原核表达的褐黄血蜱唾液腺蛋白和铁蛋白1的免疫保护效果评价
2024-03-01岳婵娟范雪阳刘颂蕊杨光友
陈 玲,陈 浩,岳婵娟,马 锐,范雪阳,刘颂蕊*,杨光友*
(1.成都大熊猫繁育研究基地,四川省濒危野生动物保护生物学重点实验室,成都 610081;2.四川农业大学动物医学院,成都 611130)
蜱是一种常见的动物体表寄生虫,也是多种动物疾病的传播媒介,据估计在全球范围内,蜱和蜱传疾病每年会造成220亿~300亿美元的损失[1]。褐黄血蜱(Haemaphysalisflava)属于硬蜱科血蜱属,可寄生在犬、猫、山羊、刺猬、野猪以及鸟类体表[2-5],同时它也是危害野生大熊猫的优势蜱种[6]。该蜱在我国广泛分布于河南、湖南、湖北、四川、江苏等地区[7-8]。据报道褐黄血蜱能够携带立克次体(Rickettsiarickettsii)[9]、贝氏柯克斯体(Coxiellaburnetii)[10]、埃氏疏螺旋体(Borreliaafzelii)[5]和无形体(Anaplasma)[11]等诸多病原体,引起宿主或人类感染多种疾病,对动物和人类健康造成极大威胁。化学防控是控制蜱和蜱传疾病有效方法,但大量使用化学药物可导致耐药蜱的产生以及环境的污染问题,相比之下抗蜱疫苗更加环保并且有效减少蜱的数量,是替代化学杀虫剂控制蜱感染和病原传播的有效方法[12],研究显示H.flava热休克蛋白70-b2具有良好的免疫原性,可诱发体液免疫[13]。
铁蛋白1(ferritin 1,Fer1)是调节蜱体内铁的储存和释放来维持铁代谢平衡的关键蛋白[14]。AV422蛋白是一种保守蜱唾液腺蛋白,其能在蜱的吸血过程中起到抑制血液凝固的作用[15]。本研究通过从褐黄血蜱转录组数据中(GSE67247 和GSE69721)筛选出唾液腺蛋白(HfAV422)和铁蛋白1(HfFer1)两个基因进行原核表达,通过动物试验来评价重组蛋白的抗蜱效果,为褐黄血蜱的防控提供参考。
1 材料与方法
1.1 虫株和实验动物
褐黄血蜱采自于四川省放牧的山羊体表,经分子鉴定为褐黄血蜱[6],并在四川农业大学动物寄生虫病研究中心通过新西兰兔进行传代繁殖与保种。饱血雌蜱置于装有湿润棉花的培养皿中,在温度为26 ℃±1 ℃,相对湿度为80%±10%的条件下孵育、产卵。卵在21 d左右孵化成幼蜱,经14~21 d静止期后进行试验。
22只新西兰兔(平均体重3 kg±0.2 kg),购自成都达硕实验动物有限公司。在试验期间,每只兔独笼饲养,给予充足的水和食物。
1.2 主要试剂与仪器
RNA提取试剂盒(TaKaRa MiniBEST Universal RNA Extraction Kit)购自北京天根生化科技有限公司;反转录试剂盒(Prime ScriptTMRT reagent Kitwith gDNA Eraser)购自成都福际生物技术有限公司;T4连接酶、QuickCutTM限制性内切酶SacⅠ/XhoⅠ/BamHⅠ/HindⅢ购自大连宝生物有限公司;HRP-标记山羊抗兔IgG购自武汉博士德生物工程有限公司;Rabbit IL-2、IL-4和IFN-γ ELISA Kit 试剂盒购自江苏酶免实业有限公司。
PCR仪、凝胶成像分析系统、蛋白质电泳仪购自美国Bio-Rad公司;中高压蛋白纯化系统购自美国Cytiva公司;控温摇床(HZQ-100)购自哈尔滨东联电子技术开发有限公司。
1.3 褐黄血蜱总RNA的提取和cDNA合成
将褐黄血蜱饥饿幼蜱按照提取总RNA提取试剂盒说明书进行RNA的提取。以抽提的褐黄血蜱总RNA为模板,使用反转录试剂盒进行cDNA合成,将所得产物保存于-80 ℃。
1.4 生物信息学分析及目的基因扩增
从褐黄血蜱转录组数据获取褐黄血蜱HfFer1和HfAV422的CDs序列,利用生物信息学分析软件分别对这两个基因进行生物信息学分析。切除信号肽和跨膜区后然后利用Primer 6.0软件进行引物设计(表1),斜体为保护性碱基。

表1 HfFer1和HfAV422的序列设计
1.5 HfFer1和HfAV422基因的原核表达、蛋白纯化及免疫印迹分析
将阳性菌液进行扩大培养和提取质粒,质粒用相对应的快切酶进行酶切然后进行T4连接测序。将测序正确的质粒转入BL21感受态细胞中,挑取单个菌落接种于1 L含氨苄抗性的培养液中,37 ℃摇床培养至菌液OD590 nm为0.6后加入诱导剂IPTG(1 mmol·L-1),37 ℃诱导12 h(160 r·min-1),随后进行可溶性分析,取收集的上清和沉淀制样进行SDS-PAGE电泳,分析两种蛋白可溶性表达情况。用镍离子亲和层析方法对重组蛋白进行纯化。将纯化后的蛋白进行免疫印迹分析,以1∶100稀释的褐黄血蜱阳性兔血清作为一抗,以1∶2 000稀释的HRP标记的山羊抗兔IgG作为二抗。
1.6 重组蛋白免疫保护性试验
1.6.1 试验分组及免疫程序 将新西兰兔随机分组,皂素对照组6只,蛋白组每组各8只。颈部皮下注射,相同剂量免疫两次,间隔14 d(表2)。

表2 实验动物分组和免疫程序
1.6.2 褐黄血蜱幼蜱的感染 在第二次免疫后一周,将褐黄血蜱幼蜱感染于兔耳部。每只兔感染100只幼蜱,为了方便收集饱血幼蜱,每只耳朵固定耳袋,并套上伊丽莎白圈。攻虫24 h后观察幼蜱感染情况并且计数,之后每天观察并收集饱血幼蜱,并将饱血蜱放入湿润的培养皿中并在温度为26 ℃±1 ℃的条件下进行蜕皮。对饱血幼蜱进行计数、称重以及观察其蜕皮情况。
幼蜱的免疫效果是基于Aguirre等[16]描述的方法进行计算:抗幼蜱免疫效果=[1-Ra×Rb]%,Ra代表免疫组平均饱血蜱数量与对照组平均饱血蜱数量的比值,Rb代表免疫组蜕皮率与皂素对照组蜕皮率的比值。
1.6.3 抗体水平检测 在首免前、免疫后每周固定时间以及攻虫后一周采集兔血并分离血清,使用间接ELISA方法检测IgG抗体水平(蛋白稀释比1∶100,二抗稀释比 1∶3 000)。
1.6.4 细胞因子检测 按照试剂盒说明书对二免后兔血清进行细胞因子(IL-2、IL-4和IFN-γ)测定。
1.7 数据处理
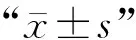
2 结 果
2.1 PCR扩增
以褐黄血蜱cDNA为模板扩增出HfFer1(GenBank No.:OP820815)、HfAV422(GenBank No.:OP820814),扩增片段大小分别为525和636 bp(图1)。
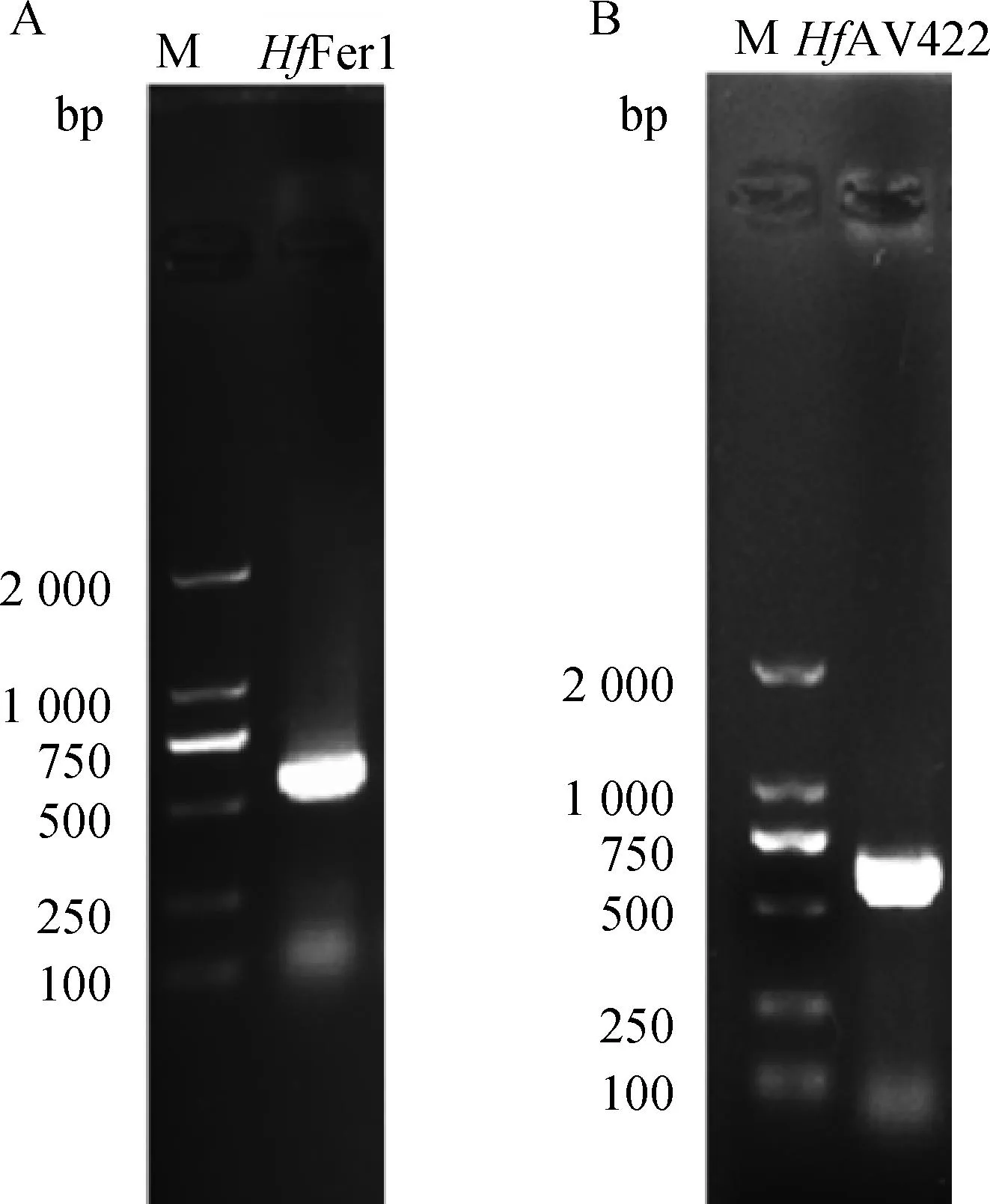
A. 基因HfFer1扩增结果;B. 基因HfAV422扩增结果;M. DNA相对分子质量标准A. HfFer1 amplication; B. HfAV422 amplication; M. DNA marker
2.2 HfFer1和HfAV422的生物信息学分析
克隆得到的HfFer1和HfAV422序列翻译成氨基酸序列与褐黄血蜱湖南株的氨基酸序列比对存在一定差异,相似度分别为99.43%和96.68%(图2)。其中HfAV422与美洲花蜱、变异花蜱氨基酸序列相似度较高,均在90%以上;与篦子硬蜱氨基酸序列的相似度较低,为85.99%(图2A)。HfFer1与长角血蜱、美洲花蜱氨基酸序列的相似度较高,分别为96.55%和94.15%;而与篦子硬蜱氨基酸序列的相似度较低,为86.47%(图2B)。

H. flava. 褐黄血蜱;A. americanum. 美洲花蜱;A. variegatum. 变异花蜱;I. ricinus. 篦子硬蜱;H. longicornis.长角血蜱;红色长条块. α螺旋;绿色箭头. β折叠;红色虚线方框. B抗原表位H. flava. Haemaphysalis flava; A. americanum. Amblyomma americanum; A. variegatum. Amblyomma variegatum; I. ricinus. Ixodes ricinus; H. longicornis. Haemaphysalis longicornis; Red rectangle. Alpha helix; Green arrows. Beta fold; Red dashed box. B antigen epitopes
2.3 原核表达、蛋白纯化及免疫印迹
测序成功后的菌液扩培提取质粒,然后进行双酶切得到的pMD19-T载体骨架和525 bp的Fer1基因片段、636 bp的AV422基因的目的片段与预期大小相符(图3)。

A. HfFer1质粒双酶切;B. HfAV422质粒双酶切;M. DNA相对分子质量标准Marker ⅢA. Enzyme digestion of HfFer1 plasmid; B. Enzyme digestion of HfAV422 plasmid; M. DNA Marker Ⅲ
rHfFer1重组蛋白在25 ℃、1 mmol·L-1浓度的IPTG的条件下诱导12 h时能够大量表达,并且表现为可溶性表达;rHfAV422重组蛋白在37 ℃、1 mmol·L-1浓度的IPTG的条件下诱导24 h后能够大量表达,其重组蛋白主要表达在包涵体中(图4)。重组蛋白通过镍离子亲和层析柱纯化后,经凝胶电泳分离,结果显示纯化后的蛋白均为单一条带(图5)。免疫印迹分析rHfFer1、rHfAV422均能识别阳性血清,具有良好的免疫反应性(图6)。
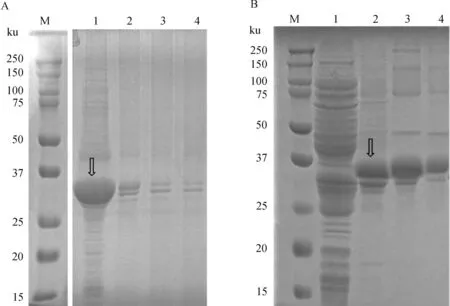
M. 蛋白质相对分子质量标准(ku);1. 重组菌诱导表达后菌体裂解物上清;2~4. 菌体裂解物先后溶解于2、4、8 mol·L-1尿素M. Protein molecular weight markers (in ku); 1. Soluble protein; 2-4. Inclusion body in 2, 4, 8 mol·L-1 urea
2.4 rHfAV422和rHfFer1的免疫保护效果
2.4.1 血清中特异性IgG抗体水平的变化 两个免疫组分别免疫接种rHfFer1和rHfAV422后,每周在固定时间进行采血和收集血清。动物试验结束后对血清中的特异性IgG抗体水平进行检测。皂素对照组在免疫前后血清中平均IgG抗体水平一直维持在较低水平。而免疫组在免疫后血清中的平均IgG抗体水平明显升高,其平均水平显著高于皂素对照组的特异性IgG抗体水平(P<0.05),并且在攻虫后一周的抗体同样保持较高水平(图7)。
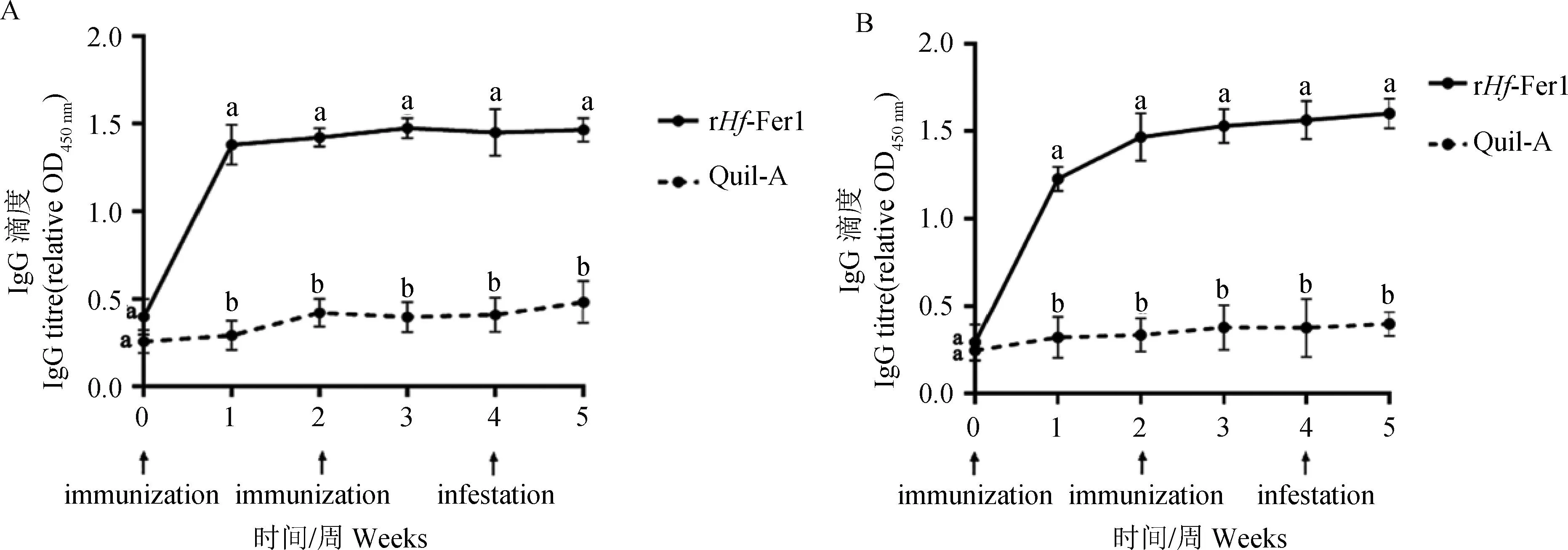
immunization分别代表第一次和第二次免疫蛋白时间;Infestation.感染幼蜱的时间。不同字母标注的数据间存在显著性差异(P<0.05),相同字母标注的数据间不存在显著性差异(P>0.05)Immunization: Time of the first and second immunization with vaccine; Infestation. Time of challenge with larvae. There is a statistically significant difference between data labeled with different letters (P<0.05) and there is no statistically significant difference between data labeled with the same letters (P>0.05)
2.4.2 细胞因子水平分析 rHfFer1免疫组IL-2水平显著高于皂素对照组,而IL-4和IFN-γ无明显变化;rHfAV422免疫组IL-2和IL-4水平与皂素对照组存在显著性差异,IFN-γ无明显差异(图8)。

不同字母标注的数据间存在显著差异(P<0.05),相同字母标注的数据间不存在显著差异(P>0.05)There is a statistically significant difference between data labeled with different letters (P<0.05) and there is no statistically significant difference between data labeled with the same letters (P>0.05)
2.4.3 抗蜱效果分析 从试验结果来看rHfFer1免疫组的幼蜱饱血率和蜕皮率(41.13%和54.98%)均明显低于皂素对照组(84.25%和80.25%)(P<0.05);rHfAV422免疫组的幼蜱饱血率和蜕皮率(64.78%和63.29%)与皂素对照组的饱血率和蜕皮率同样存在显著性差异(P<0.05)。而两个免疫组与皂素对照组的幼蜱饱血体重均无明显差异(表3)。rHfAV422和rHfFer1蛋白的抗蜱效力分别50.00%和68.06%。

表3 免疫 rHfAV422和rHfFer1蛋白对幼蜱摄食和蜕皮参数的影响
3 讨 论
使用抗蜱疫苗诱导宿主产生针对蜱虫和蜱媒病原体感染的免疫保护是一种经济有效的替代方法,它可减少化学杀虫剂的使用和耐药蜱的产生[17-19]。抗蜱疫苗的设计目的不是预防蜱的感染,而是通过蜱摄取宿主体内的抗体与蜱体内靶蛋白相互作用后破坏靶蛋白的功能,从而影响蜱的摄食、蜕皮和繁殖来控制蜱的数量[20-21]。早在1995年,Willadsen等[22]报道从微小扇头蜱(Rhipicephalusmicroplus)中肠提取出的保护性抗原Bm86免疫牛后能够明显降低牛体表蜱虫的感染数量,此后基于此抗原开发出GARDTM和GavacTM两款商品化蜱疫苗,在大规模田间试验中,基于Bm86抗原的疫苗能够有效降低牛感染微小扇头蜱(包括耐药蜱株)的概率,而且还能够控制血液寄生虫的感染[23]。不过Bm86疫苗仅对微小扇头蜱和扇头蜱属的蜱具有良好的抗蜱效果,对其他蜱种的防治效果有限。此后,有多种抗原(如丝氨酸蛋白酶抑制剂、谷胱甘肽S-转移酶、金属蛋白酶、4D8等)被证明可用作抗蜱疫苗候选抗原[24]。然而筛选高效且具有交叉保护作用的抗原仍为今后抗蜱疫苗研发的主要方向。蜱作为一种专性吸血的体表寄生虫进化出一套特殊的生存机制,包括能够长时间附着于宿主体表吸血,防止体内血液凝固以及长期储存中肠内血餐等[25]。血餐是蜱的唯一营养来源,蜱的每一次蜕皮以及雌蜱完成产卵都需要吸食足够的血液方可完成,而参与蜱吸血相关生理过程的基因成为抗蜱疫苗候选抗原的重要来源。
有研究报道,长角血蜱(Haemaphysalislongicornis)甘肃株P27/30基因的核苷酸序列和氨基酸序列与长角血蜱日本Okayama株、长角血蜱四川孤雌生殖株均存在差异,推测蜱株的地域性分布差异可能导致虫株之间的氨基酸序列存在一定的差异[26]。在本试验中扩增的褐黄血蜱HfFer1和HfAV422基因编码的氨基酸序列与褐黄血蜱湖南株的相似度分别为99.43%和96.68%,这表明褐黄血蜱四川株与湖南株也存在一定的差异。


铁蛋白(ferritin)广泛存在于动物、植物和微生物中,负责铁存储和运输铁元素,在维持生物体的中维持铁的动态平衡起着关键作用[27]。该蛋白主要由24个亚基组成,折叠成4个螺旋束后形成1个近乎球形外壳,并且有1个可以容纳多达4 000个铁原子的巨大空腔[28]。Fer1是一种关键细胞保护蛋白,其主要负责维持铁代谢稳态和保护机体免受氧化损伤[27]。在蜱中Fer1通过将进入细胞质的Fe2+转化成Fe3+后储存在细胞内以减少Fe2+的氧化毒性,当机体需铁时再将Fer1中储存的铁释放出来[29]。在有关沉默蜱体内Fer1基因的研究中,发现Fer1基因的沉默能够降低蜱的摄食、产卵及卵孵化能力,甚至导致饱血蜱死亡[30-32]。同样,有学者对兔免疫长角血蜱(H.longicornis)重组蛋白rHlFer1后,观察到饱血雌蜱产卵重量明显降低(降低16%),并且在卵孵化期间部分幼虫出现死亡[33]。这说明Fer1在蜱的吸血过程和繁殖中起关键作用。在本试验中,接种rHfFer1蛋白后明显降低了幼蜱的饱血率和蜕皮率,这可能是幼蜱从兔体内摄取的Fer1抗体与虫体体内的Fer1发生反应,导致幼蜱因氧化应激而影响其饱血和蜕皮能力甚至引起死亡。
AV422最初在美洲花蜱体内发现,其转录水平在吸血后7 d的唾液腺中明显上升,这表明其可能是一种唾液蛋白并且在吸血过程中发挥重要作用[15]。体外凝血试验显示,rHfAV422能够延长凝血酶原时间(prothrombin time,PT)、活化部分凝血酶时间(activated partial thromboplastin time,APPT)和凝血酶(thrombin time,TT)时间,从而起到抗凝血作用[34]。美洲花蜱(Amblyommaamericanus)重组蛋白rAamAV422能够延迟血浆凝固并抑制血小板聚集,并且当在沉默AamAV422 mRNA之后,该蜱饱血后的重量显著降低(约44%)[35],这说明AV422在蜱吸血过程中可能会干预多个凝血步骤而起到抑制血液凝固的作用。在蜱体内血餐被消化之前会以液体的形式在消化道储存,而血液的凝固可能会阻止血餐的摄取和消化,因此抗凝作用对蜱的存活至关重要[34]。有趣的是,AV422蛋白是蜱虫体内独有的一种蛋白,在其它节肢动物和脊柱动物体内没有同源物,这可能意味着当该蛋白作为疫苗靶向抗原时,对脊椎动物宿主可能不会产生副作用[35]。AV422蛋白通常会在蜱吸血过程中从唾液腺分泌出进入宿主体内,从而引起宿主体液免疫反应。有研究发现篦子硬蜱(Ixodesricinus)重组蛋白rIrAV422能够与感染过网纹革蜱(Dermacentorreticulatus)的大鼠血清和有过蜱虫感染史的猎犬和人血清发生交叉反应,表明AV422具备交叉免疫反应性特征[35-37]。在本研究中,作者发现rHfAV422蛋白免疫实验兔后表现出良好的抗蜱效果,与皂素对照组饱血率和蜕皮率相比较,免疫组幼蜱的饱血率和蜕皮率更低,说明AV422作为一种蜱保护性抗原,具有开发为控制不同蜱种的抗蜱疫苗的潜力。
IL-2、IL-4和IFN-γ在宿主免疫反应中发挥重要作用,其中IL-2和IFN-γ主要是由Th1细胞分泌,介导细胞免疫应答;IL-4是由Th2细胞分泌,它可促进B细胞分化和抗体的产生[38]。研究显示,蜱唾液腺能促进IL-4上调使细胞因子向Th2型极化[39-40]。蜱在自然感染宿主过程中主要是引起Th2型免疫反应[41-43],其可能是因为蜱在叮咬宿主时从蜱唾液腺中分泌蛋白进入宿主体内引起IL-4、IL-10等相关因子的上调,诱导宿主Th2型免疫反应。在璃眼蜱(Hyalomma)rHaa86对杂交牛的免疫应答中发现rHaa86体外刺激免疫后不同时间段的外周血液淋巴细胞,IFN-γ表达水平均无明显变化,这可能是因为该重组蛋白引起了宿主Th2型免疫反应[44]。不过有研究显示向兔注射重组IL-2后表现出更强的抗蜱能力,能够有效降低篦子硬蜱(Ixodesricinus)成蜱的饱血能力以及产卵重量,因此细胞免疫在蜱的免疫应答中同样扮演重要角色[45]。本次试验结果显示,两个免疫组的产生的IFN-γ水平与皂素对照组无明显差异,但IL-2浓度水平与皂素对照组存在显著差异,rHfAV422免疫组的IL-4浓度水平与皂素对照组同样存在显著性差异,这表明rHfFer1能够引起Th1型免疫反应,而rHfAV422能够同时引起Th1型和Th2型免疫反应。
4 结 论
本研究通过原核表达获得了rHfFer1和rHfAV422,rHfFer1和rHfAV422两个蛋白分别按500 μg剂量对兔进行两次免疫后产生的抗蜱效率分别为68.06%和50.00%,rHfFer1可引起Th1型免疫反应,而rHfAV422能够同时引起Th1型和Th2型免疫反应。本研究结果表明这两个抗原具有成为抗蜱疫苗候选抗原的潜力。
