梨形环棱螺11个地理种群的几何形态学分析
2024-03-05金武马学艳彭刚陈琬雯闻海波
金武,马学艳,彭刚,陈琬雯,闻海波
(1中国水产科学研究院淡水渔业研究中心/农业农村部稻渔综合种养生态重点实验室,江苏无锡 214081;2南京农业大学无锡渔业学院,江苏无锡 214128;3江苏省淡水水产研究所,南京 210017;4中美淡水贝类种质资源保护及利用国际联合实验室,江苏无锡 214081)
0 引言
梨形环棱螺(Bellamyapurificata)俗称螺蛳、豆田螺、石螺,隶属于腹足纲(Gastropoda),前鳃亚纲(Prosobranchia),田螺科(Viviparidae),环棱螺属(Bellamya),是环棱螺属个体最大的种[1]。该种分布广、产量大,用途多[2],日益得到研究者和养殖业者的关注[3]。梨形环棱螺作为全国47万hm2养殖河蟹重要的天然饵料之一,在长江禁渔后每年缺口达100~150 万t[4]。此外,石螺包括梨形环棱螺、方形环棱螺、铜锈环棱螺等作为柳州螺蛳粉的重要原料之一,在柳州地区每年缺口近3万t[5]。但对国内梨形环棱螺种质资源、分布状况以及不同群体之间的进化关系仍需进一步系统调查。
几何形态学测量法是关于形态定量比较的科学计算工具,使用数学方法描述或分析图像形状结构的方法,根据研究方式的不同分为轮廓法和地标点法[6-7],其中地标点法在水生动物形态的研究中得到广泛应用,其中以甲壳类和以鱼类为代表的脊椎动物的研究最为广泛[8-10]。国内对软体动物的报道限于单齿螺[11]、栉孔扇贝[12]、菲律宾蛤仔[13]等,本研究通过几何形态测量学(Geometric Morphometric)的方法,统计和分析不同地理种群的梨形环棱螺的形态学上的差异,以期为梨形环棱螺开展科学选育和增养殖工作提供理论依据。
1 材料与方法
1.1 样品与图像采集
梨形环棱螺样品采集自11个地点,具体采集点信息如表1。梨形环棱螺的鉴别参照文献[14]。每个地点的样品规格在2~5 g,再挑选壳顶和外壳无损伤的个体进行拍照。样品采集回实验室后,暂养于贝类循环水养殖系统中,待所有种群采集齐全后统一处理。样品放置于用白色细石英砂(Φ=0.2~0.5mm)填满的培养皿中。培养皿平面、手机、翻拍架经水平仪校验,以保证手机透镜中心轴与拍摄面垂直。镜头中心、培养皿中心、样品成像区域中心在同一纵轴。镜头与成像区域中的标本的距离为30 cm。光圈1.8,曝光时间1/144 s,拍摄角度为壳口观[15]。拍照时每个样品拍照3次,然后选择最佳的图片用于后续分析。图像保存格式为jpg。拍照由同一个人员完成。

表1 样品采集信息
1.2 数据整理与清洗
将照片中不同角度拍摄的照片统一用GIMP(Version 2.10.24)调整为壳顶在上,螺口在垂直轴线右侧。使用tpsUtil(Version 1.81)将获取的照片转换为.tps 格式文件。参照文献[16],利用tpsDig2(Version 2.31)在拍摄好的图片上建立地标标点,具体如图1 所示。获取相应地标点的x、y坐标值并建立数据文件。壳顶为第1 个地标点,顺时针编号,第1~24 个点为地标点。为了克服地标点不能涵盖整体的形态信息的缺点[7,17],在其中11~12、12~13、13~14、14~15、15~11、15~16地标点之间等距设置15个半地标点。用R[18]软件中的Geomorph 包的利用plotOutliers 检查地标点的超过上四分位的异常值情况并利用estimate.missing函数检查缺失值,缺失值检查采用薄板样条曲线(thin plate spline,TPS)法进行[19]。

图1 梨形环棱螺地标点设置
1.3 数据分析
用R[18]软件(version 4.1.1)中的Geomorph 包[20](version 4.0.0)的read.tps 函数读取创建的.tps 格式文件。利用gpagen 函数经获得地标点经普氏叠加的分布情况,对每个样品的地标点进行平移置中,计算出质心距离(Proj=T),求出平均型。根据不同地理种群划分为11 个组别,并利用geomorph.data.frame 合并坐标和群体组别。用gm.prcomp函数进行普氏形状变量的主成分分析(PCA),以确定形状变化的主轴,Geomorph包[20]中的plot.gm.prcomp 函数绘制前2 个主成分的散点图。
不同地理种群的平均外形通过mshape函数估计,再利用gpagen 函数获取不同地理种群地标点的平均值经gpagen普氏叠加的分布情况,并利用verbose选项输出普氏距离矩阵[21],并利用hclust绘制非加权配对算术平均法(unweighted pair group method with arithmetic mean,UPGMA) 聚 类 树 。 使 用plotRefToTarget 函数,生成方格变形图。用procD.lm函数检验不同地理种群对外壳形态的影响[21],置换检验采用残差随机化程序(Residual randomization in permutation procedures,RRPP)选项进行[22]。用Morpho包[23]中的relWarps绘制出几何形态的扭曲情况。
2 结果与分析
2.1 梨形环棱螺不同地理群体普氏叠印结果
排除了不同群体、不同个体之间的大小差异后,梨形环棱螺不同地理种群地标点普氏叠印的结果如图2,总体反映出梨形环棱螺11 个地理种群在不同地标点位置的差异。结果显示,螺壳形态的主要变异位置是壳顶及螺口上缘。如图3 所示,原始数据中仅将湖北潜江的第10个个体列入异常值,说明原始数据质量较好。
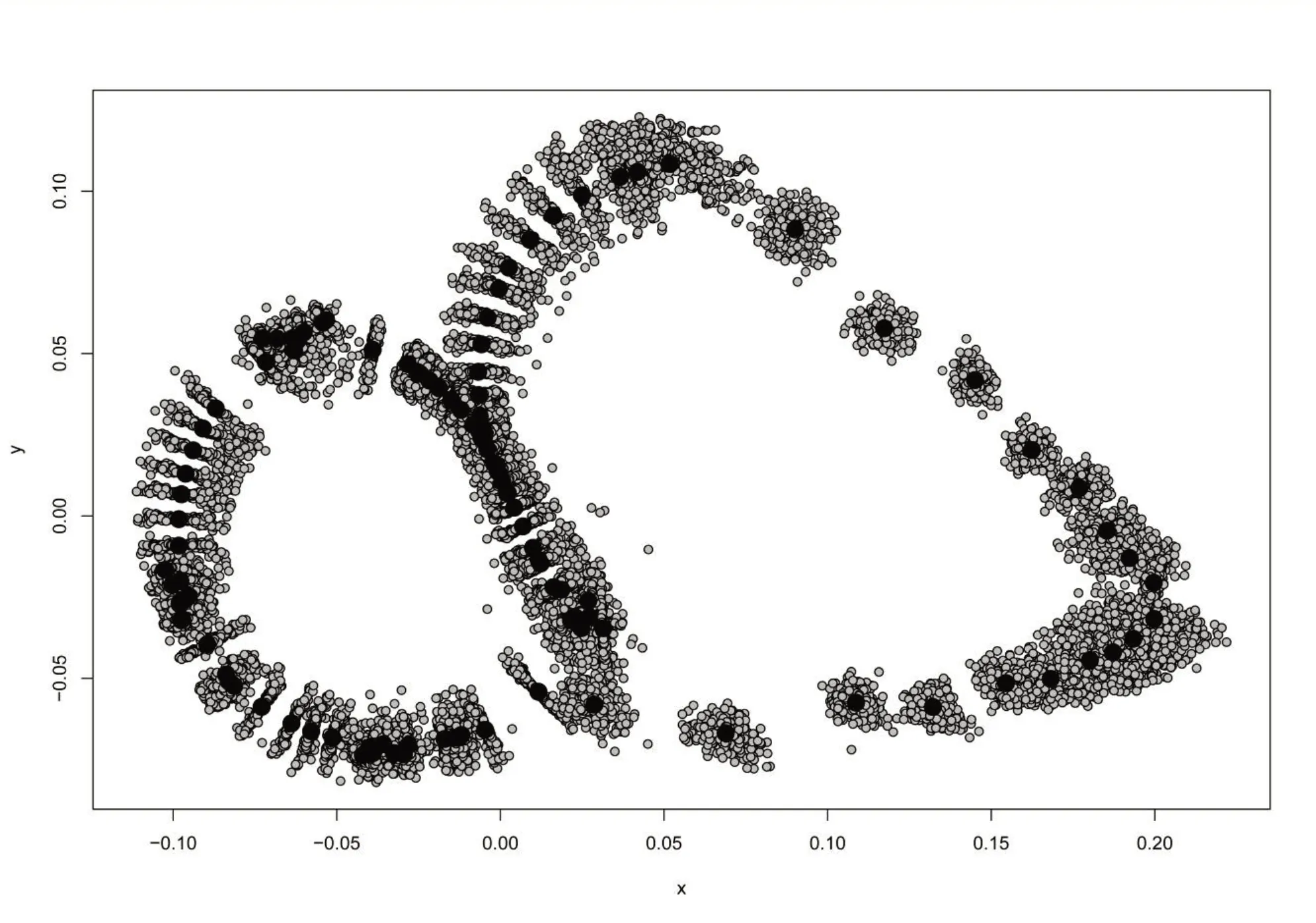
图2 梨形环棱螺所有样本地标点叠印
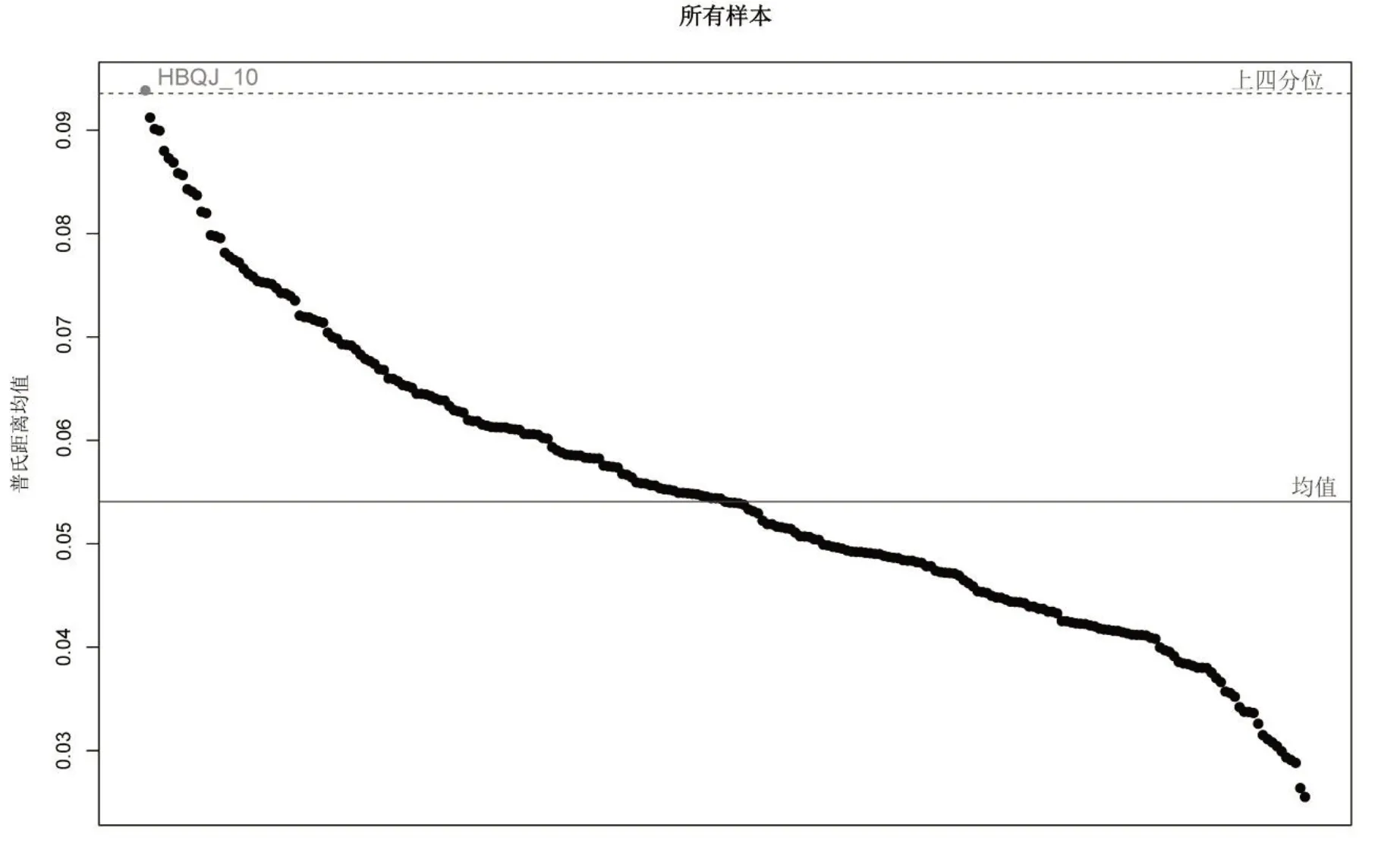
图3 异常值检查结果
2.2 梨形环棱螺不同地理群体主成分分析
在研究贝壳的形状多样性中,可以集中于形状变异的主要特征值,减少数据计算量[15]。梨形环棱螺不同地理种群壳口观的主成分的PC1 占37.53%,PC2 占16.06%,且前4 个主成分累计占比仅69.90%,说明梨形环棱螺的外形变异的集中性较差。第1主成分和第2主成分不能概括不同地理种群之间的形态学特征差异。如图4所示,不同群体不能形成较为集中的区域,仅有少数个别点重叠。
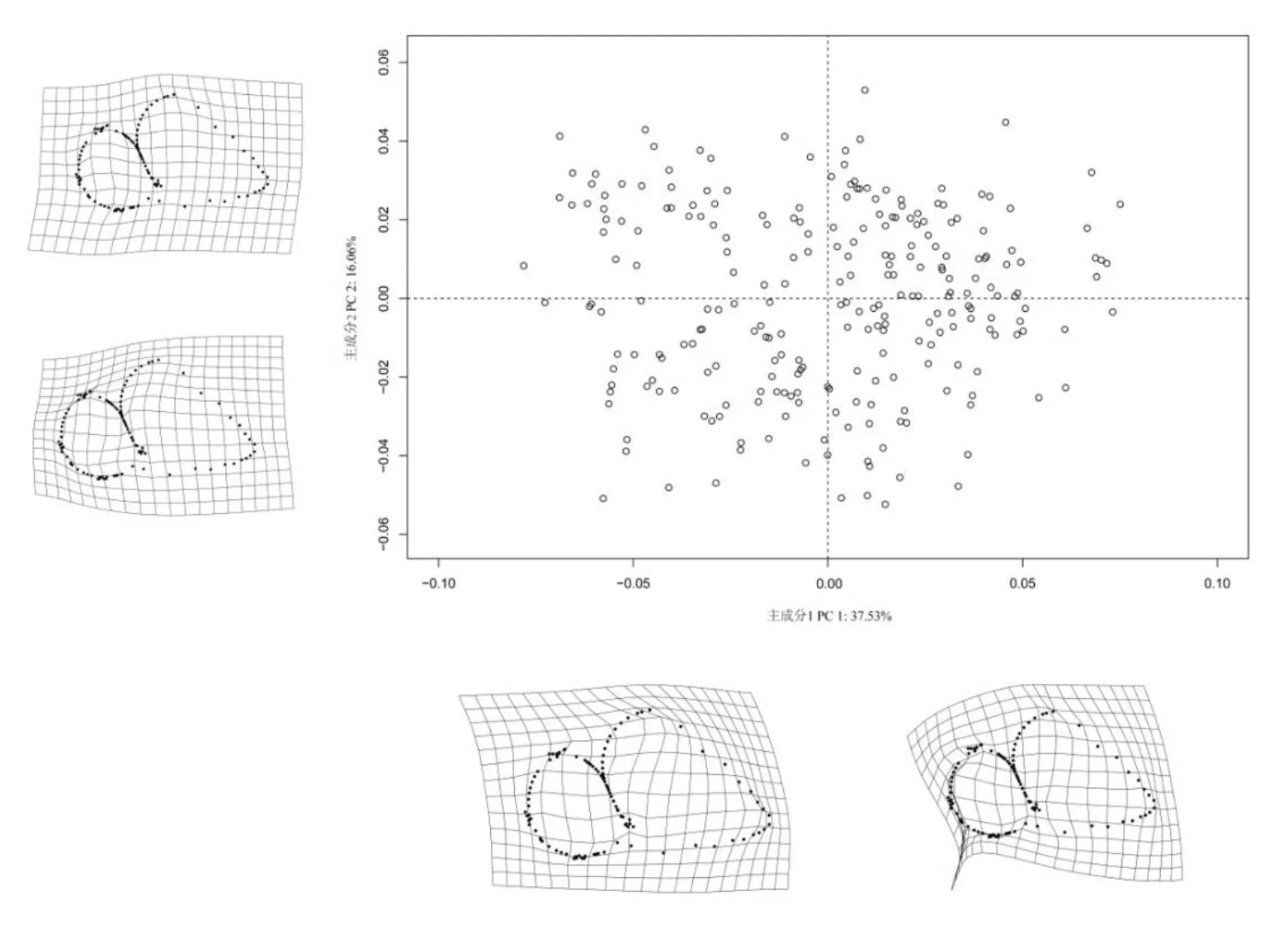
图4 249个个体普氏坐标主成分分析散点图
2.3 梨形环棱螺不同地理群体普氏距离方差分析
梨形环棱螺不同地理种群普氏距离的方差分析结果如表2所示,这11个地理种群的外形存在显著差异(p<0.01)。

表2 梨形环棱螺不同地理种群普氏距离方差分析
2.4 梨形环棱螺不同地理群体聚类
不同地理种群的梨形环棱螺聚类图如图5 所示,这11个群体主要可分为3支。浙江淳安群体和广东揭阳群体聚为一个分支,其余9 个群体被大致划分为长江流域和长江以南2支。
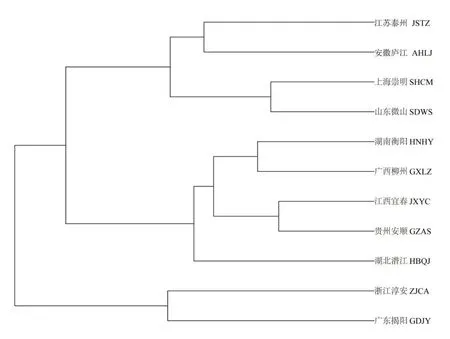
图5 梨形环棱螺11个群体聚类图
3 结论
梨形环棱螺11 个地理种群外部形态的主要变异位置是壳顶及螺口上缘。不同地理种群形态特征变异的集中性较差,主成分分析不能显著区别区分不同的群体。这11 个群体大致可以划分为长江流域群体和长江以南群体两大类,且不同群体的外形特征存在显著差异。这种外形差异可能与当地的生境有关。
4 讨论
4.1 几何形态学分析的优势
与传统的形态计量学相比,几何形态学克服了测量标准不一致、不同研究人员和不同研究对象导致的数据重复性差的缺点,同时将传统线性测量中大小和形状2个因素分开分析,具有显著的优势[12,24]。对螺类的几何形态学的研究甚至可以为开展交叉学科提供支持,深入探讨螺类对生境中的污染物的反应,如生态毒理学[25]。随着三维成像技术以及高速计算机的发展,对于某些生物类群的几何形态学的分析可能会比基于DNA 条形码技术更加简单、直观和高效[26],为水产动物表型组学研究提供支撑。不同地理种群之间的形态变异可以检验亚种分类的有效性,从而对分类上的一些疑难问题进行厘清。此外,在种上水平还可以探讨不同物种之间的形态分化及其异速进化现象[7]。通过结合分子标记对不同群体进行遗传距离分析并结合几何形态学分析的结果,还可以量化分析物种的生存环境,揭示这些生态环境的变化与形态变化之间的相关性,即生态相关性(ecological concordance)。
4.2 梨形环棱螺的几何形态学分析
除浙江淳安群体因样本量太少可能引起聚类误差外,本研究中11个群体可以大致划分为长江流域和长江以南区域两大地理区域,这与分子生物学的研究结果一致[27]。长江水系在很长一段时间与两岸湖泊相互连通,湖泊水系连通性好[28],以及长江流域频发的洪水可能促进了长江流域梨形环棱螺的扩散和种群之间的交流。梨形环棱螺11 个地理种群形态变异最大处为壳顶及螺口上缘,这与阿根廷濒危的Aylacostoma属的3 个种的几何形态学分析结果一致[16]。对非洲截头海蜷Cerithideadecollata的研究表明,贝壳形状的形态变化与遗传分化无关,与纬度梯度的分布有关,同时在地方一级的种群间又受到沿海地区生态的共同作用[29]。Aylacostoma属的3个种形态学差异与生境不同和是否暴露在水流中有关[30]。 对Littorinasaxatilis[31]、Pyrenaearia属[31]、Trophongeversianus[32]形态学变异的研究都表明与其所在的生境有关。梨形环棱螺种群间没有形成明显的地理格局分布,外部几何形态学的差异也可能与采样点所在的地理环境特点具有较强相关性,但与环境中具体哪些因子相关仍需进一步研究。
