莲花洲港不同涵养区域底栖动物群落短期变化研究
2024-02-01叶丽娟刘绍平段辛斌陈大庆郑永华
叶丽娟,王 珂,刘绍平,段辛斌,陈大庆,郑永华
(1.中国水产科学研究院长江水产研究所,国家农业科学重庆观测实验站,武汉 430223;2.西南大学水产学院,重庆 400715)
长江干线武汉至安庆段6 m水深航道整治工程的实施发挥了长江黄金水道的价值,提高了长江航运整体通过能力,但不可避免地对附近河道水生生境造成了一定的影响。杨雪[1]、叶属峰等[2]研究表明,航道工程的实施造成了底栖动物、浮游生物密度和生物量的下降。为了平衡开发与保护,武汉至安庆段航道整治工程围绕“生态优先”的整治原则,以生境修复和生态涵养为目的,设计实施了一系列生态建设工程。其中,在东流水道莲花洲港建设了生态涵养区,铺设透水框架和鱼巢排等结构,以营造水生生物庇护场所,改善水生生物栖息环境。
涵养工程的实施促进了江段底沙沉积[3,4]。随着底沙沉积,有机质逐渐富集[5],水生植物、底栖动物等也得到一定恢复。底栖动物是食物链的重要一环。由于各类底栖动物对环境条件的适应性以及对污染程度的耐受力和敏感程度不同,故其种群结构、优势种类、数量等参数可用来反映水环境质量状况[6]。随着生态涵养区实施年份的增加,其涵养效果如何,水环境是否有改善,均可通过底栖动物作为切入点进行研究。
目前,对底栖动物演替的研究已经比较成熟[7-10],但是对长江涵养工程区域底栖动物的演替并未涉及。本研究通过对长江下游莲花洲港生态涵养区大型底栖动物的多年调查,揭示底栖动物群落结构的短期变化,并且结合不同构筑物类型,了解不同涵养区域底栖动物群落变化的差异,以期为促进生态涵养区在其他水域推广和应用提供数据支撑,推动长江航道绿色发展。
1 材料与方法
1.1 研究区域
莲花洲港生态涵养区位于武汉至安庆段的东流水道,下距安庆市41 km,是武汉至安庆段6 m水深整治工程重点项目之一,该区域整体禁航,并设置了两处生境改良区。其中,第一处生境改良区为鱼巢排区,该区域放置2 m(长)×2 m(宽)×0.5 m(厚)田字形钢混透空格栅,横向放置80件、纵向放置25件,各构件相互间隔1 m,形成阵列。第二处生境改良区为透水框架区,该区域放置为“W型”的透水框架带,透水框架带与水流方向成30°夹角,长240 m,宽10 m,每平方米三层6架。
1.2 采样点布设
为探究生态涵养区底栖动物群落的变化,于2019年10月、2020年11月、2021年的10月对东流水道莲花洲港生态涵养区进行了采样调查,莲花洲生态涵养区建设完工于2019年6月,6、7月份为丰水期,采样难度大,采样结果不准确,不能很好地解释底栖动物的变化情况,因此选取了10月份、11月份(平水期)为采样时间点;为了解不同涵养区底栖动物变化的差异,本研究在生态涵养区设置5个断面,分别位于洲头、洲中、洲尾、透水框架区和鱼巢排区,共15个采样点(见表1、图1)。

图1 莲花洲港采样点图Fig.1 Sampling sites for macrozoobenthos in Lianhua Island port

表1 莲花洲港采样点坐标Tab.1 Coordinates of sampling points in Lianhua Island port
1.3 调查方法
1.3.1 水体理化指标的测定
水温(WT)、溶解氧(DO)、pH采用YSI水质分析仪(YSI Pro 2000,美国)测定,流速(FV)、透明度(SD)、水深(WD)分别使用旋杯式流速仪[11]、萨氏盘(Secchi)、采泥器绳和卷尺测定。总氮(TN)、总磷(TP)、氨氮(NH3-N)、硝态氮(NO3-N)和化学需氧量(CODCr)等环境因子现场采集水样后带回实验室,通过Hach DR1900可见分光光度计进行测定。
1.3.2 底栖动物的采集与鉴定
使用采样面积为(1/16)m2的Petersen底样采集器进行底栖动物的采样,每个采样点采集2~3次。采集的底泥使用40目铜筛网进行筛洗,筛洗后的底泥与底栖动物混合物装入标记后的样品瓶,之后带回实验室进行分拣,分拣出的底栖动物用75%的酒精固定。使用OLYMPUS SZ61解剖镜和OLYPUS BX51显微镜进行底栖动物的鉴定,参考文献[12,13]将底栖动物鉴定至尽可能低的分类单元,然后计数和称重。计数时,若样本损坏,只需记录头部。称量湿重时,先用滤纸吸干样本表面的水分[14]。
1.4 数据分析
底栖动物群落特征评价采用以下指数:Margalef丰富度指数(D)、ShannonWiener多样性指数(H′)、Pielou均匀度指数(J)、优势度(Y),若Y≥0.02,表明该底栖动物为优势种[15]。
使用Excel 2016进行数据的整理和基础计算。运用SPSS 22.0进行单因素方差分析(One-way ANOVA)和双因素方差分析(Two-way ANOVA)。
2 结果
2.1 环境参数的变化
单因素方差表明(表2),环境参数DO、TN、FV在不同时间、不同区域呈现显著性差异。WT、DO、TN、pH、FV、SD均呈现较显著的年际差异。

表2 2019-2021年调查区域环境参数变化Tab.2 The changes of regional environmental parameters were investigated from 2019 to 2021
其中,不同年份透水框架区流速均小于鱼巢排区,鱼巢排区、透水框架区FV呈现逐年降低的趋势,透水框架区透明度在2020年11月、2021年10月均大于鱼巢排区。FV、SD可能是影响不同区域、不同时间底栖动物分布差异的主要环境因素。
2.2 物种组成
2019-2021年共采集到大型底栖动物32种,隶属于6纲(昆虫纲、甲壳纲、双壳纲、腹足纲、寡毛纲、多毛纲)3门(环节动物门、节肢动物门、软体动物门)。不同时间、不同区域底栖动物状况见表3,结果显示,总体上底栖动物物种数呈现逐年上升的趋势。

表3 不同区域、点位数底栖动物物种数变化情况Tab.3 Sampling point diagram of Lianhua Island port
底栖动物优势种及优势度如表4所示,从总体上看,2021年甲壳纲的日本大螯蜚(Grandidierellajaponica)为绝对优势种,且优势度相对于2019年显著增大。鱼巢排区优势种由2019年甲壳纲的日本大螯蜚转变为2021年甲壳纲的日本大螯蜚、昆虫纲的马速达多足摇蚊(Polypedilummasudai)、多毛纲的围沙蚕一种(Perinereissp);透水框架区底栖动物优势种种类呈现逐年增加的趋势,由2019年的1种,增加至2020年的3种,2021年的4种;其他区域底栖动物优势种种类数目在2020年有所下降,到2021年显著增加。

表4 不同区域底栖动物优势种的优势度Tab.4 The dominance of dominant benthic species in different regions
2.3 密度及生物量的变化
2019-2021年调查结果显示(见图2、3),总体上,2019年10月底栖动物平均密度为13.51 ind/m2,平均生物量为0.04 g/m2;2020年11月底栖动物平均密度为26.31 ind/m2,平均生物量为2.07 g/m2;2021年10月底栖动物平均密度为88.53 ind/m2,平均生物量为2.96 g/m2,大型底栖动物平均密度、平均生物量均呈现逐年上升的趋势。

图2 底栖动物不同年份密度变化Fig.2 Changes of benthic density in different year

图3 底栖动物不同年份生物量变化Fig.3 Changes of benthic biomass in different year
从不同研究区域来看,鱼巢排区、透水框架区底栖动物密度和生物量均呈现逐年上升的趋势。鱼巢排区底栖动物生物量2020年出现较大幅度增长,2021年有所下降,但是高于2019年底栖动物生物量水平。其他区域大型底栖动物平均生物量呈现逐年上升的趋势,底栖动物平均密度2020年有所降低,2021年增加至21.33 ind/m2,且高于2019年和2020年。
2.4 多样性变化
调查结果显示(见图4、5、6),总体上,2019年10月、2020年11月、2021年10月底栖动物D、H′、J均呈现逐年上升的趋势。从不同区域来看,鱼巢排区、其他区域底栖动物D、H′、J呈现逐年上升的趋势;透水框架区丰富度指数由2019年的0.74增加至2020年的1.30、2021年的1.14,H′、J呈现逐年上升的趋势。
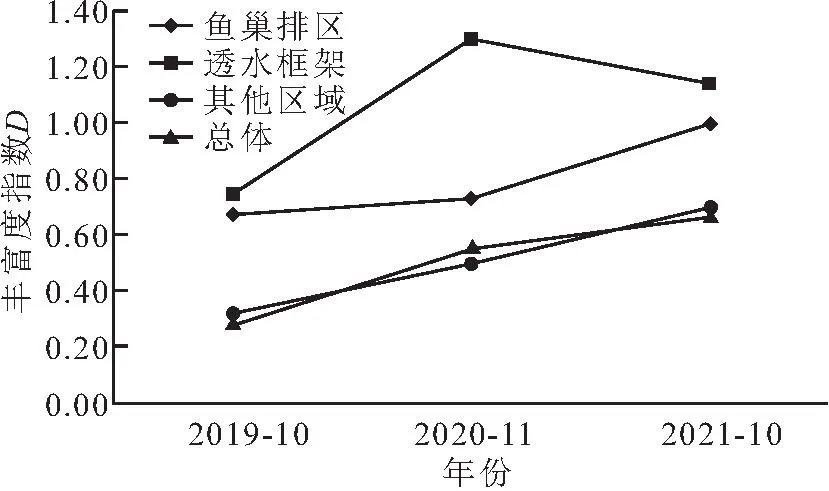
图4 不同区域底栖动物丰富度(D)指数变化Fig.4 Changes in the richness(D)index of benthic animals in different regions
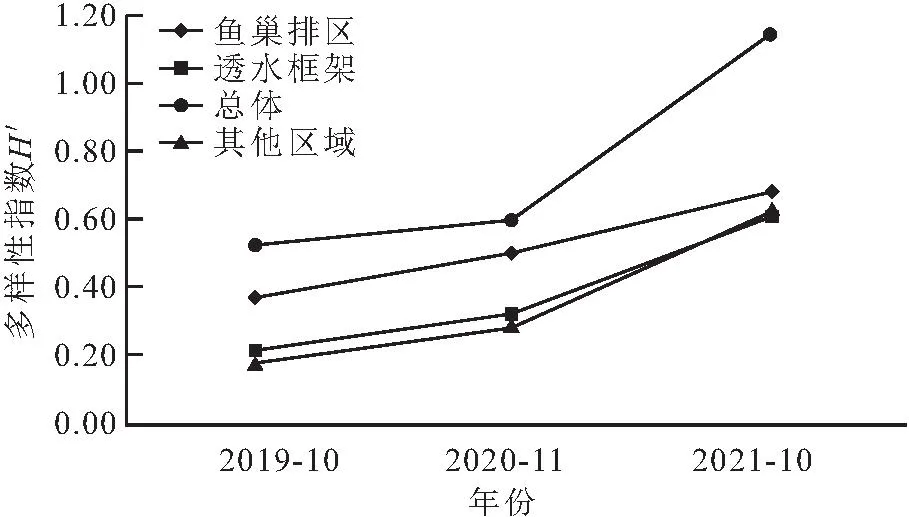
图5 不同区域底栖动物多样性指数(H′)变化Fig.5 Changes of benthic diversity index(H′)in different regions

图6 不同区域底栖动物均匀度(J)指数变化Fig.6 Changes in the evenness(J)index of benthic animals in different regions
鱼巢排、透水框架区域D、H′均大于其他区域和总体水平,且透水框架区域指数大于鱼巢排区。鱼巢排区域J在调查时间内均大于透水框架区、其他区域、总体区域。总体水平、透水框架区、其他区域J差异性不大。
3 讨论
3.1 生态涵养的积极意义
底栖动物物种数目、密度和生物量呈现逐年上升的趋势,可能由于生态涵养区随着年份的增长,降低了水体平均流速,改变了生物栖息地的底质状况[16,17],进而促进了底栖动物的生存。优势种通常控制群落中的能量流动和物质循环[18],时间尺度上优势种的变化与生物群落的演变密切相关[19]。2019年优势种主要为甲壳纲的日本大螯蜚、昆虫纲的摇蚊类,且摇蚊类优势度最大,这是由于在生态涵养区建设初期,生态涵养区涵养效果不明显,水体流速较快,而昆虫纲的摇蚊类适应于多种底质[20],在流水区域占优势[21],因此成为主要的优势种之一。2021年底栖动物优势种分属于甲壳纲的日本大螯蜚、多毛纲的围沙蚕一种,多毛纲代替昆虫纲成为优势种是因为小型多毛类主要生活在软泥底质,与沉积物有机质[22]含量密切相关,莲花洲港生态涵养区透水框架、鱼巢砖的应用,降低了水体流速,促进了有机质的沉积[23,24],营造了有利于多毛类底栖动物生存的环境。
3.2 不同生态涵养设施导致的生态涵养效果
调查结果显示不同时期、不同区域采集到的底栖动物物种种类、优势种差异比较明显。其中,透水框架区、其他区域2021年底栖动物种类数均大于2019年,段学花[25]研究表明流速对底栖动物群落的现存量和种类组成有较大的影响,本研究环境监测结果显示透水框架区、其他区域年平均流速逐渐降低,这可能是底栖动物种类变化的主要原因之一。
鱼巢排区底栖动物平均密度呈现逐年上升的趋势,且在2021年10月鱼巢排区底栖动物平均密度大于总体平均水平,说明鱼巢排区有利于底栖动物的生存,且在2021年效果显著。透水框架区底栖动物平均密度在2019年低于鱼巢排区[26],2010-2021年均高于鱼巢排区,且在2021年10月调查中,底栖动物平均密度显著大于鱼巢排区,这是由于透水框架区营造的水体环境优于鱼巢排[27],且随着时间效应的影响,透水框架生态涵养效果越来越显著。 鱼巢排区、透水框架区底栖动物生物量均呈现逐年上升的趋势,进一步验证了生态涵养区对底栖动物的生态涵养效果,其中,透水框架区底栖动物生物量虽呈逐年显著增长,但是一直低于总体水平,这一结果与ODUM[28]和JORGENSON等[29]的假设相反,本研究结果显示东流水道莲花洲港底栖动物生物量恢复的较慢。而章飞军等[8]研究表明,系统信息(结构能质)恢复的非常快,生物量最慢,一直到研究末期生物量仍低于对照区,与本研究结果相似,本研究结果显示,2019年、2020年、2021年透水框架区底栖动物密度均高于其他区域,但底栖动物生物量在2021年仍显著低于其他区域。
物种种类多样性与多种环境因素相关,在不同的时间和空间尺度上的自然或人为扰动都会导致底栖动物群落组成和生物多样性的变化[30]。总体水平上,底栖动物丰富度指数(D)、多样性指数(H′)、均匀度指数(J)均呈现逐年上升的趋势,进一步说明生态涵养区改善了底栖动物生存的环境,促进了底栖动物的生存。其他区域底栖动物多样性指数(H′)、均匀度指数(J)在2019年、2020年均小于总体水平,到2021年数值趋于接近,说明生态涵养区促进了其他区域底栖动物的恢复,且在2021年后底栖动物多样性指数(H′)、均匀度指数(J)有超越总体水平的趋势。鱼巢排区各指数增长速度较快,透水框架区底栖动物平均密度、平均生物量相对于其他区域较高,但各指数增长速度缓慢,可能由于透水框架创造的水体环境,主要对昆虫纲、多毛纲、甲壳纲某几类底栖动物生存有利,也可能是研究年限有限,不能更全面地展现透水框架区对底栖动物的影响。要想进一步探究透水框架各指标的变化,可以对该结构区域进行进一步的监测。
