赖草草地叶片养分、碳组分和防御性化合物对氮添加的响应
2024-02-02苏原何雨欣高阳阳梁雯君武帅楷郝杰刁华杰王常慧董宽虎
苏原 何雨欣 高阳阳 梁雯君 武帅楷 郝杰 刁华杰 王常慧 董宽虎
doi:10.11733/j.issn.1007-0435.2024.01.014
引用格式:
苏 原, 何雨欣, 高阳阳,等.赖草草地叶片养分、碳组分和防御性化合物对氮添加的响应[J].草地学报,2024,32(1):130-138
SU Yuan, HE Yu-xin, GAO Yang-yang,et al.Responses of the Nutrients,Carbon Components and Defensive Compounds in the Leaves of Leymus secalinus to Increasing Nitrogen Input[J].Acta Agrestia Sinica,2024,32(1):130-138
收稿日期:2023-07-12;修回日期:2023-08-24
基金项目:国家自然科学基金(32371670);山西省优秀博士来晋工作奖励资金科研项目(SXBYKY2022042);山西农业大学科技创新基金项目(2021BQ64);山西农业大学高层次人才专项资助(2021XG008);山西省重点实验室项目(202104010910017);大学生创新创业训练计划项目省级基金项目(20220164)资助
作者简介:
苏原(1989-),男,汉族,河南商丘人,讲师,主要从事草地生态学研究,E-mail:suyuan@sxau.edu.cn;*通信作者Author for correspondence,E-mail:changhui.wang@sxau.edu.cn;dongkuanhu@sxau.edu.cn
摘要:本研究依托山西右玉黄土高原赖草(Leymus secalinus)草地2017年建立的氮(Nitrogen,N)添加梯度試验平台(0~32 g·m-2·a-1),探究N添加对优势植物赖草叶片养分[N、磷(Phosphorus,P)、钾(Potassium,K)]、非结构性碳水化合物、结构性碳水化合物和防御性化合物含量的影响。结果表明:N素输入显著提高叶片N,K含量和N∶K和N∶P比值,降低叶片P含量和P∶K比值;N素输入显著降低叶片非结构性碳水化合物和结构性碳水化合物含量;N素输入显著增加叶片总酚、单宁和黄酮类化合物含量;PCA分析表明低N(≤8 g·m-2·a-1)和高N(>8 g·m-2·a-1)输入下赖草叶片性状存在显著差异,土壤无机N含量、土壤有效N∶P和N∶K是其主要影响因子。以上结果表明N素输入改变了盐渍化草地赖草叶片养分-碳组分-防御性化合物的分配策略。
关键词:氮添加梯度;叶片养分;叶片碳组分;防御性化合物;分配策略
中图分类号:S812.2 文献标识码:A 文章编号:1007-0435(2024)01-0130-09
Responses of the Nutrients,Carbon Components and Defensive Compounds
in the Leaves of Leymus secalinus to Increasing Nitrogen Input
SU Yuan1,2,3, HE Yu-xin1, GAO Yang-yang1,2,3, LIANG Wen-jun1,2,3, WU Shuai-kai1,2,3,
HAO Jie1,2,3, DIAO Hua-jie1,2,3, WANG Chang-hui1,2,3*, DONG Kuan-hu1,2,3*
(1.College of Grassland Science, Shanxi Agricultural University, Taigu, Shanxi Province 030801, China; 2. Shanxi Key Laboratory
of Grassland Ecological Protection and Native Grass Germplasm Innovation, Taigu, Shanxi Province 030801, China; 3. Youyu Loess
Plateau Grassland Ecosystem National Research Station, Shanxi Agricultural University, Youyu, Shanxi Province 037200, China)
Abstract:A field experiment was performed to explore the effects of N supplementation on nutrients (N,P,K),non-structural carbohydrate (NSC),structural carbohydrates (SC) and defensive compounds in leaves of dominant species Leymus secalinus in a salinized grassland of northern China,taking advantage of a field N addition gradients (0~32 g·m-2·a-1) experiment that has been established in the Loess Plateau in Youyu,Shanxi Province in 2017. The results showed that (1) Nitrogen input significantly increased the N and K contents,and N∶K and N∶P ratios,and decreased the P content and P∶K ratios in leaves. (2) Nitrogen input decreased the foliar NSC and SC content. (3) Nitrogen input significantly increased total phenols,tannins and flavonoids contents in leaves. (4) By PCA analysis,it showed that there was a trade-off relationship between leaf traits under low and high N treatments (> 8 g·m-2·a-1);the soil inorganic N,ratios of soil available N∶P and N∶K significantly affected most leaf traits of the investigated species. These results indicated that plants would invest more newly synthesized carbon into its defensive components and reduce the allocation to NSC and SC under the background of high N deposition,that is,N input altered the trade-off among plant traits.
Key words:Nitrogen addition gradient;Leaf nutrient;Leaf carbon component;Leaf defensive compound;Allocation strategy
由于化石燃料燃烧、土地利用改变、氮肥使用和畜禽养殖业的发展,中国的大气氮(Nitrogen,N)沉降由1980年的13.2 kg·ha-1·a-1升高到2010年的21.1 kg·ha-1·a-1,增加了60%[1]。虽然中国大气N沉降增加的速度在近几年有所减缓,但总的趋势仍在持续增加[1-2]。多项研究表明,N素是中国北方草地生态系统生产力的重要限制因子[3-4],因此外源N输入能够提高植物光合作用,促进植物生长,提高生产力。
氮、磷(Phosphorus,P)和钾(Potassium,K)在植物光合作用、遗传物质合成与运输和水分利用效率等方面有重要作用,叶片是植物进行光合作用合成有机物的重要场所,因此研究N素输入如何影响植物叶片N,P,K元素含量对理解植物生长和生态系统养分循环具有重要意义。多数研究已经证实了N输入显著增加植物叶片N含量[5-10],但N素输入对植物叶片P含量的影响存在较大差异[6,9-10,12-13]。与N,P相比,N素输入对草地植物K含量影响的研究较少,且结果差异很大[14-16]。
非结构性碳水化合物(Non-structural carbohydrates,NSC)在植物代謝、养分运输、渗透调节和土壤有机质形成过程中至关重要[17-18]。NSC主要由可溶性糖和淀粉组成,当受到环境胁迫时,两者可以相互转化[19-21]。在内蒙古典型草原的一项研究发现N素输入显著降低了羊草叶片可溶性糖和淀粉含量[7],而在相同地点一个50天的13C标记实验结果表明N素输入对叶片可溶性糖、淀粉和NSC含量的影响因测定时间而异[20]。这些研究表明N添加对植物可溶性糖、淀粉和NSC的影响与实验时间、物种以及N添加量有关[23],有待进一步深入研究。此外,植物结构性碳水化合物(Structural carbohydrates,SC)和次生代谢物(如单宁、总酚和黄酮类化合物)对凋落物分解和天敌防御有显著影响[24]。植物生长分化平衡假设认为,氮素输入下植物积累超过生长需求的初级代谢产物可以廉价地转化为次级代谢产物[20]。最近研究发现氮添加对东北红豆杉(Taxus cuspidata)幼苗叶片黄酮类化合物含量有显著影响,随N素输入量增加(0~12 g·m-2·a-1),黄酮类化合物含量表现出先上升后下降的趋势[27]。而另一研究发现N素输入(0~9 g·m-2·a-1)对油松(Pinus tabuliformis)和蒙古栎(Quercus mongolica)叶片总酚含量的影响与采样时间和N输入量有关[28],但N素输入一致降低了油松和蒙古栎叶片单宁含量。尽管以上研究提高了有关N素输入对植物防御性化合物影响的认识,但N素输入如何影响植物叶片养分-碳组分-防御性化合物之间的分配策略,尚不清楚。
农牧交错带草地是我国北方半湿润农区与干旱、半干旱牧区接壤的过渡地带,是生态脆弱区和响应环境变化的敏感区[29-30]。本研究利用山西右玉黄土高原草地生态系统研究站内N添加梯度(0,1,2,4,8,16,24,32 g·m-2·a-1)实验平台,采集优势植物赖草(Leymus secalinus)的叶片,对植物叶片养分(N,P,K)、SC(半纤维素、纤维素、木质素)、NSC(淀粉、可溶性糖)和防御性碳水化合物(总酚、单宁、黄酮类化学物)含量进行分析,拟回答以下几个科学问题:1) N素输入对叶片N,P,K及其化学计量有何影响?2) N素输入对叶片NSC、SC和防御性化合物有何影响?3) N素输入对叶片养分-碳组分-防御性化合物的分配策略有何影响?其主要影响因素是什么?
1 材料与方法
1.1 试验地概况
本研究在山西右玉黄土高原草地生态系统国家定位观测研究站(112.29°E,39.28°N)开展。试验区海拔1 348 m,近30年平均气温为4.6℃,降水量为425 mm,主要集中在生长季(5—9月)。土壤(0~10 cm)有机碳、总N和总P含量分别为5.43,0.80,0.39 mg·g-1、土壤pH值9~10,盐分浓度1.44~3.99 mg·g-1[31]。群落优势种是赖草,占群落总地上生物量70%~90%。
1.2 试验设计
试验平台于2017年4月建立,采用完全随机区组设计,共设置8个N添加水平(0,1,2,4,8,16,24和32 g·m-2·a-1),分别用N0,N1,N2,N4,N8,N16,N24和N32表示,每个处理6个重复,小区面积54 m2 (6 m×9 m),共48个小区,相邻小区间隔2 m。添加的氮为硝酸铵(NH4NO3),每年的N添加量平均分为5次,分别于每年的5—9月的月初添加[32-33]。每次将预先称好的各处理对应的NH4NO3溶于10 L水中(小于0.2 mm降水量),使用肩背充电式喷水器均匀喷洒,对照处理喷洒等量的水。
1.3 植物和土壤样品采集与测定
2022年8月进行野外调查和植物、土壤样品的采集。土壤样品:在每个样地内按照S形随机选取3个样点采集0~10 cm的土壤样品,然后混合成一个土壤样品。植物样品:采集完整和健康的赖草叶片50片,样品经105℃杀青处理30 min后,再置于65℃烘箱中烘干至恒量。
植物N,P和K的测定:全N含量采用元素分析仪(Vario MACRO cube,Elementar,Analysensysteme GmbH)测定;全P含量采用H2SO4-H2O2进行消煮,钼蓝比色法测定;全K含量用电感耦合等离子体发射光谱仪(ICP)测定[16]。
可溶性糖和淀粉含量的测定:采用蒽酮比色法测定可溶性糖和淀粉含量[34],利用紫外分光光度计在620 nm处的最大吸收值对其进行测定并计算。
纤维素、半纤维素及木质素含量测定:称取 300 mg样品于试管中,加入3.00 mL浓硫酸(72%),搅拌至其充分水解,置于马弗炉中575℃烘至恒重。再置于马弗炉中烘干,干燥器中称重,两次称重质量的差值即为酸不溶木质素含量。另取滤液20 mL,调节pH值至5~6,过0.2 μm滤膜,注入高效液相色谱的自动进样器测定纤维素及半纤维素含量。
比色法测定总酚和简单酚含量,两者相减获得单宁含量[35]。黄酮类化合物含量通过NaNO2-Al(NO)3-NaOH显色法测得[20]。
土壤无机氮(NH+4和NO-3)含量用全自动流动分析仪(Continuous-Flow Analysis-CFA,SEAL,Nordestedt,Germany)测定,土壤有效磷含量测定采用NaHCO3浸提钼锑抗比色法,有效钾含量测定用乙酸胺浸提火焰光度法[16]。
1.4 数据处理
数据分析之前,首先对数据进行正态性检验。具体分析方法如下:(1)回归分析研究N添加对土壤速效养分含量及化学计量比、叶片N,P,K含量及其化学计量比以及SC,NSC和防御性化合物含量的影响;(2)利用Pearson相关分析研究叶性状之间的两两关系,再利用主成分分析(PCA)探究叶片养分、碳组分与防御性化合物的权衡关系;(3)Pearson相关分析研究土壤速效养分与叶片养分、碳组分和防御性化合物之间的关系。最后,用R4.02和Origin 9.0进行绘图。
2 结果与分析
2.1 N添加对土壤速效养分及其化学计量比的影响
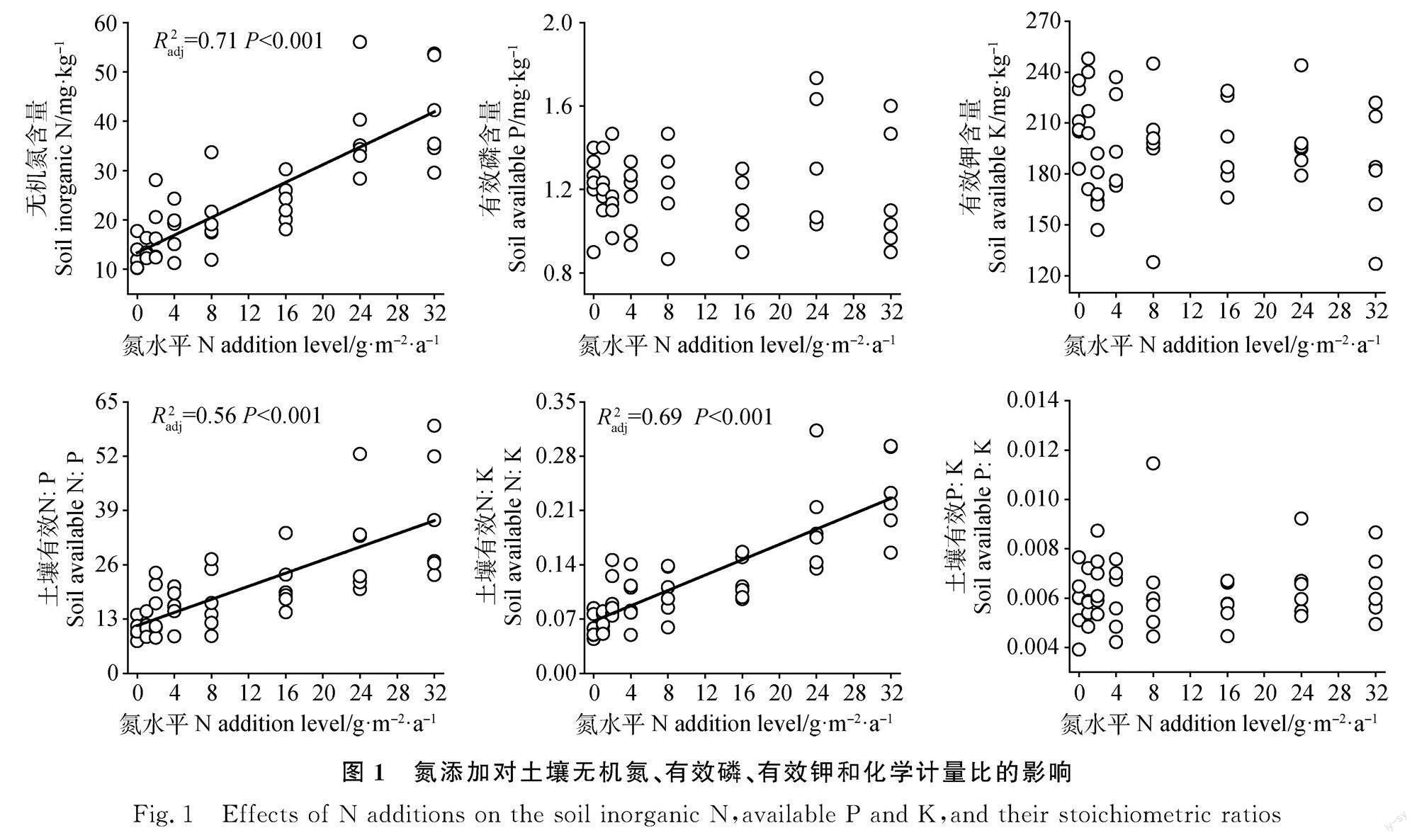
N添加显著提高了土壤无机N含量、有效N∶P和有效N∶K(图1,P<0.01),而N添加对土壤速效P、速效K和有效P∶K没有显著影响(图1)。
2.2 N添加对叶片N,P,K及其化学计量比的影响
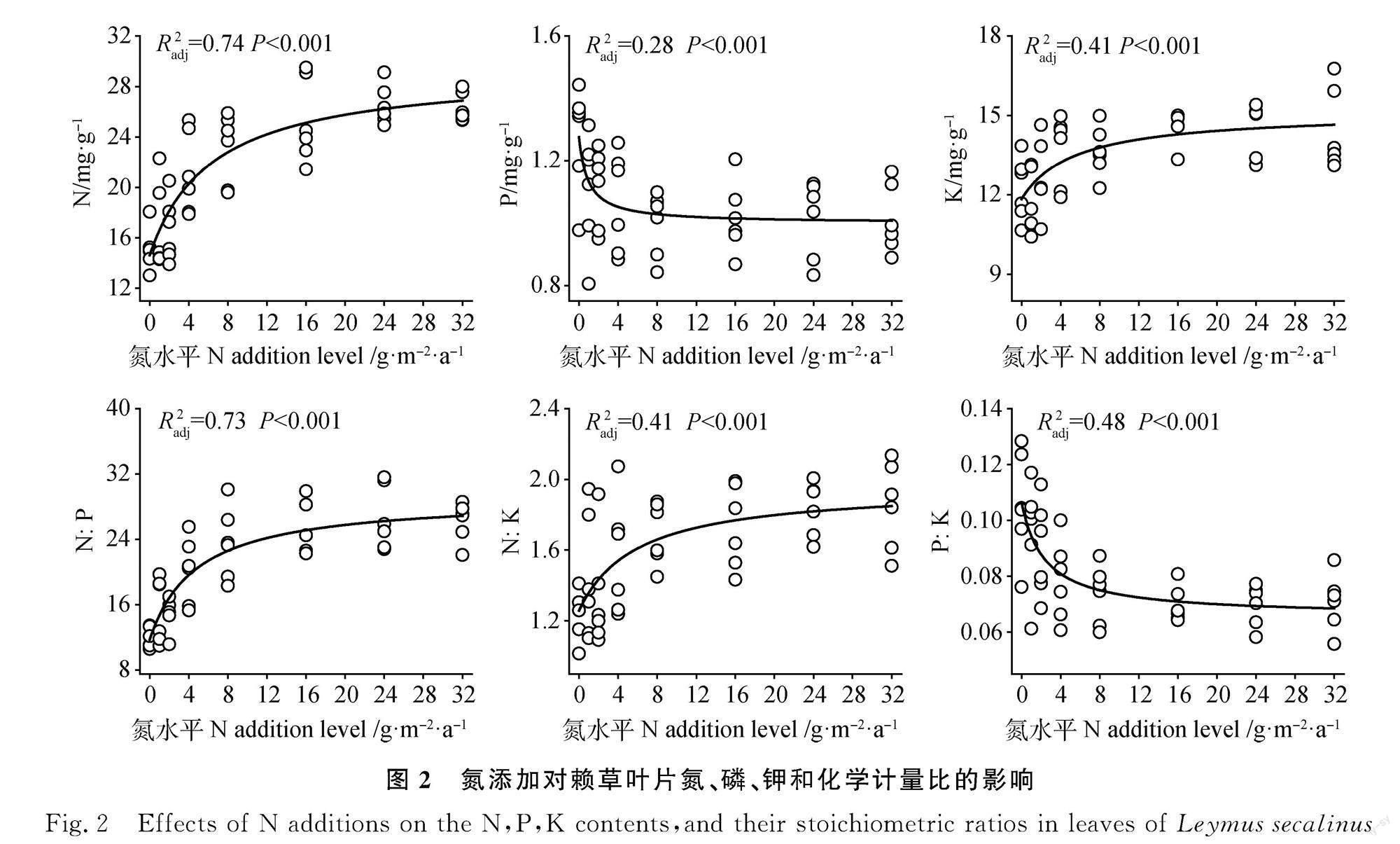
随N素添加量的增加,叶片N和K含量呈现非线性增加趋势,而叶片P含量呈现非线性降低趋势(图2,P<0.01)。此外,N添加显著提高了叶片N∶P和N∶K,显著降低了P∶K(图2,P<0.01)。
2.3 N添加對叶片SC和NSC的影响
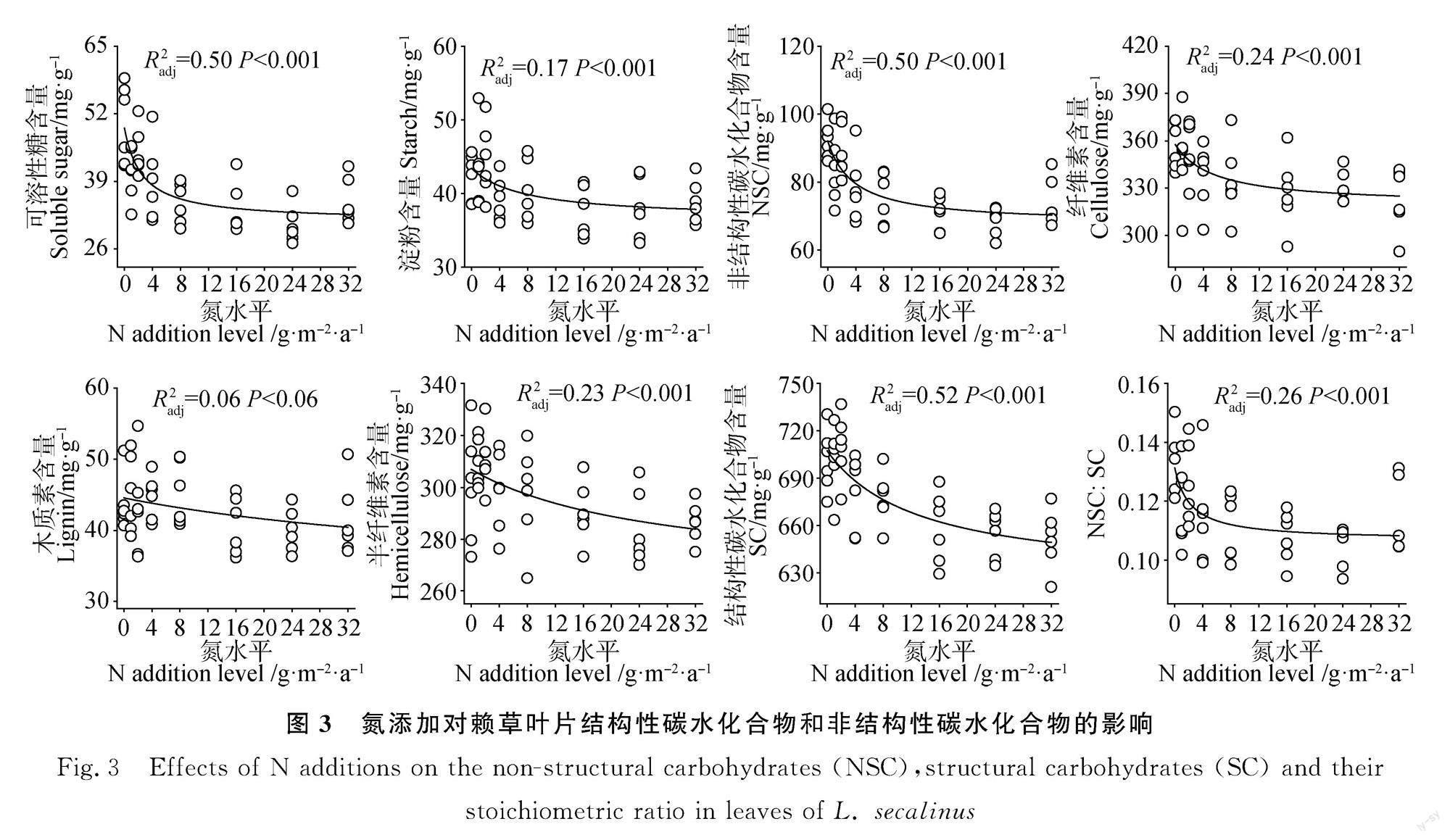
N添加显著影响了叶片NSC和SC含量(图3,P<0.01)。随N素添加量的增加,叶片可溶性糖、淀粉、NSC、木质素、纤维素、半纤维素、SC含量和NSC∶SC均呈现非线性降低趋势(图3,P<0.01)。N添加有降低叶片木质素含量的趋势。
2.4 N添加对叶片防御性化合物的影响
N添加显著影响了叶片防御性化合物含量(图4,P<0.01)。随N素输入量的增加,叶片总酚、单宁和黄酮类化合物含量呈现非线性增加趋势。
2.5 N添加下叶片养分、碳组分和防御性化合物的权衡关系
相关分析结果表明大多数叶片性状间有显著相关性(木质素除外)(图5)。例如,叶片N含量与可溶性糖、淀粉和NSC含量显著负相关,而与总酚、单宁和黄酮类化合物含量显著正相关;叶片P含量与可溶性糖、淀粉和NSC含量显著正相关,而与总酚、单宁和黄酮类化合物含量显著负相关(图5)。
PCA分析结果表明PC1和PC2的解释率分别为83.27%和6.75%,累计为90.02%(图6)。其中叶片有着更高含量的N,K和防御性化合物,会有更低含量的P,NSC和SC。此外,低N处理(≤ 8 g·m-2·a-1)和高N处理(>8 g·m-2·a-1)叶片性状存在显著差异。
2.6 土壤速效养分对叶片养分、碳组分和防御性化合物的影响
相关分析结果表明土壤无机氮和叶片所有性状有显著相关性(图7),而土壤有效P只与叶片P含量显著正相关。土壤有效K和土壤N∶K与叶片性状无显著相关性,而土壤有效N∶P和N∶K与大多数叶片性状有显著相关性(木质素除外)。
3 讨论
3.1 N添加对植物养分和化学计量比的影响
本研究发现N添加显著提高了优势植物赖草叶片N含量,而降低了叶片P含量,导致叶片N∶P显著增加,主要是由于N素输入显著提高了土壤无机N含量,对土壤速效P含量没有造成显著影响。已有的很多研究也报道了N素输入显著提高了局域尺度[6,9-10,13]、区域尺度[11]和全球尺度植物叶片N含量[36-37],而N素输入对植物P含量的影响无一致结论[6,9-10,13,38]。这些差异产生的原因可能是:(1)N素输入显著提高了土壤磷酸酶活性,加速了土壤P素周转,提高了土壤速效P含量,促进了植物P素吸收[39-40];(2)北方草地生态系统普遍受N限制,N素输入显著提高了植物生产力,生物量增加导致的稀释效应会降低植物P浓度。本研究中N素输入显著提高了赖草地上部生物量,而N素输入对土壤有效P含量无显著影响,这可能导致本研究中植物叶片P浓度降低。
此外,本研究发现N素输入显著提高了叶片K浓度。已有研究发现N素输入对植物K含量的影响变化很大。例如,N素输入增加草甸草原植物地上部K含量[5],而对典型草原和高寒草原植物K含量无显著影响[14-15]、随氮素输入量的增加,草甸草原植物K呈先增加后降低的趋势[16]。本研究中N素输入降低了表层土壤含水量[31],植物可能会增加叶片K含量来适应降低的土壤含水量[41]。在全球变化研究中K元素长期被忽视,未来更多研究应该关注K元素的生态学过程[41]。
3.2 N添加对植物碳组分和防御性化合物的影响
本研究发现N素输入显著降低了叶片SC和NSC,并显著降低了叶片NSC∶SC,这表明N素输入改变了叶片碳组分,植物将更多新合成的有机物向SC分配。有研究发现N添加(0~50 g·m-2·a-1)显著降低典型草原优势植物木质素、纤维素和半纤维素含量[25,42];而另一項研究发现N添加(0~32 g·m-2·a-1)显著增加高寒草原优势植物紫花针茅木质素含量,而对薹草和多裂萎陵菜木质素含量无显著影响[26]。这些差异可能和生态系统类型、N添加量、N形态、物种和功能群有关。
本研究发现N素输入显著降低了植物叶片可溶性糖、淀粉和NSC含量,这与N添加增加或没有显著影响半干旱草地优势植物NSC的结果不同[21-22]。全球尺度整合分析结果表明N添加增加草本植物叶片可溶性糖含量(+4.3%,n=84),降低了淀粉含量(-28.4%,n=58),但对茎秆可溶性糖(n=25)和淀粉(n=2)含量没有显著影响[23]。以上结果说明N素输入对植物NSC的影响因物种和器官而异。鉴于当前有关N素输入对草本植物NSC的研究不足,建议加强关于N素输入对植物不同器官、功能群和群落水平的NSC影响的研究。
此外,本研究发现N输入显著增加了植物总酚、单宁和黄酮类化合物含量。该结果可能有如下原因:(1)N素输入显著增加了植物叶N含量和N∶P比值,降低了木质素∶N比值,进而显著提高了植食性昆虫的数量[43],植物为了防御天敌提高了对防御性化合物的合成[17];(2)N输入增加了半干旱草地植物病原真菌感染的机会[44],进而提高防御性化合物的合成;(3)植物生长分化平衡假设认为,N素输入下植物积累超过生长需求的初级代谢产物可以廉价地转化为次级代谢产物,可以有效避免资源浪费[7,45]。目前,有关N输入对草原植物防御性化合物影响的报道还不多见,未来应该关注N素输入对草地植物次生代谢物与病虫害关系的研究。
3.3 N添加对植物养分-碳组分-防御性化合物分配策略的影响
本研究结果表明大多数叶片性状间存在显著相关性(木质素除外),如叶片N与防御性化合物呈显著正相关,与SC和NSC呈显著负相关,这说明叶片性状间存在协同和权衡关系。有研究发现N素输入同时降低了半干旱草原羊草叶片NSC和防御性化合物含量[7]。而另一项对幼苗叶经济谱和防御性状的研究则发现叶N含量与植物单宁、总酚和类黄酮含量呈显著负相关[35]。本研究中N素输入显著提高了叶片N含量,降低了叶片中SC,进而显著提高了牧草品质[42]。改善的牧草品质增加了植食性昆虫采食和病原真菌感染的机会[43-44],植物为了抵御草食动物和受病原菌的伤害会产生更多的防御性化合物,进而提高了叶中防御性化合物含量[17,46]。此外,赖草叶总酚与单宁和黄酮类化合物含量呈显著正相关关系,说明赖草叶片防御性状间存在协同作用,进而形成多种防御化合物协同作用的防御机制,这和先前研究结果一致[35]。
主成分分析结果表明赖草叶片性状对低N(≤ 8 g·m-2·a-1)和高N添加(> 8 g·m-2·a-1)的响应具有明显差异,表现为随N素输入量增加,叶片P,NSC和SC含量降低,而叶片N,K和防御性化合物含量增加。本研究结果表明赖草叶片性状间存在权衡关系,N素输入改变了叶片性状间的权衡关系。即在高N输入时,植物会提高自身基质质量(如高N含量、低木质素∶N),同时叶片会选择将更多资源分配给防御性化合物,以抵御草食动物或者病原菌等的威胁[43-44],但也有研究发现植物功能性状与防御性状间不存在权衡关系[47]。植物功能性状与防御性状的权衡对于维持植物生产力、群落稳定性和生态系统服务功能至关重要[48]。全球变化背景下,植物功能性状和防御性状的关系亟待进一步研究,特别是在研究较少的草本植物主导的生态系统。
4 结论
本研究通过模拟不同水平大气N沉降,探究了盐渍化草地优势植物叶片养分-碳组分-防御性化合物对N添加的响应。结果表明N素输入显著增加赖草叶片N和K含量,降低P含量,改变了化学计量比;降低赖草叶片SC和NSC含量,提高防御性化合物含量。以上结果表明N素输入改变了叶片养分-碳组分-防御性化合物的分配策略。土壤无机氮和速效养分比(无机氮∶有效磷和无机氮∶有效钾)是赖草叶片养分-碳组分-防御性化合物分配策略的重要影响因素。
参考文献
[1]LIU X J,ZHANG Y,HAN W X,et al. Enhanced nitrogen deposition over China[J]. Nature,2013,494:459-462
[2]YU G R,JIA Y L,PIAO S L,et al. Stabilization of atmospheric nitrogen deposition in China over the past decade[J]. Nature Geoscience,2019,12:424-429
[3]BAI Y F,WU J G,CIARK C M,et al. Tradeoffs and thresholds in the effects of nitrogen addition on biodiversity and ecosystem functioning:Evidence from Inner Mongolia grasslands[J]. Global Change Biology,2010,16:358-372
[4]PENG Y F,CHEN H Y H,YANG Y. Global pattern and drivers of nitrogen saturation threshold of grassland productivity[J]. Functional Ecology,2020,34:1979-1990
[5]高貝,胡艳宇,张志委,等. 氮素添加对呼伦贝尔草甸草原植物氮钾元素含量和计量比的影响[J]. 应用生态学报,2022,33:981-987
[6]LU X T,REED S,YU Q,et al. Convergent responses of nitrogen and phosphorus resorption to nitrogen inputs in a semiarid grassland[J]. Global Change Biology,2013,19:2775-2784
[7]GONG J R,ZHANG Z H,WANG B,et al. N addition rebalances the carbon and nitrogen metabolisms of Leymus chinensis through leaf N investment[J]. Plant Physiology and Biochemistry,2022,185:221-232
[8]HU Y,GUO A X,LI X Y,et al. Multi-trait functional diversity predicts ecosystem multifunctionality under nitrogen addition in a desert steppe[EB/OL]. https://doi.org/10.1007/s11104-022-05731-8,2022-10-05/2023-07-12
[9]LI L,GAO X P,LI X Y,et al. Nitrogen (N) and phosphorus (P) resorption of two dominant alpine perennial grass species in response to contrasting N and P availability[J]. Environmental and Experimental Botany,2016,127:37-44
[10]SU Y,MA X F,LE J J,et al. Decoupling of nitrogen and phosphorus in dominant grass species in response to long-term nitrogen addition in an alpine grassland in Central Asia[J]. Plant Ecology,2021,222(9):261-274
[11]SU Y,MA X F,GONG Y M,et al. Responses and drivers of leaf nutrients and resorption to nitrogen enrichment across northern Chinas grasslands:A meta-analysis[J]. Catena,2021,199:105110
[12]HOU S L,YANG J J,YIN J X,et al. Changes of community composition strengthen the positive effects of nitrogen deposition on litter N∶P stoichiometry in a semi-arid grassland[J]. Plant and Soil,2020,473:63-71
[13]PENG Y F,LI F,ZHOU G Y,et al. Linkages of plant stoichiometry to ecosystem production and carbon fluxes with increasing nitrogen inputs in an alpine steppe[J]. Global Change Biology,2017,23:5249-5259
[14]ZHANG D Y,PENG Y F,LI F,et al. Above- and below-ground resource acquisition strategies determine plant species responses to nitrogen enrichment[J]. Annals of Botany,2021,128:31-44
[15]ZHANG J H,REN T T,YANG J J,et al. Leaf multi-element network reveals the change of species dominance under nitrogen deposition[J]. Frontiers in Plant Science,2021,12:580340
[16]FENG X,WANG R Z,YU Q,et al. Decoupling of plant and soil metal nutrients as affected by nitrogen addition in a meadow steppe[J]. Plant and Soil,2019,443:337-351
[17]HUANG J,WANG X M,ZHENG M H,et al. 13-year nitrogen addition increases nonstructural carbon pools in subtropical forest trees in southern China[J]. Forest Ecology and Management,2021,481:118748
[18]HERRER-RAMIRE D,SIERRA C A,ROMERMAN C,et al. Starch and lipid storage strategies in tropical trees relate to growth and mortality[J]. New Phytologist,2021,230:139-154
[19]LI W B,ZHANG H X,HUANG G Z,et al. Effects of nitrogen enrichment on tree carbon allocation:A global synthesis[J]. Global Ecology and Biogeography,2019,29:573-589
[20]WANG B,GONG J R,ZHANG Z H,et al. Nitrogen addition alters photosynthetic carbon fixation,allocation of photoassimilates,and carbon partitioning of Leymus chinensis in a temperate grassland of Inner Mongolia[J]. Agricultural and Forest Meteorology,2019,279:107743
[21]郭建斌,徐紅伟,薛萐. 黄土丘陵区撂荒草地不同生态位物种非结构性碳水化合物对氮添加的响应[J]. 水土保持学报,2022,36(3):153-158
[22]王雪,雒文涛,庾强,等. 半干旱典型草原养分添加对优势物种叶片氮磷及非结构性碳水化合物含量的影响[J]. 生态学杂志,2014,33(7):1795-1802
[23]DU Y,LU R L,XIA J Y. Impacts of global environmental change drivers on non-structural carbohydrates in terrestrial plants[J]. Functional Ecology,2022,34:1525-1536
[24]CHOME M,GUITTONNY-LARCHEVEQU M,FERNANDEZ C,et al. Plant secondary metabolites:A key driver of litter decomposition and soil nutrient cycling[J]. Journal of Ecology,2016,104:1527-1541
[25]HOU S L,HATTENSCHWILE S,YANG J J,et al. Increasing rates of long-term nitrogen deposition consistently increased litter decomposition in a semi-arid grassland[J]. New Phytologist,2021,229:296-307
[26]WEI B,ZHANG D Y,KOU D,et al. Decreased ultraviolet radiation and decomposer biodiversity inhibit litter decomposition under continuous nitrogen inputs[J]. Functional Ecology,2022,36:998-1009
[27]孟秋实,秦倩倩,刘艳红. 氮添加对东北红豆杉幼苗生长发育及生理特征的影响[J]. 生态学杂志,2022,41:2325-2334
[28]陈克利. 氮添加对油松和辽东栎幼苗次生代谢过程的影响[D]. 杨凌:西北农林科技大学,2019:21-34
[29]高树琴,段瑞,王竑晟,等. 北方农牧交错带在保障国家大粮食安全中发挥重要作用[J]. 中国科学院院刊,2021,36(6):643-651
[30]刘平,刘学军,骆晓声,等. 山西北部农村区域大气活性氮沉降特征[J]. 生态学报,2016,36(17):5353-5359
[31]CHEN X P,DIAO H J,WANG S P,et al. Plant community mediated methane uptake in response to increasing nitrogen addition level in a saline-alkaline grassland by rhizospheric effects[J]. Geoderma,2023,429,116235
[32]赵晓洁,张雄伟,薛江博,等. 短期氮添加对山西右玉黄土高原盐渍化草地氨挥发的影响[J]. 草地学报,2022,30(4):992-999
[33]郑慧,薛江博,郝杰,等. 短期不同水平氮添加对华北盐渍化草地土壤磷组分的影响[J]. 草地学报,2022,30(3):712-720
[34]董涵君,王兴昌,苑丹阳,等. 温带不同材性树种树干非结构性碳水化合物的径向分配差异[J]. 植物生态学报,2022,46:722-734
[35]程思祺,姜峰,金光泽. 温带森林阔叶植物幼苗叶经济谱及其与防御性状的关系[J]. 植物生态学报,2022,46:678-686
[36]XIA J Y,WAN S Q. Global response patterns of terrestrial plant species to nitrogen addition[J]. New Phytologist,2008,179:428-439
[37]YOU C M,WU F Z,YANG W Q,et al. Nutrient-limited conditions determine the responses of foliar nitrogen and phosphorus stoichiometry to nitrogen addition:A global meta-analysis[J]. Environmental Pollution,2018,241:740-749
[38]LU X T,HAN X G. Nutrient resorption responses to water and nitrogen amendment in semi-arid grassland of Inner Mongolia,China[J]. Plant and Soil,2009,327:481-491
[39]JIAN S Y,LI J W,CHEN J,et al. Soil extracellular enzyme activities,soil carbon and nitrogen storage under nitrogen fertilization:A meta-analysis[J]. Soil Biology and Biochemistry,2016,101:32-43
[40]WANG R Z,YANG J J,LIU H Y,et al. Nitrogen enrichment buffers phosphorus limitation by mobilizing mineral-bound soil phosphorus in grasslands[J]. Ecology,2022,103,e3616
[41]SARDAN J,PENUEIA J. Potassium:a neglected nutrient in global change[J]. Global Ecology and Biogeography,2015,24:261-275
[42]HOU S L,FRESCHE G T,YANG J J,et al. Quantifying the indirect effects of nitrogen deposition on grassland litter chemical traits[J]. Biogeochemistry,2018,139:261-273
[43]GU H J,WANG H,LIU M,et al. Leaf N:P stoichiometry overrides the effect of individual nutrient content on insect herbivore population dynamics in a Tibetan alpine grassland[J]. Agriculture,Ecosystems and Environment,2022,336:108032
[44]LIU X,LU Y W,ZHANG Z H,et al. Foliar fungal diseases respond differently to nitrogen and phosphorus additions in Tibetan alpine meadows[J]. Ecological Research,2019,35:162-169
[45]NEILSO E H,GOODGER J Q,WOODRO I E,et al. Plant chemical defense:At what cost?[J]. Trends in Plant Science,2013,18:250-258
[46]BARBEHEN R V,PETER C C. Tannins in plant-herbivore interactions[J]. Phytochemistry,2011,72:1551-1565
[47]CHAUVI K M,ASNER G P,MARTIN R E,et al. Decoupled dimensions of leaf economic and anti-herbivore defense strategies in a tropical canopy tree community[J]. Oecologia,2018,186:765-782
[48]AGRAWAL A. A scale-dependent framework for trade-offs,syndromes,and specialization in organismal biology[J]. Ecology,2020,101:e02924
(責任编辑 闵芝智)
