水稻垩白形成机制的研究进展
2024-03-05程新杰施伟张梦龙岳红亮代金英胡蕾朱国永
程新杰,施伟,张梦龙,岳红亮,代金英,胡蕾,朱国永
(江苏沿海地区农业科学研究所,江苏盐城 224000)
0 引言
水稻产量一直是育种研究中的重心之一,随着人民生活水平的日益提高,消费者对稻米品质的要求也不断提高。近年来育种工作者对水稻品质的改良也逐步加以重视,但目前培育的水稻品种品质差强人意。因此,研究水稻品质遗传机理,改良稻米品质,培育出品质优良的水稻新品种或新组合,提高国内稻米的市场竞争力,已成为水稻科研工作者的新任务。稻米品质主要包括加工品质、外观品质、蒸煮食味品质以及营养品质。由于高垩白稻米在加工过程中整精米率低且极易破碎,另外在蒸煮后米饭蓬松中空、蒸煮食味品质较差。因此,垩白能够不仅影响稻米的外观品质,也会影响蒸煮食味品质。
水稻垩白是由于胚乳中淀粉体与蛋白质体松散包装形成的白色不透明部分。根据其在胚乳上或胚乳内的位置不同,可分为心白、背白和腹白[1]。水稻垩白作为一种不良性状,对稻米外观品质以及碾磨、食用和烹调品质均有不利影响[2]。目前主要以垩白度、垩白米率和垩白大小3 项指标对稻米外观品质进行评价分级[3]。垩白度是指垩白米的垩白面积总和占试样米粒面积总和的百分比。垩白米率则指的是有垩白的米粒占整个试样米粒总数的百分率。垩白大小是指垩白米粒平方时垩白面积占整个米粒投影面积的比率[4]。国家标准《GB/T 17891—2017 优质稻谷》中垩白度作为稻米分级的标准之一[5]。籼稻一级米要求垩白度不高于2.0%,二级米要求垩白度不高于5.0%,三级米要求垩白度不高于8.0%;粳稻一级米要求垩白度不高于2.0%,二级米要求垩白度不高于4.0%,三级米要求垩白度不高于6.0%。因此垩白度越低,稻米外观品质越好。
垩白作为衡量稻米外观品质的重要指标阐明其形成机理与遗传机制具有重要意义。本研究系统综述了水稻垩白形成受生理机制、遗传机制及环境因素的影响,基于这些研究结果,利用现今发展的分子育种等技术,完善对水稻垩白进行改良的育种思路,加速育种进程,为育成具有优良外观及食味品质的水稻新品种提高理论基础。
1 影响水稻形成的内在因素
1.1 生理机制
稻米品质主要是由灌浆过程中碳和氮代谢产生的淀粉与蛋白质储藏的化学组成决定的。其中,灌浆前期是垩白形成的关键时期,这一时期植株氮素营养水平对水稻品质形成影响较大。研究表明,追施氮素穗肥能够加快籽粒灌浆启动,增强灌浆初期强度,提高灌浆速率,延长活跃灌浆持续时间,促进籽粒灌浆,提高干物质积累[6],因此增施穗肥在一定程度上能够降低垩白度、垩白率[7]。
目前大部分研究均采用“源”、“库”、“流”学说解释垩白的形成,即叶片(源)进行光合作用生成的有机物质通过输导组织(流)运输到籽粒(库)进而形成胚乳。“源库”关系中,若同化产物供应不足,即“源”小“库”大时,胚乳总贮藏物质填充不充分,容易产生垩白。因此,抽穗期颖花所占叶面积越小,垩白粒率越大;抽穗期及灌浆前期剪叶处理即减“源”会提高垩白粒率,剪颖花处理即减“库”可以减少垩白[8]。隗华军等[9]以长江上游中籼迟熟优质稻组合为试验材料也有类似的发现,“源库比”较低的杂交稻组合中,源库比与垩白粒率呈负相关,而在“源库”比中等的杂交稻组合中没有发现相关性,因此能够通过调整“源库”比例来减少垩白粒率。
同时,水稻同化产物向胚乳运输的“流”也对垩白的产生具有影响。研究表明,低垩白品种的输导组织较高垩白品种更为发达,低垩白品种的籽粒背部维管束中导管数量较多,导管直径与筛管区宽带较大,珠心突起且糊粉层细胞层数较多,厚度较大[10]。灌浆速率也是“源”同化产物生产能力和“流”物质运输能力的综合体现,因此,灌浆速率的动态变化也会影响垩白的产生。
1.2 遗传机理
水稻垩白性状属于数量性状,受到多基因控制[11]。由于水稻胚乳细胞是通过双受精过程中两个极核与精核融合发育形成的,属于三倍体子代,在发育时又受母体基因型影响,因此胚乳垩白性状的遗传表现极为复杂,在研究过程中也存在诸多难点。
由于数量性状极易受环境的影响,很多群体试验是在特定环境下完成,导致遗传图谱中分子标记密度较低[12]。近年来,随着功能基因组学和分子标记的快速发展,水稻垩白性状的QTLs 定位也得到了帮助。目前,Gramene(https://archive.gramene.org/)网站上已记录82 个与垩白性状相关的QTLs 位点,分别分布在水稻的11条染色体上(除第4号染色体以外),其中第8号染色体上存在的QTLs位点数量最多,第10号染色体上QTLs 位点数目最少。这些QTLs 位点是不同研究团队分别通过F2 群体、重组自交系群体、单双倍体群体等不同的遗传群体进行定位得到的。其中,大部分QTLs 位点控制水稻垩白率,共有51 个,如:晁园等[13]利用川香29B/中国香稻群体定位的qPGWC-2、qPGWC-4、qPGWC-6-1、qPGWC-6-2、qPGWC-6-3等位点,还有周立军等[14]利用越光/Kasalath 群体定位的qPGWC-1a、qPGWC-1b、qPGWC-3、qPGWC-4等位点;而与垩白大小相关的QTLs位点有13个,如:Li等[15]利用日本晴/Kasalath 群体定位了qACE-1、qACE-3、qACE-5、qACE-6等位点;最后,与垩白度有关的QTLs位点则有21个,如:Wan等[16]利用Asominori/IR24群体定位了qDEC-1a、qDEC-1b、qDEC-2等位点。对这些QTLs位点功能进行分析,垩白性状的主效QTLs位点如表1所示。
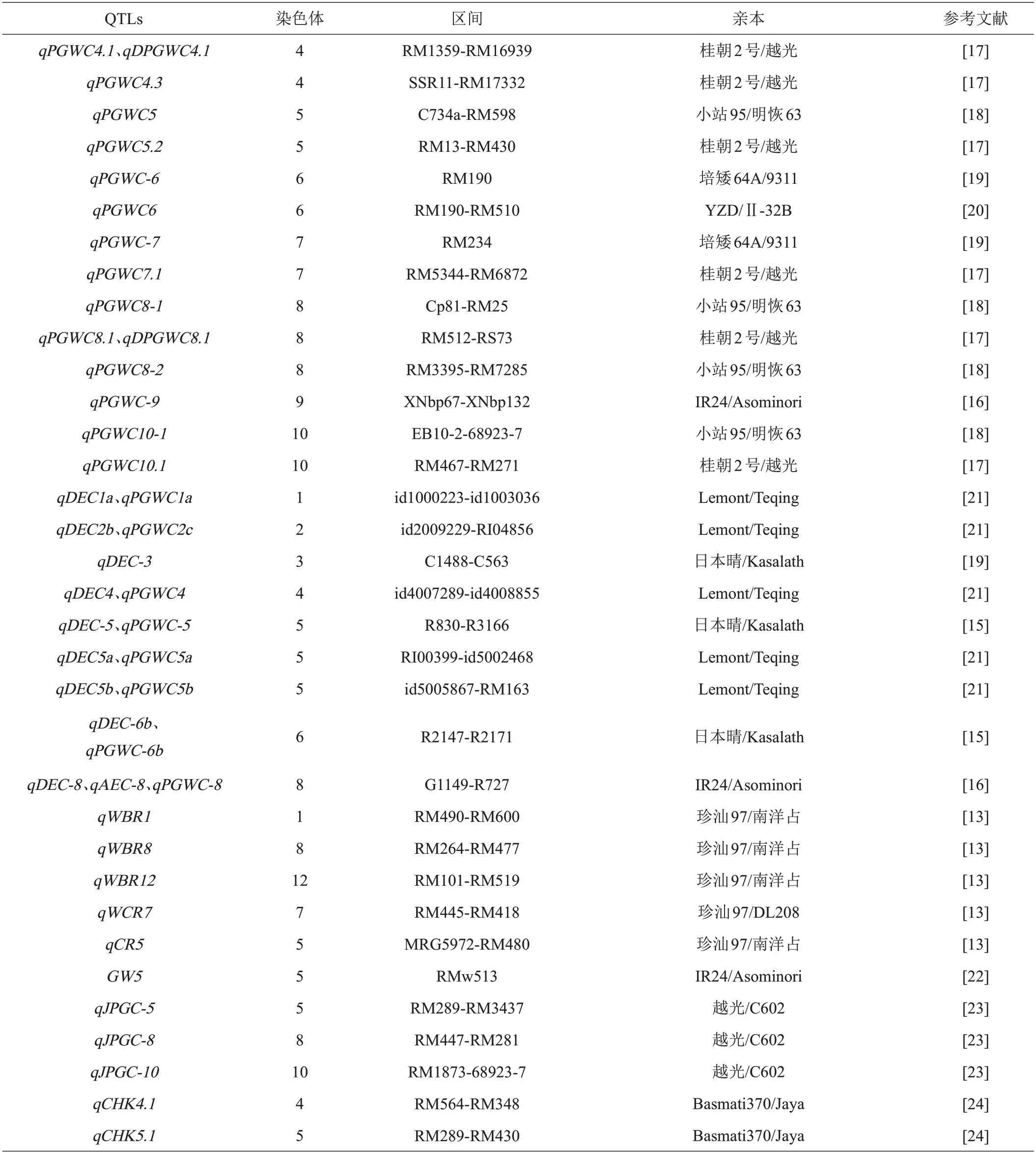
表1 水稻垩白性状主效QTLs
目前大部分研究人员还停留在垩白性状QTLs的初步定位上面,只有少量的完成了精细定位。如Zhou等[19]利用9311和培矮64S构建了染色体片段代换系群体,检测到两个控制水稻垩白率的QTL位点:qPGWC-6和qPGWC-7,分别位于水稻的第6 和第7 号染色体,其中qPGWC-7已精细定位至44 kb范围内。Guo等[25]则利用一个籼稻和粳稻的重组自交系衍生的染色体片段代换系群体,检测到一个位于第8 号染色体上的控制垩白率的QTL位点,并将其精细定位至140 kb范围内。
由于垩白性状受多种因素的影响,目前已克隆的水稻垩白性状相关基因数量较少,如表2所示,其中利用自然变异所克隆的垩白相关基因仅有Chalk5[26]、GW2[27]和GL7[28]。Li等[26]则利用两个回交群体,对第5号染色体上的一个微效QTL进行精细定位,区间约为16.8 kb,最后已将其分离克隆,并将该QTLs位点命名为Chalk5。

表2 已克隆的水稻垩白相关基因
这些已克隆的基因对水稻垩白性状的调控机理已有一定的研究。Chalk5编码液泡膜质子转运焦磷酸酶,在胚乳特异表达,可正调控水稻腹白率,影响水稻籽粒垩白的形成和精米率。GW2是一个控制粒宽和粒重的主效QTL 位点。GW2编码一个定位于2 号染色体上的环型E3 泛素连接酶,而GW2功能的缺失导致泛素无法转移到靶蛋白上,因而使得本应降解的底物不能被特异识别,进而激活颖花外壳细胞的分裂,从而增加颖花外壳的宽度,另一方面,间接地,灌浆速率也得到了提高,胚乳的大小随之也得到了增加,最终使得稻谷的宽度、粒重以及产量都得到了增加[27]。GL7位于水稻7 号染色体上,是一个控制粒长和粒宽的主效QTL。GL7位点上17.1-kb的串联重复,使得GL7表达水平上调,同时下调GL7邻近负调控因子的表达水平,从而使得水稻粒长增长并提高了稻米的外观品质[28]。
研究人员还通过T-DNA 插入、辐射诱变、化学诱变等技术获得了垩白突变体,进而构建遗传群体,克隆相关基因。利用上述方法已克隆的基因有SSⅢa[29]、OsPPDKB[30]、GIF1[31-32]、OsRab5a[33]、Flo2[34]和ms-h[35]。SS Ⅲa 定位于8 号染色体,是一个编码淀粉合成酶Ⅲa 的基因,主要参与直链淀粉B2-B4 的延伸[29]。OsPPDKB是一个位于5号染色体上,编码丙酮酸磷酸双激酶的基因,主要表达部位为胚乳和糊粉层[30]。GIF1则是一个位于4 号染色体上的编码细胞壁转化酶的基因,主要参与灌浆早期碳源分配过程[32]。OsRab5a位于12 号染色体上,编码小分子GTP 酶,对胚乳细胞器内膜形成和贮藏蛋白向蛋白体PBII 运输具有重要作用[33]。Flo2定位于4 号染色体,所编码的蛋白具有一个TRR结构域,该基因在籽粒灌浆过程中高表达,预测可能参与蛋白质互作[34]。ms-h位于9 号染色体上,编码UDP 葡萄糖磷酸酶,主要控制水稻的育性,但该基因的突变也会出现垩白表型[35]。这些基因的突变体大都表现为高垩白,在育种工作中利用难度较大,无法直接利用,但有助于科研人员研究垩白形成的遗传机理。
目前也有研究表明,植物激素同样参与了垩白形成相关的分子调控通路。Xie等[36]以‘湘早籼24’及其母本‘湘早籼11’和父本‘湘早籼7’为研究材料,并结合转录组数据分析,发现植物激素含量和多种激素信号在垩白颖果中存在显著差异,特别是ABA 和生长素,通过多种转录因子及其下游调控因子的相互作用,调控了水稻垩白的形成。
2 影响水稻形成的外部因素
2.1 环境因素
垩白的形成受多种环境因素的调控[36],如生育期间的温度、光照、水分、土壤类型等。高温与干旱的逆境胁迫下,会强烈促进稻米垩白的形成。灌浆期是籽粒形成的关键时期,有研究表明,水稻授粉至灌浆早期,水稻对高温反应最为敏感,若在这一时期进行短暂的高温处理,会导致稻米灌浆充实不佳,使得垩白明显增加,且增长幅度大于灌浆期进行高温处理[37]。同时,也有研究发现,夜间高温更容易导致垩白的产生,这主要是通过植物激素增加垩白,可以通过外源施用油菜素内酯、维生素C、维生素E和茉莉酸甲酯部分消除高温引起的垩白增加[38]。
随着全球温室效应的加剧,育种科研人员逐渐关注大气二氧化碳和臭氧的浓度对稻米品质的影响,并进一步进行研究。研究发现,二氧化碳浓度的提高会增加水稻品种的垩白粒率和垩白度;当大气二氧化碳浓度与冠层温度同时提高时,白粒率和垩白度的提高幅度会加大,二者对于垩白的增加具有累加效应,可以通过增施氮肥与磷肥减少垩白,改善外观品质[39-40]。在臭氧处理下,水稻叶片养分供应受到影响,短链支链淀粉含量增加,同时垩白程度也表现出增加[41]。也有研究表明,在大气二氧化碳和臭氧浓度同时增加时,水稻垩白度受到的影响甚微[42]。
2.2 栽培条件因素
栽培学家根据多年田间试验,发现播期、栽插密度、肥料施用等栽培措施对稻米品质均有不同程度的影响[43]。
播期影响稻米品质的主要因素是温度,且灌浆期温度对稻米品质的影响极大。研究表明,对于早熟和中熟品种,要适当推迟播期,对于晚熟品种,推迟播期对稻米品质则会产生负面影响,总体而言,适宜的播期有利于改善稻米的外观品质和蒸煮食味品质[44]。
栽插密度不仅能够影响产量,对稻米的加工和外观品质也有着较大的影响。适宜的栽插密度可避免田间植株生长郁蔽或稀植栽培下水稻次生分蘖增加导致稻米垩白粒率或垩白度增加,同样有利于提高稻米品质[45]。
稻米外观品质受种植方式的影响较大[46],一般情况下,直播稻的外观品质较优,手栽与机插稻米的外观品质优劣则取决于稻米自身品质,而钵苗机插和摆栽稻米的垩白粒率与垩白度均有一定程度的提升,使得外观品质降低。
肥料的施加对于稻米品质有着重要影响。氮肥作为影响稻米品质的重要因素,不同地区,适宜的基蘖肥与穗肥比率,能够降低垩白利率,提高稻米的加工与营养品质[47]。磷、钾肥作为水稻生长的关键肥料,其施加量对于稻米的外观品质也有着较大的影响。黄丽芬等[48]、王伟妮等[49]发现,施用磷肥能显著降低垩白度和垩白大小;但龚金龙等[50]则发现施磷过量会导致垩白粒率提高,而钾肥施用量的增加则会使得垩白度与垩白粒率先下降后上升。由于氮、磷、钾肥间存在互作效应,因此三者配施能够有效降低稻米垩白度[49]。有机肥的施用则能够降低垩白粒率和垩白度[51]。
3 水稻垩白的遗传改良
由于田间垩白性状测量较为麻烦,而传统水稻遗传育种中,对于垩白性状只进行表型选择,因此导致垩白性状的改良效率较低,这也是目前优质稻米育种工作中遇到的重点难题。另外,水稻垩白是由多个QTLs控制的数量性状,且环境对垩白性状具有极大的影响[52]。随着现代分子生物学的发展,育种人员对于垩白性状改良的方法也在不断的进步。对于已稳定的QTLs 位点可进行进一步的研究,对其进行精细定位、分离和克隆,解析其调控垩白性状的分子机理,这能够为后续低垩白优质稻米遗传育种提供理论基础。也能够利用分子标记辅助选择育种,聚合有利的等位基因位点,同时去除增加垩白性状的等位基因位点,能够更快速的选育出优良品种,降低时间成本。
但传统育种存在耗时长,改良效率低等问题。因此,利用目前已精细定位的QTLs 位点和控制垩白的QTLs基因,不仅可以进行分子标记辅助育种或基因聚合。同时还能够使用转基因技术改良水稻的垩白品质。目前垩白性状的相关基因的分离、克隆大部分来源于水稻突变体的研究成果。但这些突变体大都表现出垩白面积大或垩白率高的表型,在大田育种过程中,这些基因的利用难度较大。但这些基因都能作为稻米品质遗传改良的资源,为水稻的品质改良提供指导作用。近年来,随着基因工程的快速发展,可通过RNAi等转基因技术抑制籼稻品种中mRNA的表达,改良籼稻的垩白性状,进而提高稻米品质。例如,由于目前转基因技术在应用中还存在一定争议,这些转基因品种在实际推广种植中较难应用。
4 展望
垩白作为水稻品质的重要性状之一,且对稻米的外观品质、加工品质、蒸煮食味品质和营养品质均具有重要的作用。因而,水稻的垩白品质能够衡量稻米的品质和市场价格。但是垩白性状作为一个多位点控制的数量性状,存在多个QTLs 控制垩白性状,同时,环境与栽培因素对垩白性状也具有较大的影响作用,垩白形成涉及的“源库流”理论,在水稻生长过程中,若三者中任何一个发生变化或其关系不稳定,都能导致稻米产生或增加垩白。因此,垩白性状的形成是一个复杂的过程,对其进行改良也是极为艰难。
当前快速发展的功能基因组学和生物学分子技术,对于人们研究控制垩白性状的相关基因与QTLs位点的研究具有极大的促进作用。这也加快了垩白性状的相关基因与QTLs 位点的挖掘与定位,同时有助于后续垩白性状的遗传机理和分子调控网络的研究。这也为分子标记辅助选择、转基因技术在育种上的应用提供了理论基础。未来的育种工作,可在传统的杂交、回交等育种方式的基础上,辅以全基因组选择育种、分子标记辅助选择育种等手段,实现垩白相关基因和QTLs 位点的转移与聚合,能够缩短水稻垩白性状改良的时间,更为高效地实现育种目标。另外,还可运用基因工程等手段,将远缘物种的优良基因转入现有水稻品种中,快速定点改良水稻的垩白性状。但转基因技术在育种工作中仍旧存在争议,在实际育种工作中还需进一步探讨。
