棘脑通的抗抑郁作用及机制研究Δ
2024-02-26夏辰希董婷霞詹华强
黄 雯,高 雄,夏辰希,段 然,董婷霞,詹华强
(1.广州医科大学附属肿瘤医院药学部,广州510095; 2.香港科技大学生命科学部暨中药研发中心,香港特别行政区 999007)
抑郁症是常见的情感性精神障碍。随着现代社会工作生活压力增加,抑郁症已成为世界性的公共卫生难题。2019年中国精神障碍的流行病学调查结果表明,我国抑郁症患者约9 500万例,终生患病率达6.9%。抑郁症患者在情绪上表现为持续性的低落、悲观、冷漠,也存在睡眠障碍、认知功能受损等,常伴发食欲减退、便秘等胃肠道症状[1-4]。庞大的患病群体及较高的致残或自杀比例也表明抑郁症需要引起更多的关注和重视。
棘脑通是由香港科技大学中药研发中心研发、香港宏康生物技术有限公司生产的中药保健食品(产品编号:899959028045,批号:HKUST4)。其主要成分为沙棘提取物及花生壳提取物,功能主治为认知力下降、记忆力减退、决定能力下降、语言表达衰退和社交障碍。研究表明,沙棘总黄酮类具有改善心肌细胞功能、降血糖和提高免疫等广泛的药理活性,可以模拟和增强神经营养因子的功能,诱导神经细胞分化,可用于各种脑部疾病。槲皮素作为沙棘的主要成分,属于黄酮醇类化合物,近年来有大量文献报道其在抗抑郁方面有良好的活性,而且中药沙棘因其多成分、多通路、多靶点及不良反应小的优势成为抗抑郁药物的研究热点[5-8]。花生壳为豆科植物花生的果壳,其多作为农作物废物当柴烧或制成肥料,近年来有文献指出花生壳提取物具有抗氧化、降血脂和降血压等活性,其主要化学成分为黄酮类化合物,有研究证实了花生壳乙醇提取物及其主要成分木犀草素对抑郁症的治疗效果,以及对炎症和肠道菌群的调节作用,而肠道菌群的稳态对抑郁症的各项症状具有显著影响[9-10]。以上研究表明,沙棘提取物和花生壳提取物单独使用均可以增强神经营养因子的功能或治疗抑郁症。以两种提取物结合为主要成分的棘脑通是否具有协同抗抑郁的作用?本研究将从细胞水平及整体动物水平系统评价棘脑通的抗抑郁药效,为棘脑通的临床应用提供实验依据。在整体水平上,通过建立小鼠慢性不可预知温和应激(CUMS)抑郁模型,每日对小鼠进行随机轻度刺激,连续刺激60 d,模型建立后连续14 d给予不同剂量的棘脑通,最后进行抑郁行为学检测。
神经营养因子是一类能促进神经细胞生长、发育和分化的多肽或蛋白质,能调节神经元存活,激活酶的活性,阻止成年神经元损伤后的死亡,具有促进神经元损伤修复以及轴突再生,调节突触可塑性和神经递质等功能[4]。神经营养因子已被用于预防或治疗神经退行性疾病。目前已鉴定的神经营养因子包括神经生长因子(NGF)、脑源性神经营养因子(BDNF)、神经营养因子-3(NT-3)和神经营养因子-4(NT-4)。其中BDNF广泛存在于大脑中,是参与机体情绪和认知功能的重要调节因子,能够促进脑内神经发生和突触发育。本研究将重点围绕棘脑通对BDNF释放的调节作用,以阐明其抗抑郁作用机制。
1 材料
1.1 实验动物
健康SPF级ICR小鼠,雄性,体重15~25 g,动物申请许可证号为SCXK(沪)2022-0005,购自上海斯莱克实验动物有限责任公司。动物饲养于南京中医药大学实验动物中心SPF环境中,常规饲养,温度22~25 ℃,湿度40%~70%。
1.2 细胞株
大鼠肾上腺髓质嗜铬瘤分化细胞株(PC12细胞);人神经母细胞瘤细胞株(SH-SY5Y细胞);小鼠小胶质细胞株(BV2细胞);小鼠单核巨噬细胞白血病细胞(RAW 264.7细胞),由均美国ATCC公司提供。
1.3 药品与试剂
棘脑通由香港科技大学中药研发中心提供;Dulbecco改良的Eagle培养基(DMEM培养基,批号为2472941,美国Thermo Fisher公司),用于细胞培养的胎牛血清(FBS,批号为2391593,美国Invitrogen公司),马血清(批号为2310170,美国Invitrogen公司),全反式维A酸(U0126,美国Sigma公司);胰蛋白酶和磷酸盐缓冲溶液(PBS,批号为5325271,美国Thermo Fisher公司);Thioflavin T(ThT,批号为T3516,美国Sigma公司);姜黄素(批号为C7727,美国Sigma公司);N-乙酰半胱氨酸(NAC,CS-6942,美国Sigma公司);叔丁基过氧化氢(tBHP,批号为416665,美国Sigma公司);黄芩素(批号为A2309297,上海阿拉丁公司);Erastin(批号为HY-15763,美国Sigma公司);地塞米松(DEX,批号为D4902-1G,美国Sigma公司);脂多糖(LPS,批号为048M439V,美国Sigma公司)。
2 方法
2.1 棘脑通对CUMS小鼠模型的抗抑郁作用评价
2.1.1 CUMS模型建立和给药:动物安置7 d以适应环境,进行体重与糖水偏嗜率测试,剔除数值异常的小鼠。选取测试评分正常的小鼠50只,随机分为5组,每组10只。分别为空白组、模型组、棘脑通低浓度组(对应60 kg的人,600 mg/d)、棘脑通高浓度组(对应60 kg的人,1 600 mg/d)和氟西汀组。每日随机给予不同的2种刺激建立CUMS模型小鼠,刺激方式包括禁水24 h、禁食24 h、斜笼、电击、束缚4 h、湿笼、昼夜颠倒。持续8周,进行糖水偏嗜测试,判断抑郁表型,然后对抑郁小鼠进行灌胃给药,持续14 d。空白组小鼠与CUMS模型小鼠分开饲养,不给予任何刺激。动物的分组和给药情况见表1。
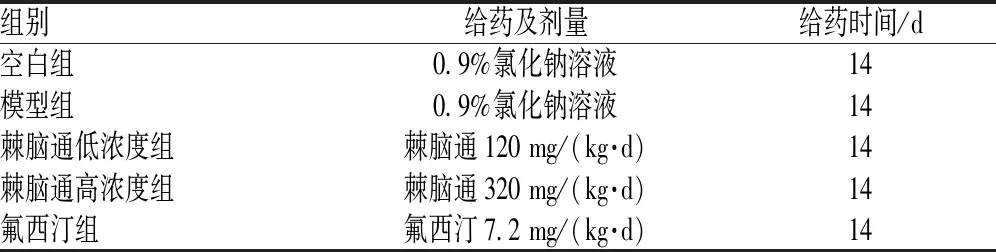
表1 棘脑通小鼠的分组及给药情况(n=10)
2.1.2 糖水偏嗜实验:通过糖水偏嗜实验考察棘脑通对CUMS模型小鼠快感缺失行为的影响。糖水偏嗜实验是通过测量小鼠的蔗糖偏好和摄入量的降低来显示小鼠对奖励敏感性的变化,从而评估小鼠的抑郁程度。实验分为糖水适应阶段和测试阶段,糖水适应阶段的小鼠单笼饲养,第1日给予小鼠200 mL 1%的蔗糖水溶液2瓶,适应24 h后将1瓶糖水换成1瓶200 mL的纯水,第3日禁食禁水,第4日正式测试阶段分别给予小鼠1%蔗糖水溶液和纯水各200 mL,记录各水瓶重量。4 h后记录小鼠糖水和纯水的消耗量,计算小鼠的糖水偏嗜率,糖水偏嗜率=糖水消耗量/(糖水消耗量+纯水消耗量)×100%。
2.1.3 悬尾实验:悬尾实验用于考察棘脑通对小鼠行为绝望的影响。在距离小鼠尾部约1 cm处粘贴胶布,悬挂于平台上,使其头部与实验台面约15 cm,悬挂6 min,小鼠以头朝下的姿势悬挂,刚开始悬挂时会挣扎和晃动,随之会放弃挣扎,记录小鼠在2~6 min内的不动持续时间。
2.1.4 强迫游泳实验:给药结束后对CUMS小鼠进行强迫游泳实验,考察棘脑通对其行为绝望的影响。将小鼠单独放置于装有干净水的圆柱玻璃缸内,保持缸内水温25 ℃,水深约20 cm,强迫小鼠游泳6 min,游泳时使小鼠后爪不能碰触水缸,小鼠的不动状态为小鼠放弃挣扎,身体被动漂浮在水面上,记录小鼠在2~6 min内的不动持续时间。
2.2 荧光素酶报告基因法检测棘脑通协同NGF促进神经分化和神经微丝的表达
为了探讨棘脑通抗抑郁的可能机制,首先考察不同浓度的棘脑通与不同浓度NGF在单独或协同作用下诱导PC12细胞分化的作用,并利用含有神经微丝编码基因启动子的荧光素酶报告基因载体pNF200-Luc考察神经微丝表达。将PC12细胞接种于24孔细胞培养板上,培养基为DMEM培养基+6%胎牛血清+6%马血清。24 h后,将含有神经微丝编码基因启动子的荧光素酶报告基因的质粒pNF200-Luc转染至PC12细胞中。4 h后,换成低血清培养基,随后加入不同浓度的棘脑通(0、10、30、100 μg/mL)辅以不同浓度的NGF处理。24 h后,经处理得到含有荧光素酶的细胞裂解液。将50 μL细胞裂解液转移至不透光的96孔酶标板上,加入荧光素酶的底物,用化学发光仪检测荧光素酶活性。
2.3 荧光素酶报告基因法检测棘脑通协同BDNF、NT-3和NT-4促进神经微丝的表达
除了NGF外,本研究还考察了棘脑通对神经营养因子BDNF、NT-3和NT-4诱导神经微丝表达的协同作用。将SH-SY5Y细胞接种于24孔细胞培养板上,培养基为DMEM培养基+15%胎牛血清。24 h后,更换培养基继续培养5 d。将含有神经微丝编码基因启动子的荧光素酶报告基因的质粒pNF200-Luc转染至SH-SY5Y细胞中。8 h后,换成无血清的DMEM培养基,随后加入100 μg/mL的棘脑通辅以不同种类不同浓度的神经营养因子处理。24 h后,经处理得到含有荧光素酶的细胞裂解液。将细胞裂解液50 μL转移至不透光的96孔酶标板上,加入荧光素酶的底物,用化学发光仪检测荧光素酶活性。
2.4 棘脑通对抗β-淀粉样蛋白(Aβ)引起神经损伤的作用考察
Aβ是淀粉样前体蛋白被β-和γ-分泌酶分解后形成的一系列蛋白肽,阿尔茨海默病的主要标志之一为出现Aβ不溶性原纤维聚集体。本研究考察了浓度梯度的棘脑通对Aβ聚集的抑制效果,以及保护PC12细胞活性、降低Aβ引起的神经毒性损伤的作用。(1)分别将浓度梯度的棘脑通或姜黄素(阳性对照)与100 μmol/L的Aβ混合,37 ℃孵育。7 d后分别与10 μmol/L的ThT混合,加入黑色不透光的96孔酶标板,使用FlexStation 3 Multi-Mode Microplate Reader(Molecular Devices)检测荧光读数。(2)将PC12细胞接种至96孔板中培养(DMEM培养基+6%胎牛血清+6%马血清),24 h后,将培养基更换为低血清培养基,并使用浓度梯度的棘脑通及姜黄素(阳性对照)对细胞进行预保护4 h,后加入100 μmol/L的Aβ处理细胞48 h。细胞活性采用四甲基偶氮唑盐比色(MTT)法进行检测。
2.5 棘脑通对抗氧化应激诱导神经损伤的作用考察
(1)将PC12细胞接种至96孔板中培养,24 h后,将培养基更换为低血清培养基,并使用浓度梯度的棘脑通或5 mmol/L的NAC(阳性对照)对细胞进行预保护4 h,后加入200 μmol/L的tBHP处理细胞24 h。细胞活性采用MTT比色法进行检测。(2)将SH-SY5Y细胞种于96孔板中培养(DMEM培养基+15%胎牛血清)。24 h后,将培养基换成低浓度培养基,并使用浓度梯度的棘脑通及10 μmol/L黄芩素(阳性对照)对细胞进行预保护4 h,后加入1 μmol/L的Erastin(铁死亡的诱导剂)处理细胞24 h。细胞活性采用MTT比色法进行检测。
2.6 棘脑通的抗炎作用考察
(1)将BV2细胞种于24孔板中培养(DMEM培养基+10%热灭活胎牛血清)。24 h后,使用浓度梯度的棘脑通或地塞米松(阳性对照)对细胞进行预保护4 h,后加入100 ng/mL的LPS处理细胞以诱导炎症反应。24 h后,移除细胞培养液,并用PBS冲洗。随后用RNAzol行总细胞RNA提取,反转录后对cDNA进行荧光定量聚合酶链反应,测定促炎因子白细胞介素(IL)1β的表达水平。(2)将RAW 264.7细胞种于24孔板中培养(DMEM培养基+10%热灭活胎牛血清)。24 h后,将含有核因子κB(NF-κB)基因启动子的荧光素酶报告基因的质粒pNF-κB-Luc转染至RAW 264.7细胞中。8 h后更换培养基,并使用浓度梯度的棘脑通及地塞米松为阳性对照对细胞进行预保护4 h,后加入100 ng/mL的LPS处理细胞。24 h后,经处理后得到含有荧光素酶的细胞裂解液。将50 μL细胞裂解液转移至不透光的96孔酶标板上,加入荧光素酶的底物,用PromegaGlomax 96孔化学发光仪(美国Glomax公司)检测荧光素酶活性。
2.7 统计学方法
采用SPSS 20.0统计软件进行统计分析,组间比较采用单因素方差分析,P<0.05为差异有统计学意义,P<0.01为差异有极显著统计学意义。
3 结果
3.1 棘脑通对CUMS小鼠糖水偏嗜率的影响
模型组小鼠糖水偏嗜率较空白组显著降低,差异有统计学意义(P<0.01);棘脑通给药组小鼠糖水偏嗜率较模型组显著升高,提示棘脑通的摄入会刺激抑郁小鼠摄入更多的糖水,作用与阳性药物氟西汀相当,见图1。
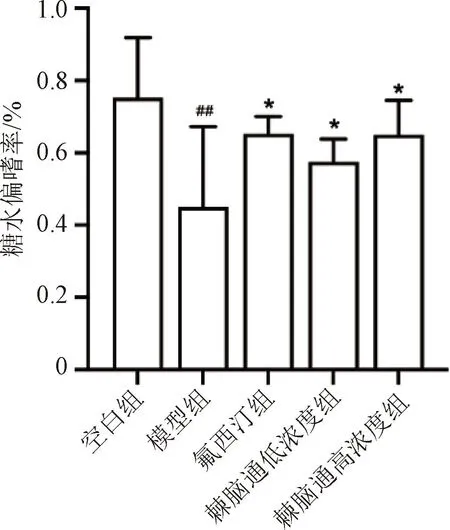
与空白组比较,##P<0.01;与模型组比较,*P<0.05。
3.2 棘脑通对CUMS模型小鼠体重的影响
与空白组相比,模型组小鼠体重显著降低(P<0.01),棘脑通给药组、氟西汀组小鼠体重较模型组显著升高(P<0.01),差异均有统计学意义;棘脑通给药组、氟西汀组小鼠体重的差异无统计学意义(P>0.05),见图2。

与空白组比较,##P<0.01;与模型组比较,**P<0.01。
3.3 棘脑通对CUMS小鼠悬尾不动时间的影响
与空白组相比,模型组小鼠悬尾不动时间显著延长(P<0.01);棘脑通低、高浓度组小鼠悬尾不动时间较模型组显著缩短(P<0.01、P<0.05),上述差异均有统计学意义,提示棘脑通可改善抑郁小鼠的绝望行为,见图3。

与空白组比较,##P<0.01;与模型组比较,*P<0.05,**P<0.01。
3.4 棘脑通对CUMS小鼠强迫游泳不动时间的影响
与空白组比较,模型组小鼠强迫游泳不动时间显著延长(P<0.01);棘脑通给药组小鼠强迫游泳不动时间较模型组显著缩短(P<0.01),差异均有统计学意义,提示棘脑通可改善抑郁小鼠的行为绝望状态,见图4。
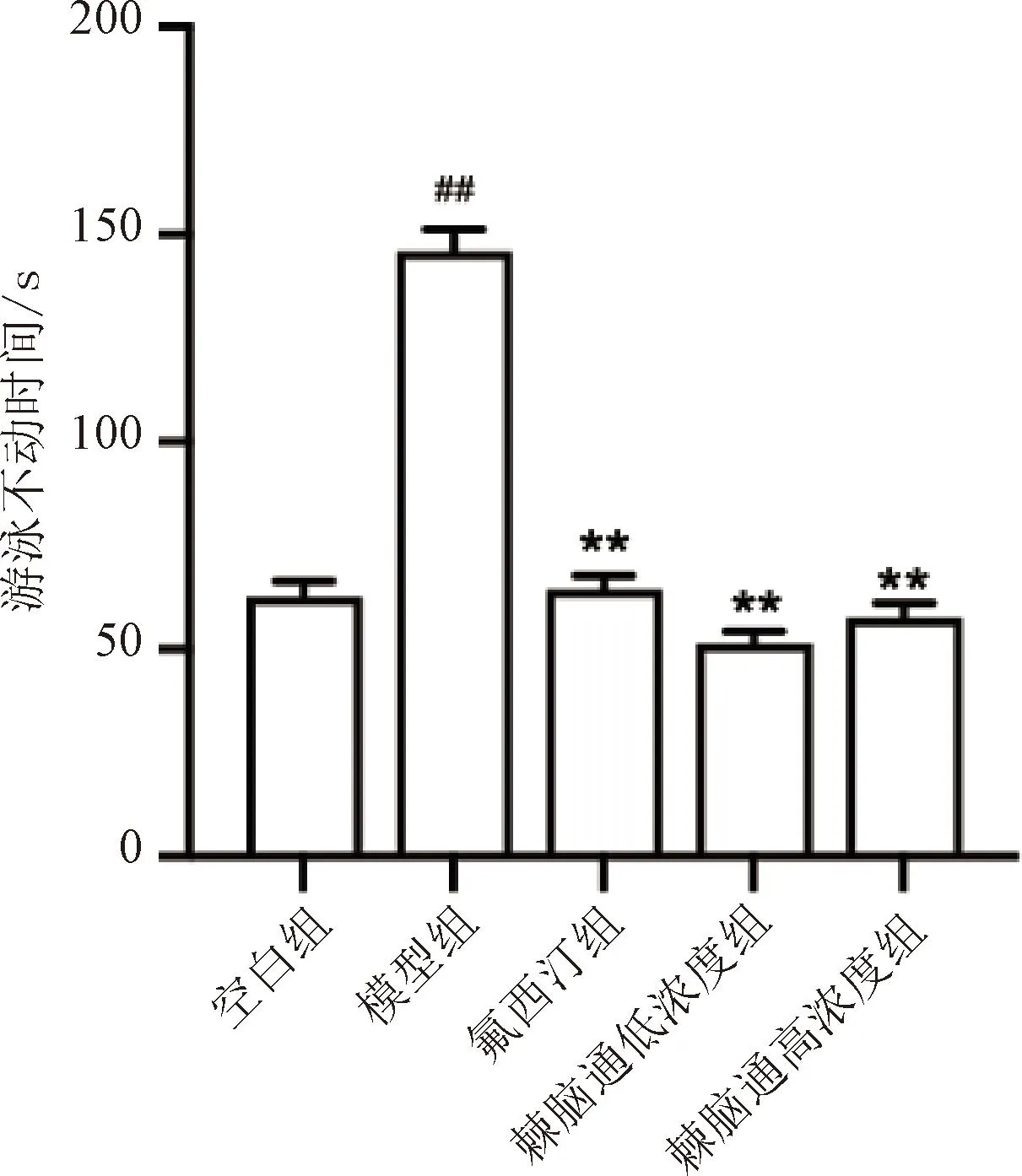
与空白组比较,##P<0.01;与模型组比较,**P<0.01。
3.5 棘脑通对NGF诱导PC12细胞分化和神经微丝表达的作用
不同浓度的棘脑通(0、10、30、100 μg/mL)在单独使用时对PC12细胞分化或对pNF200-Luc表达荧光素酶的诱导作用不明显。当棘脑通与NGF(5、50 ng/mL)协同使用时,PC12细胞被诱导分化的程度及pNF200-Luc表达荧光素酶活性被诱导提高的程度均显著高于相应浓度NGF单独使用时的效果,分化细胞比例及荧光素酶活性的提高百分比或倍数呈浓度依赖形式,提示棘脑通可协同NGF促进神经分化和诱导神经微丝的表达,见图5—6。
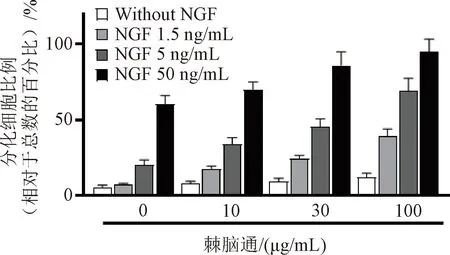
图5 不同浓度棘脑通单独或协同不同浓度NGF诱导PC12细胞分化的作用

图6 不同浓度棘脑通单独或与不同浓度NGF诱导pNF200-Luc表达荧光素酶的作用
3.6 棘脑通协同BDNF、NT-3、NT-4促进神经微丝表达的作用
棘脑通(100 μg/mL)及低剂量(5 ng/mL)神经营养因子在单独使用时,均可分别诱导pNF200-Luc荧光素酶活性提高2~3倍。棘脑通与BDNF、NT-3和NT-4(5 ng/mL)协同使用时,荧光素酶的活性显著提高,其中棘脑通与BDNF(5 ng/mL)联用时可使荧光素酶活性提高8倍,提示棘脑通可协同神经营养因子(BDNF、NT-3、NT-4)诱导神经微丝的表达,见图7。
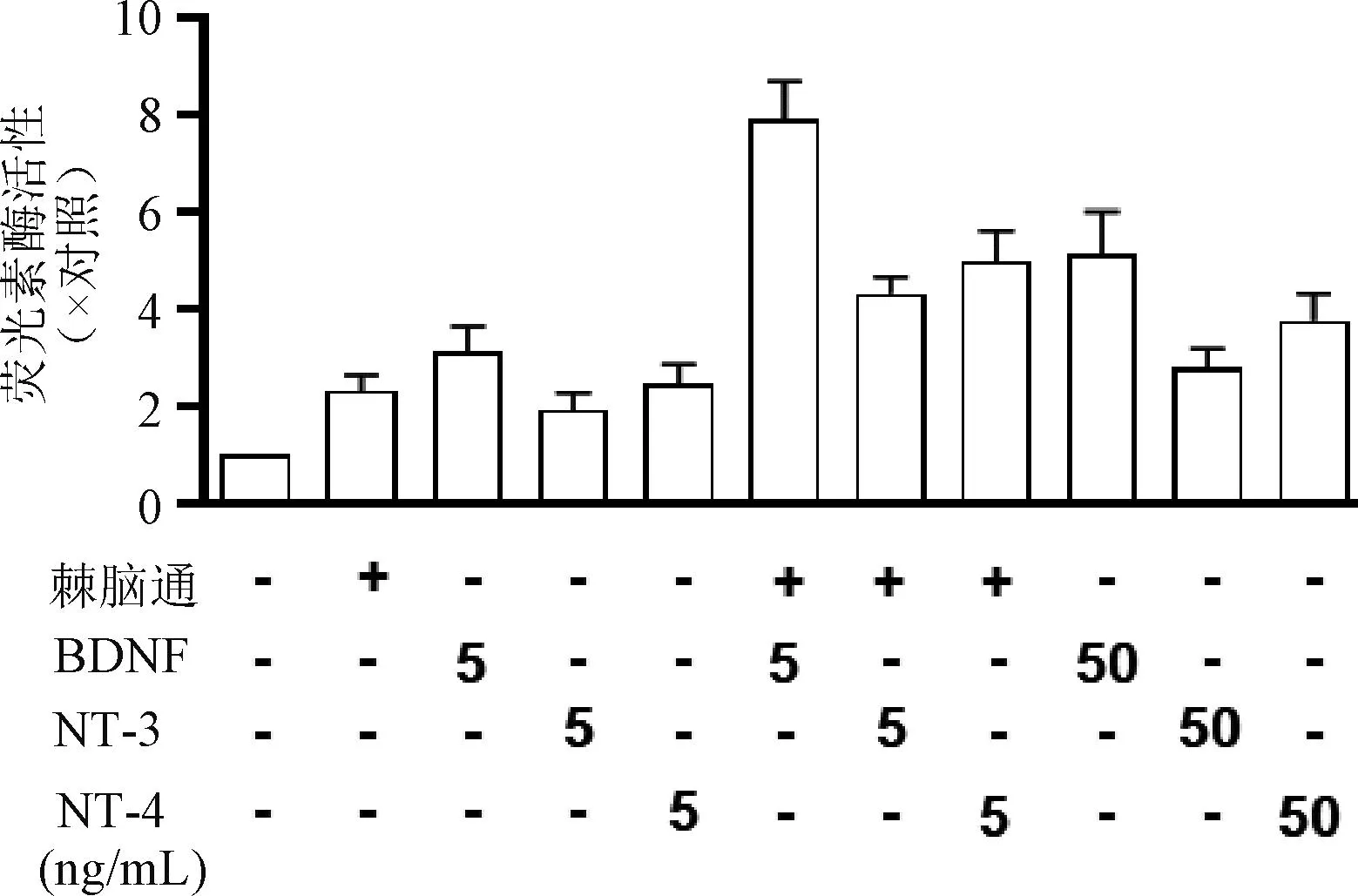
图7 棘脑通与不同神经营养因子协同诱导pNF200-Luc表达荧光素酶的结果
3.7 棘脑通对Aβ诱导神经损伤的作用
棘脑通在100~1 000 μg/mL的浓度范围内可显著降低Aβ的聚集;50 μg/mL的棘脑通可使得PC12细胞活性恢复至50%,与姜黄素的效果相同,提示棘脑通具有抑制Aβ聚集及保护神经细胞对抗Aβ造成的神经毒性损伤的作用,见图8。
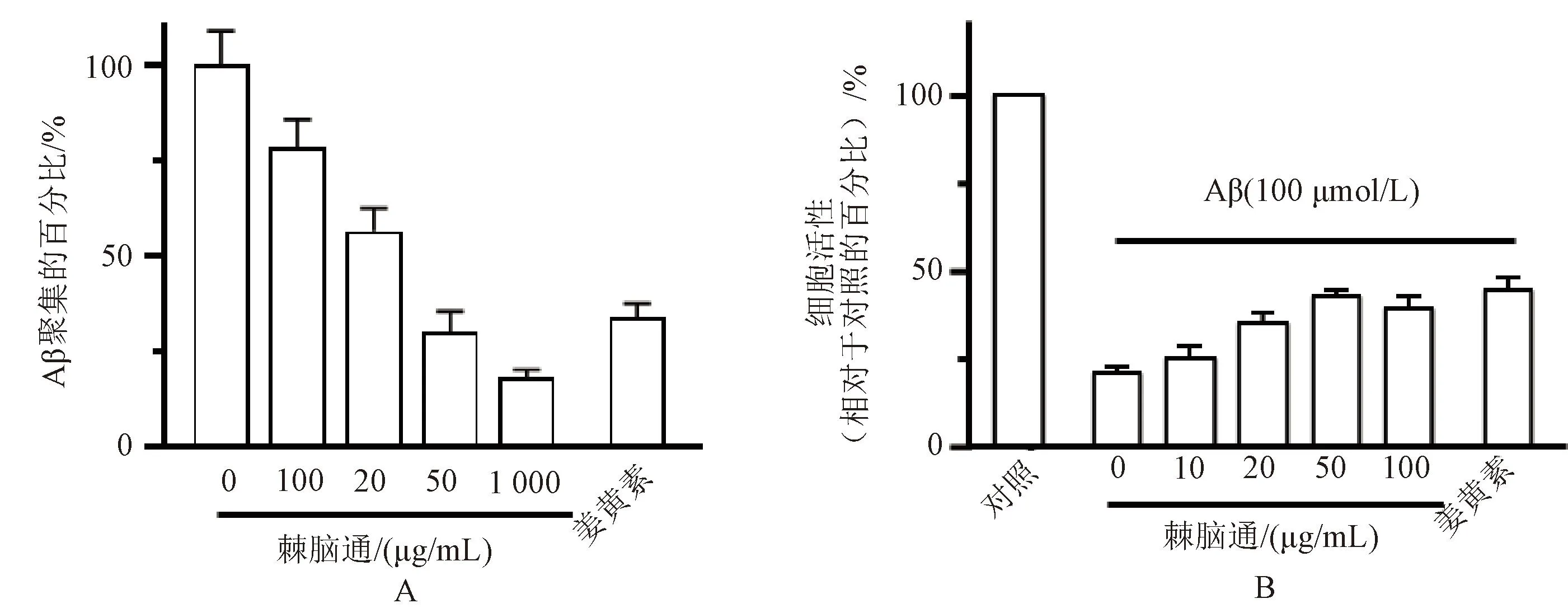
A.棘脑通对Aβ聚集的作用;B.棘脑通对Aβ诱导神经细胞损伤的作用。
3.8 棘脑通对抗氧化应激诱导神经损伤的作用
采用tBHP诱导PC12细胞的氧化损伤后,应用棘脑通可提高PC12细胞活性,且浓度越高,效果越好,在浓度为100 μg/mL时几乎可以使细胞活性恢复至对照组水平;采用Erastin诱导SH-SY5Y细胞铁死亡后,应用棘脑通可提高SH-SY5Y细胞活性,且浓度越高,效果越好,在浓度为100 μg/mL时可达到与阳性对照黄芩素相近的保护效果,提示棘脑通具有保护细胞、抑制氧化损伤及铁死亡损伤的潜在药用功能,见图9。

A.棘脑通对tBHP诱导损伤的细胞保护作用;B.棘脑通对Erastin诱导损伤的细胞保护作用。

A.抑制IL-1β的表达水平;B.抑制NF-κB转录活性水平。
3.9 棘脑通的抗炎作用
本研究在BV2、RAW 264.7细胞上考察了不同浓度的棘脑通抑制LPS引起的炎症损伤的作用。提示棘脑通可抑制脂多糖诱导的炎症损伤。将LPS用于BV2、RAW 264.7细胞后,应用棘脑通可降低IL-1β的mRNA水平、NF-κB转录活性,且具有浓度依赖性,见图10。
4 讨论
抑郁症是一种精神情感障碍,主要表现为情绪低落、快感和兴趣丧失、疲劳感和精力不济等,常伴有躯体和认知变化[11]。2008年,世界卫生组织将重度抑郁症列为全球疾病负担的第三大原因,预计2030年重度抑郁症将居全球疾病负担的首位。目前,关于抑郁症病理机制的研究较多,主要包括单胺假说、下丘脑-垂体-肾上腺轴变化、炎症因子、神经可塑性和神经发生、大脑结构和功能变化、基因、环境因素及表观遗传学等,但目前尚无任何单一模型或机制能够解释抑郁症发病的所有方面[12]。中医药基于整体观出发,发挥中药复方多成分、多靶点的优势,通过调节神经递质、抑制神经炎症与保护肠道屏障等环节发挥整体调节作用,实现其抗抑郁功能[13]。中医认为,抑郁症属于“郁证”范畴。《黄帝内经》中提及“愁忧者,气闭塞,而不行”,提出如若情绪忧郁,则易致身体气机不畅,“思则心有所存,神有所归,正气留而不行,故气结”,提出了引起机体气机失调的重要原因是情志因素[14]。
沙棘(Hippophaerhamnoides L.)属于胡颓子科沙棘属的落叶性灌木或乔木,具有药食同源性。沙棘营养丰富,含有维生素E、维生素C、维生素K、维生素A、维生素B、维生素P,氨基酸,脂肪酸(饱和、不饱和脂肪酸),有机酸,挥发油,类胡萝卜素,甾类化合物,黄酮,多酚,植物蛋白和脂肪等多种营养成分。在沙棘的各种营养成分中,黄酮类化合物是主要功能性成分之一,具有抗氧化活性、抗炎活性、抑制肿瘤生长、改善心血管疾病和改善肾脏纤维化等多种生理活性[15]。
花生是豆科落花生属一年生草本植物,是全球最重要的四大油料作物之一。我国花生种植面积逐年递增,产量已超过1 500万吨,位居世界第一。花生壳是花生果加工成食品和食用油脂后的废弃物,除含有大量纤维素、木质素外,还含有少量的蛋白质、脂肪、多糖、黄酮类物质以及矿物质等营养成分。花生壳提取物的有效成分是以木犀草素为主的黄酮类化合物。现代研究表明,花生壳黄酮具有降脂减肥、抗自由基、抗氧化、抑菌、抗病毒和抗肿瘤等作用[16-17]。花生壳中纤维素占比高达38.9%,因此,花生壳廉价易得,且可用于生产膳食纤维。膳食纤维应用在食品中,可以提高人们的膳食水平。膳食纤维还具有降低患癌风险、预防结肠疾病、降低血液胆固醇浓度、控制体重和降低心血管疾病发病率等生理活性[18]。因此,对于花生壳的开发利用不仅有很大的生物学价值,也有丰富的资源再利用价值。
沙棘和花生均属药食同源类中药材,作为民族医药的应用已有很多年的历史,但现今对其的药理学研究大多局限在抗糖、抗炎和抗氧化作用等,较少有关于二者合用治疗神经退行性疾病的报道。本研究利用CUMS抑郁小鼠模型,系统研究了以沙棘提取物和花生壳提取物为主要成分的棘脑通的抗抑郁作用。本研究结果证实,棘脑通可逆转CUMS引起的体重减轻,并通过糖水偏嗜、强迫游泳和悬尾等行为学测试证实棘脑通可显著改善抑郁小鼠的绝望情绪和快感缺乏,其疗效与临床使用最广泛的选择性血清素再摄取抑制剂氟西汀相当。为了尝试阐明其抗抑郁作用机制,本研究通过细胞模型,证明了棘脑通能够模拟和增强神经营养因子的功能,包括促进细胞突起生长、激活神经微丝编码基因表达,可抗氧化、抗炎及抗Aβ的作用,并可协同神经营养因子诱导神经细胞分化。以上作用阐明了棘脑通缓解抑郁症状的机制,也提示棘脑通可能对改善多种神经退行性疾病等认知障碍有疗效。
综上所述,本研究从动物及细胞水平上阐释了棘脑通抗抑郁的药效及作用机制,为其临床用于改善抑郁症疾病提供了支撑,也为抗抑郁产品的开发提供了借鉴。同时,本研究为沙棘及花生壳的药理研究提供了新方向,希望能开拓沙棘及花生壳的药用资源应用。
