猫犬主要过敏原蛋白的最新研究进展
2024-03-01李春晓高博泉王振龙王金全王秀敏
李春晓,安 尉,高博泉,王振龙,韩 冰,陶 慧,王金全,王秀敏
(中国农业科学院饲料研究所,农业农村部饲料生物技术重点实验室,北京 100081)
过敏是日常生活中很常见的问题之一,严格来说,过敏是机体产生的一种过激的免疫反应,机体将外来的一些物质视为异物从而引发机体免疫反应过度,继而导致人或动物体发生一系列过敏反应。宠物过敏原诱发的症状严重程度差异很大,从与鼻炎和结膜炎相关的不适到严重的哮喘;哮喘可能发展成危及生命的疾病,猫犬过敏原被认为是哮喘和过敏性鼻炎发展的主要危险因素[1]。对猫过敏患者而言,在自然条件下接触一定的猫过敏原会导致严重的外周气道阻塞,它可以发展成危及生命的状况[2]。根据一项针对吸入性过敏原的大规模皮肤点刺试验(SPT)研究(全球哮喘和过敏欧洲网络)显示,因疑似吸入性过敏原而就诊的欧洲成年人中约26%对猫过敏,27%对犬过敏[3-4]。我国对儿童支气管哮喘、变应性鼻炎、湿疹、过敏性咳嗽、急慢性荨麻疹、过敏性紫癜等常见过敏性疾病的过敏原进行分析的流行病学调查显示,学龄前儿童和学龄期儿童吸入性过敏原过敏者分别占66%和73.1%[5]。家养猫和家养犬中最主要的过敏原分别是Fel d 1和Can f 1[6]。猫犬过敏原提取物含有多种过敏原,目前已有八种猫的过敏原(Fel d 1~Fel d 8)以及八种犬的过敏原(Can f 1~Can f 8)在WHO/IUIS中注册。
猫犬过敏原在世界范围内作为常见的室内吸入性过敏原,参与免疫球蛋白E(IgE)介导的过敏反应,IgE介导的过敏反应属于Ⅰ型免疫,与其他类型免疫反应相比时间进程更快,抗原识别后继发反应的幅度更大,这些后果可能高度危及过敏患者的生命[6]。目前,还缺乏完全有效和安全的方法治疗宠物过敏。
1 猫犬主要过敏原蛋白的结构与功能分析
1.1 猫源过敏原蛋白
在猫的皮屑、毛发、唾液、舌下腺等提取物中发现有多种过敏原,包括猫过敏原蛋白Fel d 1~Fel d 8。过敏原Fel d 1是分泌珠蛋白家族的成员,是最主要的猫过敏原[7],主要在舌下和皮脂腺中产生,肛腺和泪腺也会产生[8]。Fel d 1主要储存在唾液和毛发中,当猫咪梳理自己毛发时,Fel d 1也会转移到猫毛上。另外,含有Fel d 1过敏原的猫皮屑作为小的空气传播颗粒传播到环境中[9]。Fel d 1是由两个异源二聚体形成的四聚糖蛋白(图1),二聚体之间以非共价键的方式链接,每个异源二聚体由一个70残基肽和一个85、90或92残基肽组成。这些链在每个异源二聚体中通过二硫键共价连接[10]。Fel d 1的结构与兔子宫珠蛋白的结构具有相似性,其链Ⅰ与子宫珠蛋白共享氨基酸同源性,而链Ⅱ是具有N-端连接寡糖的糖蛋白[11]。在Fel d 1内部有一个不对称空腔,可以结合内源性配体[7];在体内,Fel d 1的过敏原性由黏膜抗原呈递细胞(如树突状细胞)上的甘露糖受体识别决定[12]。

图1 猫主要过敏原Fel d 1蛋白结构[10]Fig.1 Structure of major cat allergen Fel d 1[10]
Fel d 2是一种血清白蛋白,是皮屑中的重要蛋白质,它是一种引起轻微过敏症状的猫过敏原,可在单重和多重免疫分析中作为天然纯化和重组分子获得。Fel d 3是一种半胱氨酸蛋白酶抑制剂,也是一种次要过敏原,属于半胱氨酸蛋白酶抑制剂(CPI)的半胱氨酸蛋白酶抑制剂超家族,是一种小的酸性蛋白,没有半胱氨酸残基或二硫键。Fel d 4和Fel d 7为脂质激素,Fel d 8是一种独特的泡沫蛋白样蛋白,属于脂多糖结合蛋白/杀菌通透性增加家族[13]。
在以上过敏原蛋白中,最重要的过敏原蛋白为Fel d 1,能使93.9%的猫科动物敏感患者敏感[14],过敏患者血清中针对猫过敏原的IgE抗体多数指向Fel d 1,占总过敏原活性的60%~90%[12]。
猫过敏原及其特性详见表1[15-16]。
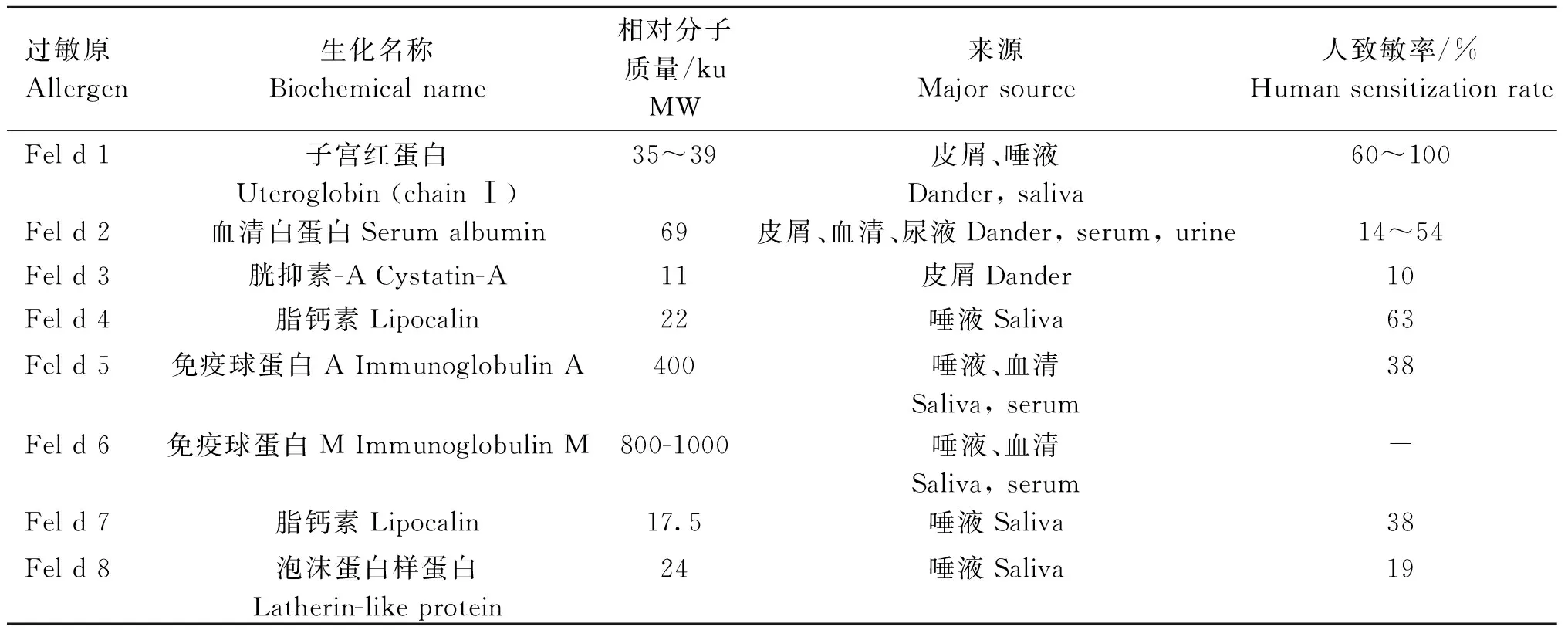
表1 猫过敏原及其特性[15-16]
1.2 犬源过敏原蛋白
犬过敏原Can f 1是一种在犬的唾液腺中产生的脂质运载蛋白,与人类泪液脂质运载蛋白同源[17-19],在多达50%~75%对犬皮屑敏感的受试者中结合IgE抗体[6],是犬主要的过敏原之一。目前,Can f 1仍然是流行病学研究中的主要过敏原[20],其蛋白结构见图2[21]。

该蛋白结构由Min等[21]解析上传至在线存储库http:∥www.wwpdb.org/,8epu 代表 Can f 1,该图像由网站 (https:∥swissmodel.expasy.org/templates/8epu.1) 生成The protein structure was analyzed by Min et al[21]and uploaded to the online repository http:∥www.wwpdb.org/, and 8epu stands for Can f 1. The image was generated by the website (https:∥swissmodel.expasy.org/templates/8epu.1)
Can f 2、Can f 4、Can f 6组分与Can f 1同属于载脂蛋白家族[5]。Can f 3来源于犬血清白蛋白,与猫的过敏原Fel d 2属于同一类蛋白家族,而同一蛋白家族的成员间可能发生发叉反应[22]。Can f 4是舌上皮组织表达的一种158个氨基酸的脂质运载蛋白,该过敏原主要存在于唾液和皮屑中[23]。
Can f 5是一种前列腺激肽释放酶(28 ku蛋白),在2009年也被确定为主要的犬过敏原,在70%的过敏患者中结合IgE,作为一种前列腺蛋白,它仅存在于公犬尿液中,并且与人类前列腺特异性抗原表现出交叉反应性,为犬过敏原暴露引起的精液超敏反应提供了潜在的解释[24]。Can f 7又称 NPC2蛋白,也称为CE1蛋白,是MD-16相关脂质识别(ML)家族中的2 ku过敏原[25-26]。此外,也在犬的皮屑中观察到了Can f 8,并将其归为胱抑素家族的一员[27]。
犬过敏原及其特性见表2[4,28-29]。
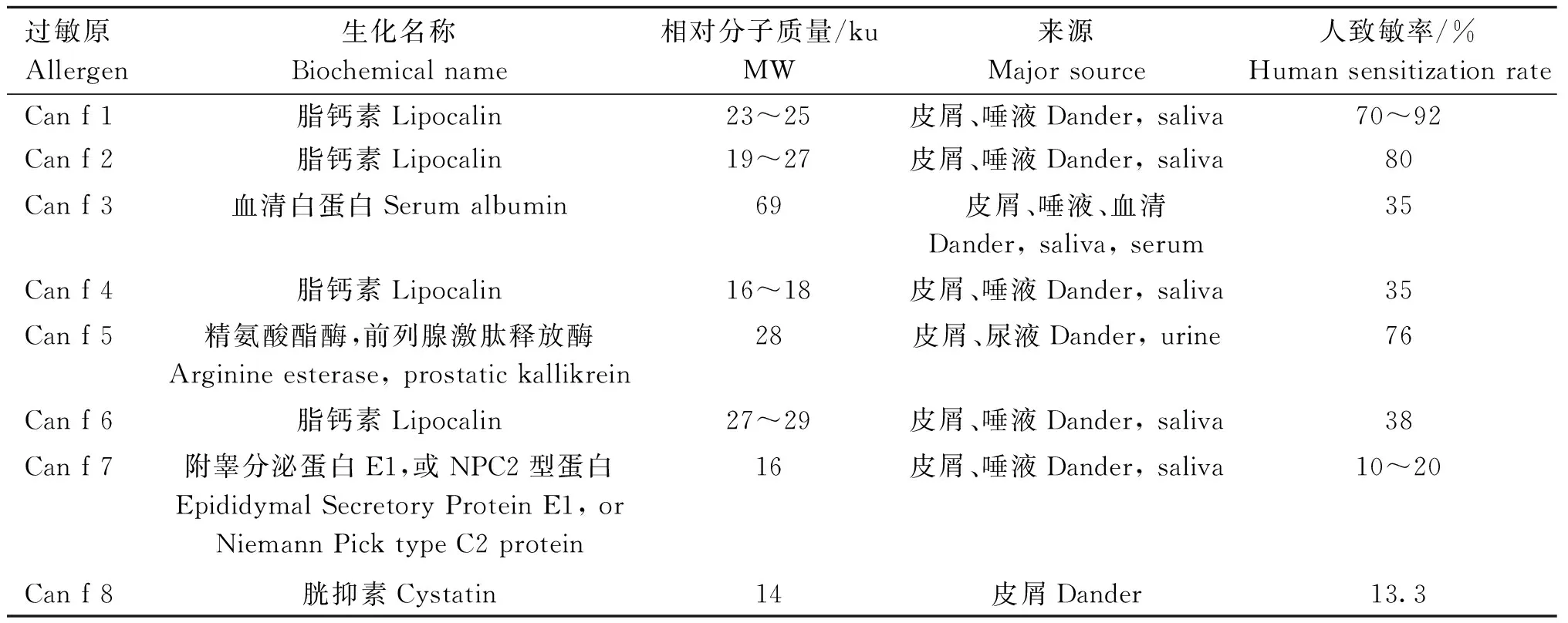
表2 犬过敏原及其特性[4,28-29]
2 猫犬过敏原蛋白的重组表达
2.1 重组表达系统
重组蛋白药物生产的表达系统主要包括真核表达系统(如哺乳动物细胞、昆虫细胞、酵母细胞等)和原核表达系统(如大肠杆菌、乳酸乳球菌等)[30]。目前批准上市的重组蛋白药物大多是在这两个表达系统中生产[31]。用于表达重组蛋白的哺乳动物细胞主要包括中国仓鼠卵巢(CHO)细胞、小鼠骨髓瘤细胞Sp2/0和NS0、人胚胎肾细胞HEK293、仓鼠肾细胞BHK-21等,它们具有类似于人源细胞的翻译后修饰、外源基因的稳定整合、可以大规模悬浮培养等优点,其中大约有70%的重组蛋白药物是由CHO细胞生产的[32]。在原核表达系统中,大肠杆菌遗传背景清楚,相对于CHO细胞,其表达量较高,能在培养基上快速繁殖、成本低, 是最常用的原核表达系统[33],但由于缺乏翻译后修饰(如糖基化),主要用于表达分子量较小、不需要翻译后修饰的重组药物[34]。
2.2 重组过敏原蛋白
在目前的研究报道中,猫过敏原Fel d 1重组蛋白一般采用原核表达系统进行表达。裴业春[35]将Fel d 1链Ⅰ、Ⅱ拼接后插入到乙肝病毒核心抗原(HBcAg)中,构建了pET28a-HBcAg-rFel d 1表达载体,将其转化入大肠杆菌BL21(DE3)中;通过诱导表达后获得了HBcAg-rFel d 1融合蛋白,大小约为37 ku。Schmitz等[36]将噬菌体Qβ的外壳蛋白的病毒样颗粒(Qβ-VLP)作为Fel d 1表达载体,并在大肠杆菌中进行了高效表达,获得了Qβ-Fel d 1融合蛋白。Curin等[37]设计了一系列Fel d 1的衍生肽(包括Fel d 1 M1、M、MA、MB、MC、ME和MF),该系列肽均无典型的α-螺旋结构,不具有IgE反应性;这些低过敏性衍生肽均含有诱导过敏原特异性阻断IgG抗体和T细胞表位的序列元件,插入表达载体pET-28b后,在大肠杆菌BL21(DE3)中进行表达,获得了重组的Fel d 1低过敏性衍生物,其表达量为2~6 mg·L-1、纯度高达90%以上,可用于过敏疫苗接种和耐受诱导。周婕[38]在构建好的Fel d 1-IL-10(FIL)重组嵌合蛋白(含Fel d 1、IL-10)的基础上,通过基因工程定点突变技术对重组猫过敏原Fel d 1序列中抗原值高的位点进行改造(23V→P,34V→P),插入载体,转化至大肠杆菌中进行诱导表达,获得猫过敏原突变体蛋白,但未验证其抗原性。
许卓谦和刘志刚[39]利用pET-24(+)构建表达载体,获得了带有his标签的Fel d 1 Ⅰ、Ⅱ链,但该重组Fel d 1蛋白发生了突变(如42 Met→Lys、55 Ile→Leu、74 Met→Thr)。此外,Grönlund等[40]发现大肠杆菌表达后的Fel d 1链Ⅰ、Ⅱ混合后与IgE抗体结合能力比天然Fel d 1下降了25倍,可能与链Ⅰ、Ⅱ之间的连接及Fel d 1的三对二硫键(Cys3~Cys73、Cys44~Cys48、Cys7~Cys70)的正确配对有关。由此可见,重组过敏原Fel d 1蛋白有时不能正确折叠,或表达量不高、位点突变及二硫键错配等问题可能会影响其免疫原性。
3 降低猫犬过敏原蛋白的治疗方法
3.1 过敏原特异性免疫疗法
过敏原特异性免疫疗法,又称脱敏治疗,是目前唯一一种治疗过敏性疾病的有效疗法,其通过长期注射过敏原诱导患者耐受该过敏原从而不再产生过敏反应,即使停药也有长期的疗效[41]。国际上从20世纪70年代开始已经开展对猫犬过敏原特异性免疫治疗的研究。过敏原特异性免疫疗法有多种给药途径,例如皮下、舌下或淋巴注射,并在小鼠和人类受试者中测试了该治疗方法的安全性和有效性,已经观察到一些非常有希望的结果[42]。多项研究表明,这种方法对过敏性患者具有安全性和多重益处,能够缓解呼吸道的主要症状,可以减少炎性反应[43-44]。Alvarez-Cuesta[45]等使用标准化猫过敏原提取物对猫毛皮屑过敏患者进行治疗,发现13.2 μg的Fel d 1抗原剂量具有一定的疗效,且呈剂量依赖性,高剂量Fel d 1提取物(15 μg)达到有效的治疗效果。据报道,患者获益与诱导针对猫主要过敏原Fel d 1的IgG抗体以及减少皮肤和呼吸道症状有关。另一项研究显示,与安慰剂组相比,接受猫科动物过敏原Fel d 1提取物皮下免疫疗法的受试者中IgG4水平升高、对过敏原的皮肤反应性降低、支气管反应性降低以及症状评分改善;临床疗效呈剂量依赖性,3.0和15.0 μg组在安慰剂组中均显示出显著变化,但彼此之间无显著差异,可能需要高剂量的过敏原以实现更有效的治疗[46]。然而,使用天然过敏原提取物或者未经改变的重组蛋白进行过敏原特异性免疫疗法更易产生副作用。Mellerup等[47]研究表明,41%接受猫过敏原提取物特异性免疫治疗的患者出现了严重的副作用。另外,用犬过敏原Can f 1提取物对狗毛皮屑过敏患者进行特异性免疫治疗的效果不佳,有待进一步探讨。
为了降低使用过敏原进行免疫治疗对患者的危害,通常需要降低过敏原蛋白的过敏性。聚合是降低过敏原蛋白过敏性最常见的方法[48]。过敏原聚合会导致暴露的IgE结合位点数量减少,使其对过敏性患者外周血中嗜碱性粒细胞的活化能力低于正常过敏原,从而降低机体的过敏症状[49]。Calzada等[27]将Can f 1和Can f 5的聚合产物作为新型犬皮屑过敏物质进行研究,发现该聚合物具有低结合IgE和激活犬过敏患者嗜碱性粒细胞的能力,这种过敏性疫苗可以提供比天然提取物更安全的特性。
3.2 疫苗
另外,基于短肽的过敏原免疫疗法是常规免疫疗法的替代方法,由于短肽缺乏使肥大细胞和嗜碱性粒细胞表面上的IgE交联所需的三维结构,可以降低IgE介导的严重反应风险。因此,基于短肽的重组过敏原有望为过敏原免疫治疗提供新的思路[50]。通过重组表达Fel d 1的T细胞抗原表位、开发一种Fel d 1低致敏原,使其能对免疫接种、诱导阻断IgG抗体以及诱导耐受性都有用;这种过敏原特异性免疫疗法一方面可以降低严重的副作用,另一方面能够减少肥大细胞或嗜碱性粒细胞上IgE的结合,从而减少过敏反应。另外,这种短肽疫苗可以靶向过敏原特异性T细胞,比传统疫苗更安全、保护率更高[51]。此外,Kaiser等[7]解析了重组Fel d 1蛋白的结构(图3),发现其由两个链组成,N端和6个C端His残基不受电子密度的限制;折叠后由8个螺旋组成,H1-H4和H5-H8对应于天然Fel d 1的链Ⅰ和Ⅱ;对重组Fel d 1抗原表位进行预测,发现IgE结合的3个重要的抗原表位是25~38、46~59与15~28位(图4),第一个抗原表位与人类血清的识别率达到46%。
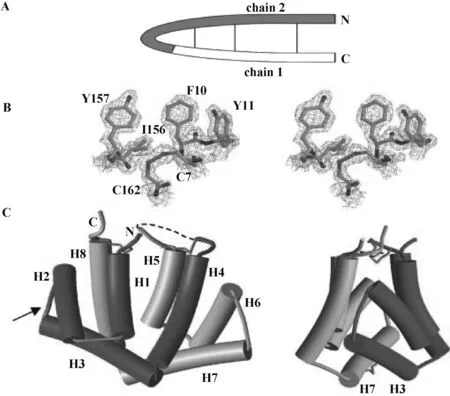
A. Fel d 1链Ⅰ和Ⅱ; B. 电子密度云; C. Fel d 1单体A. Fel d 1 chains Ⅰ and Ⅱ; B. Electron density clouds; C. Fel d 1 monomer

3个IgE表位的相对定位显示在Fel d1的分子表面上。a.方向与图3C(左)相同;b.与a垂直的视图;c.与b垂直的视图The relative localization of three IgE epitopes is indicated on the molecular surface of Fel d 1. a.The same orientation as in Fig. 3C, left; b.The perpendicular view to a; c.The perpendicular view to b
通过对猫自身的主要过敏原Fel d 1进行免疫来治疗人类由Fel d 1引起的过敏[52]。裴业春[53]按照猫过敏原Fel d 1链Ⅰ、链Ⅱ连接后进行表达,制备了重组Fel d 1蛋白疫苗,免疫虎皮猫后,发现其毛发、下颚下腺中的Fel d 1含量下降15%~60%。同时,使用表达Fel d 1的真核表达质粒proVAX-rFel d 1作为DNA疫苗,与蛋白疫苗按照1∶1比例混合后作为DNA/蛋白共免疫疫苗,并在BALB/c小鼠上进行了预防和治疗试验。结果显示,注射DNA/蛋白共免疫疫苗的过敏小鼠血清中总IgE水平降低了600~800 ng·mL-1,抑制了肺部呼吸阻力的上升,降低了应激过敏反应的激烈程度[53]。
Nakatsuji等[54]基于序列比对和三级结构,预测了犬过敏原Can f 1蛋白中IgE表位的五个关键残基(His86、Glu98、Arg111、Glu138和Arg152)(图5);结果表明,残基His86和Arg152为IgE构象表位的候选者。Immonen等[55]发现了七个Can f 1的T细胞表位区域,由这七个表位区域组成的肽可用于犬过敏的免疫治疗,这些肽表现出广泛的T细胞反应性,并且它们能够有效地与白人群体中最常见的HLA-DRB1分子结合。Juntunen等[56]合成了过敏原Can f 1 T细胞表位的肽片段,发现这些合成肽比天然肽高10~30倍的致敏性,进一步说明重组犬过敏原短肽疫苗有望成为治疗过敏的有效途径。

图5 预测的Can f 1的 IgE 表位残基[54]Fig.5 Predicted IgE epitope residues of Can f 1[54]
3.3 抗体
有研究表明,过敏原特异性中和抗体可以阻断IgE介导的过敏反应[53]。卵黄抗体(immunoglobulin of yolk, IgY)是鸟类、爬行以及两栖动物体内产生的主要抗体,与哺乳动物的IgG相似,由蛋鸡产生的IgY经转移聚集在卵黄当中,提取后能够作为预防和治疗相应疾病的多克隆抗体。一只蛋鸡每年可以产生20 g以上的IgY[57],Akbari等[58]制备的抗LPS IgY提取物的产量为40 mg·蛋黄-1;周继萍等[59]IgY抗体制备研究表明,蛋鸡在经四次免疫后IgY抗体效价达到1:51 200;段海龙等[60]制备的抗白唇竹叶青蛇毒IgY可在常温下进行制备、保存,且具有较强的耐酸性,在pH值为2的环境下仍可保持74%的活性;经胰蛋白酶处理1 h时其活性下降不明显;表明IgY具有制备简单、产量大、特异性强、稳定性强等优点。近年来已经有许多IgY针对毒素、细菌、病毒、寄生虫等方面的治疗报道,例如蛇毒[60]、诺如病毒[61]、猪带绦虫[62]、大肠杆菌[63]、鸽毛滴虫与白色念珠菌[64]等。
Satyaraj等[65]研究发现抗Fel d 1的兔多克隆抗体、过敏原特异性鸡卵黄抗体IgY能够与猫唾液中的Fel d 1结合,并有效地降低过敏患者血浆中Fel d 1的浓度,阻断Fel d 1与IgE的结合以及IgE介导的嗜碱性粒细胞脱颗粒,从而降低过敏反应。另外,他们将Fel d 1特异性卵黄抗体IgY添加到猫粮中(添加量为8 mg·kg-1),饲喂3周后,发现卵黄抗体IgY显著降低了猫毛发中Fel d 1的含量;饲喂10周后,毛发中Fel d 1的平均含量下降了47%[66]。由此可见,重组Fel d 1蛋白卵黄抗体能够中和过敏原蛋白,降低猫过敏原蛋白的分泌量,特异性抗体为对猫犬过敏症的防治提供了一种新的方法。
4 存在的问题及解决策略
近几十年来,人们为猫犬过敏原进行免疫治疗也作出了巨大努力。目前,对猫过敏患者的治疗主要通过脱敏治疗法实现,通过反复注射猫皮屑的粗提取物或源自Fel d 1的多肽,治疗后的患者仍然存在过敏症状或者产生了副作用[67-68]。过敏原阻断抗体为中和猫犬过敏原提供了一种新的思路,包括猫犬过敏原的重组、过敏原阻断抗体的制备及其在靶动物体内阻断的实际应用效果等研究。但是,重组过敏原全长蛋白的策略往往不能正确折叠,其表达量不高、位点突变及二硫键错配等问题可能会影响其免疫原性,有待进一步从抗原表位序列上进行优化。今后,基于抗原多表位融合的策略研发高效特异性生物制剂(如特异性IgY抗体),不仅可以中和猫犬体内的过敏原蛋白,从源头上降低其含量,而且比疫苗更容易被机体吸收,有望解决宠物毛发过敏症的源头问题。
