590份燕麦种质资源营养器官表型性状遗传多样性分析
2024-02-02王凤宇梁国玲刘文辉
王凤宇 梁国玲 刘文辉

doi:10.11733/j.issn.1007-0435.2024.01.017
引用格式:
王鳳宇, 梁国玲,刘文辉.590份燕麦种质资源营养器官表型性状遗传多样性分析[J].草地学报,2024,32(1):158-167
WANG Feng-yu, LIANG Guo-ling, LIU Wen-hui.Genetic Diversity Analysis of Nutritional Organs Phenotype in 590 Oat Germplasms[J].Acta Agrestia Sinica,2024,32(1):158-167
摘要:为明确燕麦(Avena sativa)种质资源营养器官表型性状的遗传多样性和各性状之间的相关关系,进而为燕麦育种和种质资源创新利用提供理论依据,本研究以国内外收集的590份燕麦种质资源为研究对象,在青海省西宁市湟中区对其从地上器官质量性状、茎部性状、叶部性状和分蘖性状的遗传多样性进行评价分析,并利用相关性分析和聚类分析方法,评价其营养器官表型性状的遗传变异水平。结果表明:590份燕麦种质资源质量性状遗传多样性指数以茎节绒毛最高(1.026),苗期株型最低(0.162);数量性状的遗传多样性指数以第三茎节茎粗最高(2.09),旗叶叶面积最低(1.87);变异系数以无效分蘖数(54.92%)最大,茎节数最小(12.20%);茎部性状和倒二叶长、倒二叶叶面积是影响燕麦种子和饲草产量的重要因子。590份燕麦资源可分为4大类群,其中类群III可作为饲草型育种目标的亲本材料,类群IV可作为抗倒伏育种材料。
关键词:皮燕麦;茎部性状;叶部性状;表型性状;遗传多样性
中图分类号:S512.6 文献标识码:A 文章编号:1007-0435(2024)01-0158-10
Genetic Diversity Analysis of Nutritional Organs Phenotype in 590 Oat Germplasms
WANG Feng-yu, LIANG Guo-ling*, LIU Wen-hui
(Key Laboratory of Superior Forage Germplasm in the Qinghai-Tibetan Plateau, Qinghai University (Qinghai Province)
Academy of Animal Science and Veterinary Medicine, Xining, Qinghai Province 810016, China)
Abstract:To clarify the genetic diversity of nutritional organ phenotype of oat (Avena sativa) germplasm resources and the correlations among the phenotypic traits,and provide a scientific basis for oat breeding and innovative utilization of germplasm resources,this study evaluated and analyzed the genetic diversity of aboveground organ quality traits,stem traits,leaf traits,and tiller traits of oat in Huangzhong District,Xining City,Qinghai Province. 590 oat germplasm resources collected from home and abroad was evaluated using the methods of correlation analysis and cluster analysis The results showed that the genetic diversity index of qualitative traits was the highest in the node hairiness (1.026),while the lowest in seedling type (0.162). The genetic diversity index of quantitative traits was the highest in the third internode diameter (2.09),while the lowest in flag leaf area (1.87). The coefficient of variation was the largest in tiller number (54.92%),while the smallest in internode number (12.20%). The traits related to stem characteristics and the length of penultimate leaf and the area of penultimate leaf were important factors influencing oat seed and forage yield. The 590 oat germplasm resources were divided into four clusters by cluster analysis. Cluster III could be used as parent materials for forage breeding goals,and Cluster IV could be used as lodging-resistant materials for breeding.
Key words:Covered oats;Stem traits;Leaf traits;Phenotypic traits;Genetic diversity
收稿日期:2023-05-11;修回日期:2023-07-16
基金项目:青海省科技厅帅才科学家负责制项目(2023-NK-147);青海省高端人才千人计划;现代农业产业技术体系建设(CARS);青海省“昆仑英才·科技领军人才计划”共同资助
作者简介:
王凤宇(1998-),女,汉族,山东济宁人,硕士研究生,主要从事草学研究,E-mail:wangfy512@163.com;*通信作者Author for correspondence,E-mail:qhliangguoling@163.com
燕麦(Avena sativa)为禾本科(Poaceae)燕麦属(Avena)植物,在甘肃、内蒙古、青海、四川、东北等地广泛种植[1]。燕麦喜冷凉、耐寒、耐贫瘠、耐盐碱、抗旱、抗杂类草[2-3],具有生产性能好、家畜喜食、饲用价值高和营养全面等优点[4],是公认的高产、稳产和营养价值高的优质饲草料[5]。在青藏高原地区,燕麦是人工草地建植中应用最广泛的优质饲草[6],在维系青藏高原高寒牧区草地畜牧业可持续发展中发挥了重要作用。前人已对青藏高原高寒区燕麦从引种[7]、资源评价[6]、栽培技术[8-9]和新品种选育[10-11]等方面做了大量工作,有效推动了燕麦产业的发展。但不同生态区域和环境因子对燕麦生长发育的影响、种植栽培技术及品种的要求却不尽相同。同时,随着国家对种业的重视,有关青藏高原高寒区燕麦方面的研究也越来越深入,迫切需要加强对燕麦种质资源的挖掘利用和新品种的选育工作,以满足我国燕麦种业的发展。因此,在燕麦种质资源研究、资源评价和品种选育中,应因地制宜并结合当地生产需求,制定切实可行的评价体系、育种目标、技术路线和实施方案等,为选育高产、适应性强和不同需求的专用型燕麦品种提供科学依据,促进燕麦产业化发展进程,为高寒区畜牧业发展提供优质燕麦饲草,实现农牧民增产增收。
开展燕麦种质资源遗传多样性研究,有利于燕麦优异和特异种质资源的挖掘与创新,对燕麦资源鉴定、性状评价和育种等具有重要的指导意义[12-13]。表型鉴定具有直观、简便易行和成本低的优点,不仅是鉴定评价资源的首要选择,也是细胞学、生物化学和分子标记鉴定的基础。王建丽等[14]对来自不同地区51份皮、裸燕麦从13个农艺性状进行形态多样性分析发现,主穗长、株高和主穗粒重遗传多样性指数较高,主穗小穗数、主穗粒重和单株分蘖数变异系数最大;王娟等[15]在山西地区对39份燕麦的研究表明,15个表型性状中粒型和单株粒重遗传多样性指数较高,叶相遗传多样性指数较低;贾志锋等[16]在青海地区对100份皮、裸燕麦的研究指出,主穗小穗数、芒色和粒形的遗传多样性指数较高,株高和主穗轮层数的变异系数最小,上数第2片叶长和叶宽的变异系数较大;张琦等[17]在江淮地区对141份燕麦的研究指出,单株鲜重、单株干重和千粒重多样性指数较高,千粒重和穗粒数变异系数较大。上述研究发现燕麦种质资源具有丰富的遗传多样性和变异类型,这些研究对燕麦资源评价和育种等方面具有一定的指导意义,但上述研究选择的燕麦群体较小,且选择的评价指标、生态区域和种质来源不同,得出的结论也不尽相同,同时在表型性状遗傳多样性分析中多集中于穗部性状,对叶部和茎部器官表型性状的关注相对较少。叶片和茎秆作为植物地上部重要的养分输送和光合作用有机物制造器官[18-21],有必要对其表型性状进行详细研究,为高寒区燕麦种质资源引种、资源评价和品种选育等提供理论依据。
因此,本研究以来源于全球不同区域的590份燕麦为研究对象,系统评价燕麦叶部和茎部的形态特征,阐明青藏高原高寒区燕麦叶片和茎秆形态特征遗传多样性。旨在通过研究,明确燕麦种质资源叶部和茎部形态性状的表型差异及适应性变异,为高寒区燕麦种质资源筛选、评价和品种选育等提供科学依据。
1 材料与方法
1.1 试验材料
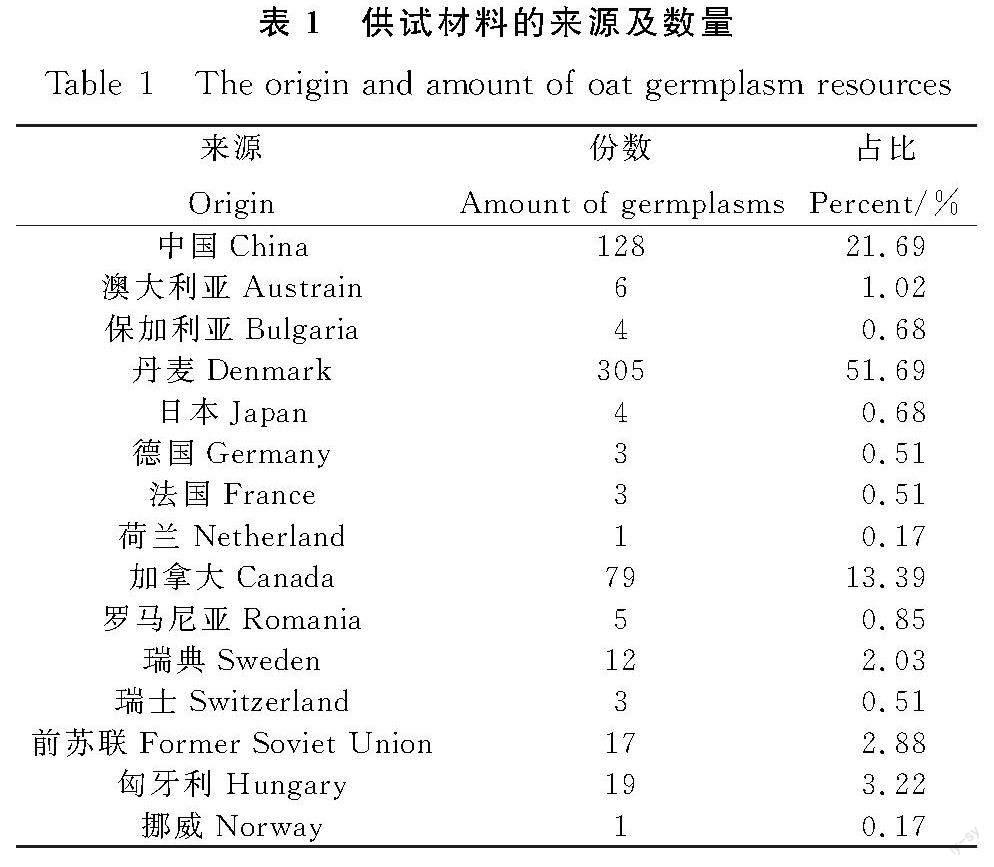
供试燕麦材料来源于中国、加拿大、丹麦和匈牙利等15个国家的种质资源,共计590份,均为皮燕麦,供试材料来源及数量见表1。以上材料均由青海省畜牧兽医科学院提供。
1.2 试验设计与方法
1.2.1 试验设计 试验地位于青海省燕麦生产主产区和优势产区的西宁市湟中区甘河滩镇下营村(101°33′20″ E,36°30′57″ N),海拔2 592 m。试验区域地势平坦,前茬为油菜(Brassica napus),灰钙土。小区面积为2 m×3 m,播种方式为条播,行长3 m,行距25 cm;种子播深为3~4 cm,4月下旬进行播种。小区播量根据每份种质材料的千粒重、发芽率和纯净度,按照525万株·hm-2保苗数计算其实际播量,小区间距50 cm,不设重复。以磷酸二铵(150 kg·hm-2)和尿素(75 kg·hm-2)作基肥,旱作,分蘖期田间除草一次。
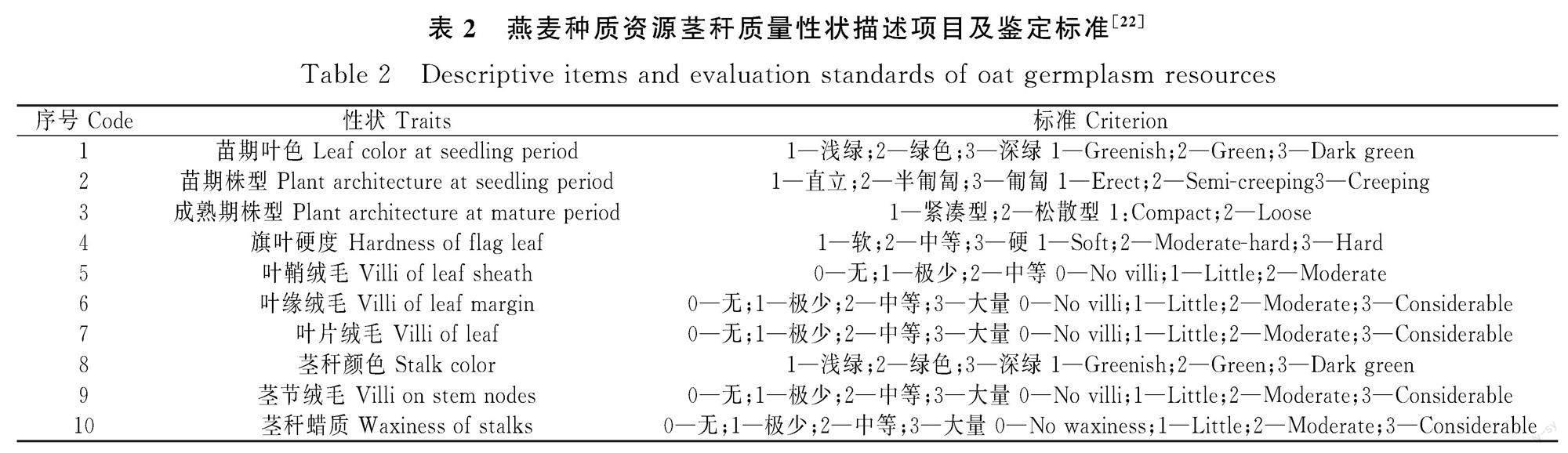
1.2.2 评价方法 农艺学性状测定分别在分蘖期、开花期和完熟期进行观测和取样。性状调查参照《燕麦种质资源描述规范和数据标准》[22]进行,描述的性状有苗期叶色和株型,花期旗叶硬度、叶鞘绒毛、叶缘绒毛、叶片绒毛、茎秆颜色、绒毛和蜡状物以及成熟期株型共10个质量性状并分别予以赋值(表2)。在成熟期时测定株高、分蘖数、叶片数、旗叶长、旗叶宽、倒二叶长、倒二叶宽、茎节数、基部第二和第三茎节的茎粗、旗叶至花序基部长和倒二叶至花序基部长等14个数量性状。
1.3 数据处理及统计分析
数据采用Excel 2019分析各材料数量性状的数据;利用SPSS 19.0软件进行相关性分析、主成分分析(PCA)和聚类分析(CA),其中聚类方法采用离差平方和法(Wards method),种质间遗传距离采用欧式距离[20-21];采用Origin 8.0软件绘图;利用Popgene 1.32 软件计算遗传多样性指数(H′)。
2 结果与分析
2.1 地上营养器官性状变异及分布
2.1.1 质量性状遗传多样性分析 对590份皮燕麦10个质量性状的遗传多样性分析(图1)发现,茎节绒毛遗传多样性指数最高(1.026),说明该性状具有稳定的遗传性。其次是茎秆颜色、茎秆蜡质和苗期叶色遗传多样性指数,都在0.951以上。苗期株型和叶鞘绒毛遗传多样性指数相对较低,分别为0.162和0.202。各表型性状遗传差异较大,存在丰富的遗传多样性。10个质量性状中遗传多样性指数大小依次为茎节绒毛>茎秆颜色>茎秆蜡质和苗期叶色>叶缘绒毛>旗叶硬度>叶片绒毛>成熟期株型>叶鞘绒毛>苗期株型。
2.1.2 质量性状变异及分布 由燕麦地上营养器官10个质量性状分布类型和分布频率(图2)可知,燕麦苗期叶色以绿色为主(56.8%),其次为深绿色,而少量的燕麦资源苗期颜色呈浅绿色(13.4%);从株型来看,苗期直立型最多(96.8%),仅有极少部分燕麦呈匍匐(2.2%)或半匍匐(1.0%)状态;成熟期株型以紧凑型(78.0%)为主,其余为松散型;大多数燕麦资源旗叶硬度中等(65.4%),并由一部分呈硬(28.1%)的状态,只有极少部分呈柔软状态(6.4%);多数资源无叶鞘和叶缘绒毛,分别约占资源总数的95.6%和72.4%;叶片有中等量绒毛的资源约占78.5%,仅少量资源有大量绒毛或无;茎秆颜色以绿色居多,约占56.1%,且有部分资源茎秆呈深绿和浅绿色;茎节上无绒毛资源约占51.7%,中等量约占36.8%,绒毛较多或极少的资源相对较少;茎秆上有适中量蜡状物的资源约占84.2%,无蜡状物覆盖的资源极少。
2.1.3 数量性状变异及遗传多样性 对燕麦地上部营养器官16个数量性状的变异情况进行分析(表3),结果显示,平均变异系数23.57%,平均遗传多样性指数2.02,该资源各数量性状具有丰富的多样性。茎部特性中变异系数较大的包括第二茎节茎粗(16.26%)和第三茎节茎粗(16.04%),第二茎节茎粗和第三茎节茎粗范围分别为1.63~5.29 mm,1.84~5.62 mm,茎部特性中茎粗的变异最大,较其他性状而言稳定性差;最小的是茎节数(16.26%),茎节数为3.40~7.00节。分蘖特性中变异系数以无效分蘖数最高(54.92%),分蘖数为0~4.20个,以总分蘖数最小,为30.38%。叶部特性中以旗叶叶面积变异系数最高(38.73%),其次为倒二叶叶面积(32.88%);较小的包括叶片数(12.93%)和倒二叶长(16.84%)。变异系数由大到小依次为:无效分蘖数>旗叶叶面积>有效分蘖数>倒二叶叶面积>总分蘖数>旗叶宽>倒二叶宽>旗叶长>倒二叶长>第二节茎粗>第三节茎粗和旗叶至花序基部长>株高>倒二叶至花序基部长>叶片数>茎节数。茎部特性的遗传多样性指数比其他两个特性的遗传多样性指数大,其遗传多样性指数都在2.02以上,叶部特性的遗传多样性指数最小;茎部性状的遗传基础最广,叶部性状的遗传基础较窄。遗传多样性指数由大到小依次为:第三茎节茎粗>株高>旗叶至花序基部长>第二茎节茎粗>无效分蘖数、倒二叶长和倒二叶至花序基部长>叶片数和倒二叶宽>总分蘖数和茎节数>旗叶长、旗叶宽和倒二叶叶面积>有效分蘖数>旗叶叶面积。
2.1.4 数量性状的分布 16个数量性状次数分布柱形图表明(图3),各数量性状分布间存在差异。经分布函数X2检验,株高、總分蘖数、有效分蘖、叶片数、旗叶长、旗叶宽等16个数量性状均符合正态分布。其中70%以上的资源株高在85~120 cm之间,茎节数大部分为4.5~5节和5.5~6节,50%以上的材料植株都比较高大。旗叶和倒二叶至花序基部长分别在35~40 cm,60~70 cm之间的资源较多,一半以上资源的茎粗都集中在3.0~3.5 mm之间。由下图可知该资源中多数资源叶片数集中在4.5~5.5片,且叶面积较大。
2.2 相关性分析
燕麦地上营养器官农艺性状相关性分析显示(表4),燕麦种子产量与茎部特性均呈极显著正相关关系(P<0.01),与叶部特性中的倒二叶长和倒二叶叶面积也呈极显著正相关关系(P<0.01);燕麦饲草产量与茎部特性以及叶部特性的叶片数、倒二叶长和倒二叶叶面积呈极显著正相关关系(P<0.01);表明茎部性状和叶部性状中的倒二叶长与倒二叶叶面积对产量影响较大。其中各茎部性状间均存在显著(P<0.05)或极显著(P<0.01)正相关关系,这些性状中无论提高哪一个性状其他性状都会随着该性状的提高而提高。表型性状与燕麦饲草产量相关性大小顺序为株高>叶片数>茎节数>倒二叶至花序基部长>倒二叶长>叶面积>旗叶宽。有效分蘖数和无效分蘖数分别与总分蘖数之间呈极显著正相关关系(P<0.01)。叶部特性中除了叶片数与旗叶长、旗叶宽以及倒二叶宽之间相关关系不显著,剩余其他叶部性状之间都存在显著正相关关系(P<0.05)或显著负相关关系(P<0.05)。株高、倒二叶花序至基部长以及茎粗与茎部特性、分蘖特性和叶部特性之间都存在显著相关关系(P<0.05),这3个性状对燕麦的影响最大。
2.3 主成分分析
16个数量性状主成分构成中主要信息集中在前4个主成分中(表5),特征值均大于1,累计贡献率78.514%。第一主成分的特征值6.430,贡献率40.815%,特征向量中除总分蘖数和有效分蘖数载荷为负外,其余载荷均为正。载荷值在0.800以上的性状包括倒二叶长(0.815)、倒二叶叶面积(0.888)、茎基部第三茎节茎粗(0.814),这些性状与饲草产量和抗倒伏性有关,可作为抗倒兼草产量高的品种进行选育。第二主成分特征值为3.142,贡献率19.635%,载荷较高的性状有总分蘖数、有效分蘖数和旗叶叶面积,特征向量值分别为0.524,0.542和0.479,从载荷数值可看出,分蘖数和旗叶叶面积对种子产量具有重要作用。第三主成分的特征值为1.781,贡献率11.129%。载荷较高的性状是总分蘖数,特征值为0.802,对饲草产量有较大影响。第四主成分特征值为1.211,贡献率7.566%,特征向量中有正有负。载荷较高的性状是无效分蘖数、叶片数和茎节数,分别为0.477,0.439和0.430,说明这些性状对燕麦的草产量和抗倒伏性有影响。
2.4 聚类分析
对590份燕麦种质资源茎部和叶部性状进行聚类分析,发现在欧式距离为5~7处可将参试材料聚为4大类群(表6)。第I类群包含106份材料,约占资源总数的18.0%,这一类群品种特征为株高最矮(79.13 cm)茎粗最细,叶量较少,分蘖数最多,其他性状表现一般。第II类群包含265份材料,约占资源总数的44.92%,该类群主要特征是株高较高(107.49 cm)茎粗较粗,变异系数都比较小,茎节数平均为5.52个,叶量较丰富,且变异系数较大,这一类群植株综合性状较好,可作为一般种质资源进行保存。第III类群包含103份材料,约占总资源的17.46%,这一类群的品种植株高大(124.64 cm),茎粗最粗,且变异系数均最小,茎节数最多(5.90个),叶片数平均为5.22个、旗叶和正二叶叶面积较大,叶量丰富,分蘖数较多,该类群具有较好的饲草生产性能,可以对其进行饲草型燕麦品种的筛选。第IV类群包含116份材料,约占资源总数的19.66%,该类群品种主要特征表现为株高较矮,茎粗较粗,有效分蘖数也比较多,可作为选育抗倒伏育種材料。茎部性状是影响抗倒伏性能的关键因素,茎秆不仅支撑植株重量,还是被外界环境直接作用的组织器官。
3 讨论
3.1 燕麦种质资源营养器官表型遗传多样性
表型性状鉴定和描述是种质资源研究中最基本和最便捷的方法,590份燕麦种质资源营养器官表型性状的遗传多样性分析表明,质量性状中成熟期株型以占总资源的78.0%。由此可以发现国外引进的皮燕麦资源在株型上主要为紧凑型。这与雷雄等[23]研究结果相一致,而与王建丽等[14]对黑龙江地区种植的51份皮、裸燕麦研究结果不一致,造成差异的原因可能是王建丽研究的燕麦群体较小,生态区域和种质来源不同所致。本研究发现16个质量性状中,有效分蘖数、无效分蘖数和叶面积变异系数最大。这与王娟等[15]、武敬也等[24]和耿小丽等[25]的研究结果一致。分蘖数和叶面积受环境影响较大,变异类型丰富,可以通过环境和栽培措施进行品种改良,发挥其潜在的生产性能。对数量性状进行遗传多样性分析时,发现多样性指数较高是茎部性状的茎粗、株高、旗叶至花序基部长和分蘖特性的无效分蘖数。这与张琦等[17]和周启龙等[26]研究结论基本一致,表明株高和茎粗遗传变异较丰富,对环境适应能力强。不同燕麦种质资源的株高、茎粗、分蘖数和叶面积具有较大的表型差异和丰富的遗传多样性,可为青藏高原地区燕麦种质资源的科学分类提供借鉴,也可为不同类型的燕麦品种选育提供种质材料。
3.2 燕麦种质资源营养器官与产量之间的相关性分析
目前有关燕麦表型遗传多样性研究较多的是对生殖器官穗部性状的分析[13,27-28],大多数研究人员认为燕麦种子产量受穗部性状影响较大[27,29]。但叶片和茎秆是植物地上部分最重要的营养器官,其中叶片是植物重要光合器官,有研究发现叶片对高粱、晚稻、谷子等植物产量的形成有重要的影响[30-32];茎是植株输送水分和营养的主要器官,对燕麦的饲草产量也有一定影响[6]。通过对燕麦种子产量和饲草产量与表型性状进行分析,可以发现促进和限制燕麦种子和饲草增产的主要性状,为今后生产提高产量提供理论依据。本研究发现,燕麦种子产量与部分茎部性状(株高、叶片至花序基部长和茎节数等性状)呈极显著正相关关系(P<0.01);也与叶部特性中的倒二叶长和倒二叶叶面积呈极显著正相关关系(P<0.01)。耿小丽等[25]研究也发现株高、茎粗、叶片数等因素对种子产量有较大影响。孙建平等[33]对饲用燕麦品种“牧王”研究发现饲草产量与株高、茎粗和叶面积等性状呈极显著(P<0.01)正相关关系;李晶等[34]研究发现,饲草产量与株高、叶片数呈极显著(P<0.01)正相关关系;本研究也有类似的结果。燕麦茎部性状(株高、叶片至花序基部长和茎节数等性状)和叶部性状(倒二叶长和倒二叶叶面积)是影响燕麦种子产量和饲草产量的重要影响因子。
3.3 不同用途燕麦种质资源分类
聚类分析已成为牧草育种研究常用的方法之一[35]。本试验按照k-均值(k-mean)法在欧式距离为5~7时划分为4大类群。在4大类群中,第I类群材料潜在生产性能整体表现一般。第II类群包含材料最多(265份),该类群植株综合性状较好,可作为一般种质资源进行保存。第III类群植株与饲草产量相关的性状相比其他类群的性状表现最优,可作为选育饲草型燕麦品种材料。研究发现茎叶比是衡量牧草质量的重要指标,可以反映燕麦饲草的品质和适口性,茎叶比越小,适口性越好[27]。大量研究表明,株高是构成燕麦饲草产量的重要部分,分蘖数也是影响饲草产量的一个重要因素,分蘖数的增加可以提高作物的饲草产量[36];燕麦饲用潜力较大的品种一般表现为茎秆粗壮、叶片较大、分蘖较多[37]。这与雷雄等[23]研究结果相一致,也是将植株高大、茎秆粗壮、叶量丰富的资源聚集在一起。株高、基部节间长和茎粗等表型性状是影响燕麦抗倒伏的重要性状,是燕麦抗倒伏评价中关键的参考指标[38-41]。纪明雪等[42]研究发现株高和茎基部第二节间长越短,抗倒伏能力越强。梁国玲等[43]发现抗折力越大、第二节间茎粗越粗,抗倒伏能力越强。研究发现,燕麦茎秆矮且粗,坚韧性会更强,重心高度降低,负荷轻,植株的抗倒性能力便会加强[44-45]。第IV类群,有关抗倒伏性的性状(茎部性状)表现较为突出,可作为选育抗倒伏育种材料。王娟等[15]对燕麦进行聚类分析也发现,株高较矮、茎粗较粗的材料聚集在一起。通过对590份燕麦资源茎部和叶部表型性状的遗传多样性评价,发现燕麦资源丰富,不同来源的燕麦种质资源均在高寒区表现出较好的适应性及生产性能。因此,在今后燕麦品种选育过程中,应该加大国外和国内不同地区的燕麦引种,来丰富我国高寒区燕麦种质资源,为燕麦品种选育和种质创新提供优异资源。
4 结论
590份国内外燕麦种质资源营养器官表型性状具有丰富的遗传多样性,质量性状中以茎节绒毛的多样性指数最高,为1.026,苗期株型最低,为0.162。数量性状中第三茎节茎粗的遗传多样性指数最高(2.09),旗叶叶面积的遗传多样性指数最小(1.87)。变异系数最大的是无效分蘖数,最小的是茎节数。相关性分析发现,茎部性状和倒二叶长、倒二叶叶面积是影响燕麦种子和饲草产量的重要因子。通过聚类分析将590份燕麦材料分为4大类群,类群III作为饲草型育种目标的亲本材料,类群IV可作为抗倒伏育种材料。
参考文献
[1]张向前,刘景辉,齐冰洁,等. 燕麦种质资源主要农艺性状的遗传多样性分析[J]. 植物遗传资源学报,2010,11(2):168-174
[2]贾志锋. N、P对裸燕麦生产性状的影响[J]. 青海畜牧兽医杂志,2013,43(4):8-10
[3]刘凯强,刘文辉,贾志锋,等. 不同播量、行距及播种方式对青燕1号燕麦饲草产量的影响[J]. 草地学报,2019,27(4):1060-1067
[4]李春喜,叶润荣,周玉碧,等. 高寒牧区不同燕麦品种饲草产量及品质的研究[J]. 草地学报,2014,22(4):882-888
[5]赵桂琴,师尚礼. 青藏高原饲用燕麦研究与生产现状、存在问题与对策[J]. 草业科学,2004,21(11):17-21
[6]周青平,颜红波,梁国玲,等. 不同燕麦品种饲草和籽粒生产性能分析[J]. 草业学报,2015,24(10):120-130
[7]郑殿升,张宗文. 中国燕麦种质资源国外引种与利用[J]. 植物遗传资源学报,2017,18(6):1001-1005
[8]刘凯强,刘文辉,魏小星,等. 不同播量和行距对‘青燕1号燕麦种子产量的影响[J]. 草业学报,2020,29(2):82-91
[9]徐长林. 高寒牧区燕麦丰产栽培措施的研究[J]. 草业科学,2003,20(3):21-24
[10]周青平,顔紅波,梁国玲,等. 早熟高产燕麦新品种青燕1号选育报告[J]. 西南民族大学学报(自然科学版),2014,40(2):161-167
[11]刘彦明,李朴芳. 旱地燕麦新品种定莜6号的选育及其特征分析[J]. 干旱地区农业研究,2010,28(5):1-4
[12]梁国玲,刘文辉,马祥. 590份皮燕麦种质资源穗部性状遗传多样性分析[J]. 草地学报,2021,29(3):495-503
[13]齐冰洁. 燕麦种质资源遗传多样性研究[D]. 呼和浩特:内蒙古农业大学,2009:1-2
[14]王建丽,马利超,申忠宝,等. 基于遗传多样性评估燕麦品种的农艺性状[J]. 草业学报,2019,28(2):133-141
[15]王娟,李荫藩,梁秀芝,等. 北方主栽燕麦品种种质资源形态多样性分析[J]. 作物杂志,2017,(4):27-32
[16]贾志锋,马祥,雷生春,等. 燕麦种质资源在青海生态区的形态多样性评价[J]. 种子,2019,38(12):58-63
[17]张琦,魏臻武,闫天芳,等. 燕麦种质资源农艺性状遗传多样性的鉴定评价[J]. 草地学报,2021,29(2):309-316
[18]钟悦鸣,董芳宇,王文娟,等. 不同生境胡杨叶片解剖特征及其适应可塑性.[J]. 北京林业大学学报,2017,39(10):53-61
[19]于海燕,胡潇予,何春霞,等. 文冠果不同种源叶片结构对水分胁迫的差异性响应[J]. 北京林业大学学报,2019,41(1):53-61
[20]WRIGHT I J,REICH P B,WESTOBY M,el al. Strategy shifts in leaf physiology,structure and nutrient content between species of high- and low-rainfall and high- and low-nutrient habitats[J]. Functional Ecology,2001,15(4):423-434
[21]CHARTZOULAKIS K,PATAKAS A,KOFIDIS G,el al. Water stress affects leaf anatomy,gas exchange,water relations and growth of two avocado cultivars.[J]. Scientia Horticulturae,2002,95:39-50
[22]郑殿升,王晓鸣,张京,等. 农作物种质资源技术规范丛书[M]. 北京:中国农业出版社,2006:90
[23]雷雄,游明鸿,白史且,等. 川西北高原50份燕麦种质农艺性状遗传多样性分析及综合评价[J]. 草业学报,2020,29(7):131-142
[24]武敬也,马琳,吴欣明,等. 180份饲用燕麦种质资源表型遗传多样性研究[J]. 草地学报,2023,31(5):1501-1510
[25]耿小丽,张榕,张少平,等. 130份燕麦种质表型性状多样性分析及评价[J]. 草业科学,2020,37(10):2022-2034
[26]周启龙,多吉顿珠,土登群培,等. 拉萨18个引进燕麦品种主要农艺性状和营养成分的遗传多样性分析[J]. 草业科学,2020,37(3):550-558
[27]胡佳燕,刘畅,李志坚,等. 不同类型皮燕麦在吉林省西部的产量与农艺性状的比较研究[J]. 草地学报,2021,29(10):2247-2257
[28]张坤,颜红波,周青平,等. 42份燕麦种质资源农艺性状多样性研究[J]. 青海大学学报(自然科学版),2014,32(4):11-16
[29]武俊英,刘景辉,王怀栋,等. 不同燕麦品种产量及其与构成因素的相关性研究[J]. 作物杂志,2011(5):36-40
[30]刘正理,程汝宏,李香月,等. 不同类型谷子品种叶片对产量形成效应的研究[J]. 中国农学通报,1995(3):12-16
[31]伏军,周清明,尹大德,等. 晚稻叶片性状及其在产量形成中的作用[J]. 湖南农学院学报,1993(3):211-216
[32]ARUNAH U L,CHIEZEY U F,ALIYU L,et al. Correlation and path analysis between sorghum yield to growth and yield characters[J]. Journal of Biology,Agriculture and Healthcare,2015,5(19):32-34
[33]孙建平,薛竹慧,杨国义,等. 施氮对晋北燕麦饲草主要农艺性状及干物质产量的影响[J]. 草地学报,2018,26(4):964-970
[34]李晶,南铭,刘彦明,等. 不同燕麦品种产量和品质及饲喂性能综合评价[J]. 草地学报,2023,31(4):1089-1098
[35]白晓雷,刘艳春,生国利,等. 35份皮燕麦种质遗传多样性的SSR和SRAP分析[J]. 内蒙古农业科技,2015,43(4):6-11
[36]王茜,李志坚,李晶,等. 不同類型燕麦农艺和饲草品质性状分析[J]. 草业学报,2019,28(12):149-158
[37]王桃. 高寒牧区36种燕麦营养生态特性及其生产效能评价[D]. 兰州:兰州大学,2010:5
[38]杨钰洁,梁国玲,刘文辉,等. 硅肥对青藏高原高寒地区燕麦抗倒伏性状及种子产量的影响[J]. 草业科学,2022,39(3):551-561
[39]BRULE V,RAFSANJANI A,PASINI D,et al. Hierarchies of plant stiffness[J]. Plant Science,2016,250(6):79-96
[40]KHOBRA R,SAREEN S,MEENA B K,et al. Exploring the traits for lodging tolerance in wheat genotypes:A review[J]. Physiology and Molecular Biology of Plants,2019,25(1):589-600
[41]SHEN Y S,XIANG Y,XU E S,et al. Major co-localized QTL for plant height,branch initiation height,stem diameter,and flowering time in an alien introgression derived Brassica napus DH population[J]. Frontiers in Plant Science,2018,9:390
[42]纪明雪,张智勇,齐冰洁,等. 燕麦种质资源抗倒伏及生物学性状的差异评价[J]. 麦类作物学报,2023,43(4):453-462
[43]梁国玲,刘文辉,秦燕,等. 不同燕麦资源生物量构成和茎秆特征与倒伏间的相关性研究[J]. 草地学报,2019,27(5):1339-1346
[44]ZHANG R,JIA Z,MA X,et al. Characterising the morphological characters and carbohydrate metabolism of oat culms and their association with lodging resistance[J]. Plant Biology,2020,22(2):267-276
[45]SILVEIRA D C,CAVERZAN A,EBONE L A,et al. Morphological traits of stem to indirect selection of resistance to lodging in Avena sativa L[J]. Journal of Crop Science and Biotechnology,2022,25(1):39-50
(责任编辑 闵芝智)
