比率大于1(95%的置信区间)时表示此参数与幼苗存活呈正相关,小于1(95%的置信区间)时则呈负相关。使用R-3.5.1中的“Ime4”和“ggplot2”等软件包进行数据分析和绘图。
2 结果与分析
2.1 群落水平幼苗存活影响因素分析
在3种模型中,群落水平上AIC 值最小即拟合效果最好的是生物模型(表2)。

表2 秦岭25 hm2 样地幼苗存活广义线性混合模型分析的AIC值Table 2 Akaike's information criterion(AIC)values of GLMMs of seedling survival in the 25 hm2 Qinling dynamic plot
通过模型分析(图3)可知,同种幼苗邻体密度和同种大树邻体与幼苗存活呈显著负相关(P<0.001),即同种幼苗邻体密度和同种大树胸高断面越大越不利于幼苗的存活。异种幼苗邻体密度与幼苗存活呈显著正相关(P<0.001),而异种大树邻体与幼苗存活呈正相关但是影响并不显著(P<0.32)。

图3 秦岭25 hm2 动态监测样地群落水平幼苗存活优势比率分析Fig.3 Odds ratio of seedling survival at community level in the 25 hm2 Qinling forest dynamics plot
2.2 不同苗龄幼苗存活影响因素分析
1~2年生幼苗和3~4年生幼苗均在进行全模型拟合时AIC值最低,拟合效果最好(表2)。由模型分析可知(图4),1~2年生幼苗的存活率与土壤因素1(优势比率为0.87,P<0.001)、同种大树邻体(优势比率为0.81,P<0.01)还有海拔(优势比率为0.81,P<0.01)均呈显著负相关。异种大树邻体(ahet)的胸高断面积与幼苗存活呈显著正相关(优势比率为1.31,P<0.001)。
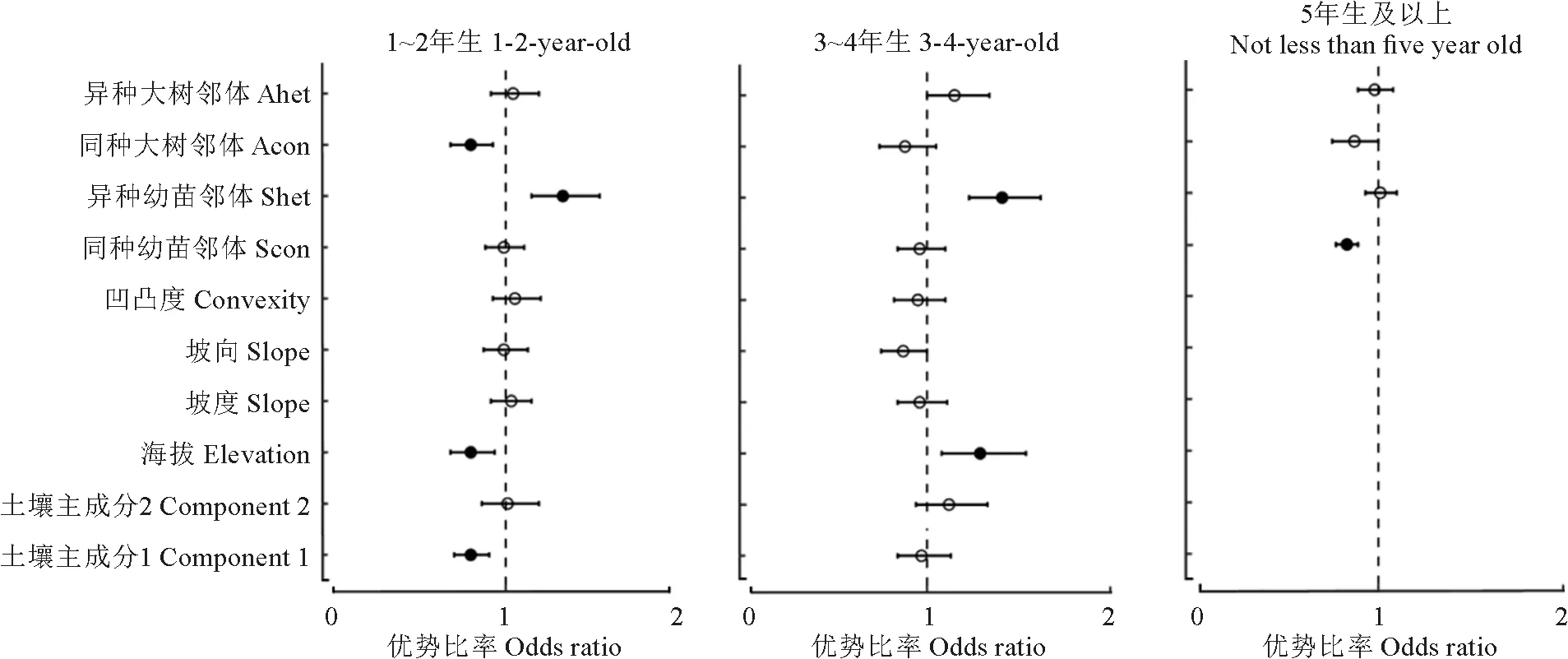
图4 秦岭25 hm2 动态监测样地不同年龄段幼苗存活优势比率分析Fig.4 Odds ratio of seedling survival at different ages in the 25 hm2 Qinling forest dynamics plot
其余因素对幼苗存活也有一定影响,但效应均不显著。3~4年生幼苗拟合的最优模型中(图4),异种幼苗邻体密度(优势比率为1.31,P<0.001)和海拔(优势比率为1.29,P<0.01)与幼苗存活呈显著正相关,其余所有因素对幼苗存活影响的效应均不显著。≥5年生的幼苗拟合效果最优的是生物模型(表2),其中同种幼苗邻体密度与幼苗存活呈显著负相关(优势比率为0.83,P<0.001),表明幼苗邻体密度较大时幼苗存活率降低。
3 讨论
3.1 群落水平幼苗存活影响因素分析
在群落水平上,生物因素与非生物因素对幼苗的存活均有一定的影响,但生物模型的拟合度要高于非生物模型,生物因素中的密度变量对幼苗存活影响显著,而生境因子的影响较小。图3中显示异种幼苗邻体密度与目标幼苗存活呈显著正相关,异种大树邻体的胸高断面积对幼苗存活也呈正相关但不显著,表明异种邻居的存在对目标幼苗的存活有利。同种幼苗邻体密度与同种大树邻体胸高断面积和目标幼苗的存活均呈显著负相关,表明同种之间具有明显的负密度制约效应。在不同气候区进行的研究结果与本研究均类似,在西双版纳热带森林中,同种和异种物种密度对幼苗的存活分别呈显著的负相关和正相关[25]。刘何铭等在天童山亚热带常绿阔叶林的研究结果显示,幼苗存活及幼苗的相对生长速率与异种大树邻体呈显著正相关[26]。在热带喀斯特森林的研究显示,同种和异种幼苗邻体密度与幼苗的存活均呈负相关,但同种幼苗邻体的负效应更加显著[27]。在凉水自然保护区的温带森林中,幼苗存活率随同种物种的增加而显著降低[28]。
同种邻近个体之间存在负密度制约可能是由于同种幼苗产生了资源竞争或者天敌在同种之间的扩散[29]。种子扩散距离受到各种因素限制,多数散落在母树附近,种子萌发后需要竞争有限的光照、水分、土壤等资源,激烈的竞争导致较高的幼苗死亡率,种群密度也随之下降[7,30]。宿主专一性天敌主要包括专一性食草动物和病原微生物等天敌,其会造成同种幼苗的大量死亡,所以被认为是影响物种分布和植物物种密度的主要因素[31]。有研究表明部分物种的母树附近会积累更多的专一性土壤病原菌,会使母树附近的幼苗大量死亡,且物种越丰富的种群会遭受更强的负密度制约[32]。负密度制约效应是维持物种共存的重要机制,同种幼苗的死亡为其他幼苗的存活提供空间和资源。异种邻居对幼苗的促进作用可以用“异种保护假说”来解释[33],本研究中异种幼苗邻体密度与目标幼苗存活呈显著正相关的结果,就符合此假说的预测。不同物种附近的食草动物或微生物等的种类不同,可以降低目标幼苗与其专一性天敌相遇的概率,从而提高目标幼苗的存活率[33]。
在群落水平上,非生物因素模型虽然不是最优模型,但检验后发现土壤主成分1与幼苗存活呈显著负相关。即土壤中有机碳、全氮和全磷含量较高不利于幼苗生长存活。而含水量和p H 值解释度较高的土壤主成分2与幼苗存活呈正相关,但效应不显著,可能是因为幼苗时期根系并不发达,只能利用土壤表层的水分,使得土壤表层的水分含量成为幼苗生长和存活的限制因素[34]。另外土壤的p H 值越大越有利于幼苗存活,秦岭大样地的p H 值在4.54~6.21之间,为酸性偏中性土壤,有利于幼苗的存活。
3.2 不同苗龄幼苗存活影响因素分析
幼苗苗龄不同,影响其存活的主要因子也不同。1~2年生和3~4年生幼苗拟合的最优模型都是全模型,但显著影响幼苗存活的因素有差异。1~2年生幼苗的存活与土壤主成分、海拔和同种幼苗邻体均呈显著负相关,与异种大树邻体呈显著正相关。3~4年生幼苗的存活与海拔和异种幼苗邻体呈显著正相关。5年生及以上幼苗与生物因子模型拟合较好,其幼苗存活只与同种幼苗邻体密度呈显著负相关(图4)。此结果与前人在八大公山和长白山等地的研究[35-36]有差异。其结果为幼苗存活时间越长,生物邻体的影响逐渐减小,而环境因子影响变大,其原因可能是幼苗在生长过程中,茎干逐渐木质化,对病菌感染和动物啃食的抗性增加[37],生物因素的影响逐渐减少,在生长过程中幼苗对环境中的光照、水分、土壤肥力等资源的需求上升,易受到环境胁迫[38]。本研究的结果显示随着幼苗年龄增长,显著影响幼苗存活的因素逐渐减少,可能是由于幼苗在早期经过环境过滤与密度制约后,将分布在较为适宜的环境中[27]。另外幼苗在生长中组织和器官逐渐发达,对环境的耐受力逐渐增加,受环境的影响会逐年减弱。
4 结论
影响幼苗存活的因子是多样的,在不同水平上分析时影响因子也有一定差异。在群落水平上,幼苗存活受生物因素的影响较大,与同种幼苗邻体和同种大树邻体呈显著负相关,与异种幼苗邻体呈显著正相关,表明物种幼苗受到强烈的负密度制约。在不同苗龄水平上,1~2年生和3~4年生幼苗的最优拟合模型是全模型,≥5年生幼苗的最优模型是生物模型。苗龄较小时受到生物因素和环境因素的共同影响,随着苗龄增大,环境因素的影响逐渐减小。综合来看,生物和非生物因素对幼苗存活均有显著影响,但生物因素的影响更加明显。秦岭大样地中物种幼苗共存的主要调控机制为负密度制约效应。
