基于NLRP3炎症小体介导的细胞焦亡探讨针康法对缺氧缺血性脑损伤新生大鼠的影响
2024-02-01丁小连单昊宇康玉莹
刘 波,丁小连,单昊宇,康玉莹,白 露
1.黑龙江中医药大学附属第二医院,黑龙江 哈尔滨 150001;2.黑龙江中医药大学,黑龙江 哈尔滨 150040;3.齐齐哈尔医学院,黑龙江 齐齐哈尔 161003
新生儿缺氧缺血性脑损伤(Hypoxic-ischemic brain damage,HIBD)是指围生期由于脑内氧供给和血流量的不足所致的脑组织损伤[1],是造成新生儿长期残疾的主要神经系统疾病之一。在缺血缺氧后的几个小时内,大脑组织会发生氧化应激、炎症反应等一系列的神经毒性损伤,造成神经细胞的广泛死亡与神经功能缺损。炎症小体活化是神经炎性反应中必不可少的一步,同时也是神经细胞死亡的主要诱因。细胞焦亡是一种典型依赖于炎症反应的程序性细胞死亡方式[2]。核苷酸结合寡聚化结构域样受体蛋白3(NLRP3)炎性小体可诱导细胞焦亡,其机制[3]是通过切割半胱氨酸天冬氨酸酶1(Caspase-1)以及促进白细胞介素(IL)-1β、IL-18的成熟和消皮素D(GasderminD,GSDMD)的裂解来增加促炎因子的释放。研究证实[4],细胞焦亡触发的无菌性炎症与HIBD的发生与发展紧密相关,抑制NLRP3介导的细胞焦亡可改善脑损伤。在头穴丛刺长留针的过程中对HIBD患儿进行康复功能训练,可有效改善患儿的运动功能障碍,但其作用机理尚未明确。因此,本研究通过构建新生大鼠HIBD模型,以探讨针康法是否通过抑制NLRP3炎症小体介导的细胞焦亡发挥脑保护作用,以进一步探究其改善HIBD后运动功能缺损的潜在机理。
1 材料与方法
1.1 实验动物及分组
新生7 d龄Wistar大鼠120只,体质量10~15 g(辽宁长生),实验动物饲养于黑龙江中医药大学脑功能与神经康复实验室,温度(23±1)℃,湿度(50±10)%,12 h光暗循环,动物自由饮水和摄食。随机将其分成5组:正常对照组(A组)、模型对照组(B组)、头穴丛刺组(C组)、康复训练组(D组)和针康组(E组),每组再分成24 h、7 d和14 d共3个亚组。本实验经黑龙江中医药大学实验动物伦理委员会审批(批准号:20210911-06)。
1.2 仪器与试剂
1.2.1 仪器 8%O2+92%N2混合气体(哈尔滨鼎新工业气体);转棒疲劳仪(上海欣软科技有限公司);针灸针(0.25mm×25mm,苏州医疗用品厂有限公司);荧光显微镜(德国leica公司);2135切片机(德国leica公司);DYY-6C电泳仪(北京六一生物科技有限公司)。
1.2.2 试剂 兔抗单克隆抗体NLRP3、Caspase-1及Alexa Fluor594标记的山羊抗兔IgG(北京博奥森生物科技有限公司,批号分别是Bs-10021R、Bs-10743R与Bs-0295G-AP);兔抗单克隆抗体GSDMD-N端(美国Abcam公司,批号ab-215203);TUNEL荧光试剂盒(大连美仑生物科技有限公司,批号MA0223-1)。
1.3 造模方法
采用改良RICE法制备HIBD模型[5]。7 d新生大鼠吸入异氟烷麻醉后,用医用胶布将其仰卧位固定在手术台上,颈正中位置剪开皮肤。剥离该切口皮下的软组织,暴露出左侧颈总动脉并分离与其伴行的迷走神经及静脉,5-0结扎线穿入左颈总动脉的底部做双重结扎以阻断血流。在完成结扎后,剪断两根结扎线之间的血管,缝合切口,用碘伏对缝合处进行擦拭消毒,解开固定,将新生大鼠放入37 ℃恒温箱中,休息2 h。随后将大鼠放置在含8%氧气和92%氮气的37 ℃缺氧箱中2 h,在此过程中,缺氧箱的气体流速保持在0.5 L/min。A组仅行左颈总动脉游离,未作结扎、缺氧处理。模型制备完成后,将新生大鼠放回母鼠笼中,自由饮食水。
1.4 模型评价
参照Longa评分,对制备HIBD模型的新生大鼠进行功能评估[6],评价模型是否成功,具体如下:0分:活动正常,神经功能未缺损;1分:患侧的前爪不能充分伸展,呈现轻度缺损状态;2分:活动时,躯干向患侧转圈,呈现中度缺损状态;3分:活动时,躯干向患侧倾倒,呈现重度缺损状态;4分:失去意识且不能自主活动。
评定结束后,将1~3分的新生大鼠纳入实验,其他予以剔除。
1.5 干预方式
C组采用于致顺的头穴丛刺法[7],选择百会及其左、右侧各旁开2 mm处(顶区)向前平刺至顶前区,针刺深度约15 mm,捻转1 min,200 r/min,留针2 h,1次/d。D组进行转笼转棒训练。①转笼训练:采用长55 m、直径45 cm的滚筒装置,以6 r/min的速度进行训练,每日训练10 min。②转棒训练:运用转棒疲劳仪以20 r/min的速度进行训练,每日训练10 min。E组在造模完毕后进行头穴丛刺的基础上进行康复训练。其余组不予干预。
1.6 观察指标及检测方法
1.6.1 运动功能缺损评分 不同时间点采用网屏实验对各组大鼠运动功能进行评估[8]。网屏采用40~50 cm网片,网格1 cm×1 cm,除了下侧的部分都有木质边框,把大鼠置于其上。2 s内将网屏翻转90°呈竖直状态,并维持5 s。
评分时大鼠的前爪锁住网线5 s,未脱落为0分;大鼠的前爪暂时锁住网线,下落一小段距离,但不掉落为1分;大鼠的前爪锁住网线并留在网屏上5 s为2分;当网屏转动时,大鼠立即掉下为3分。
1.6.2 神经细胞焦亡的测定 TUNEL+Caspase-1免疫荧光双染色检测神经细胞焦亡。在对应的时间点每组取4只大鼠,将大鼠吸入异氟烷进行麻醉。麻醉后把大鼠仰卧位固定,剪开腹部皮肤,将心脏完全露出,进行心脏灌注,直到大鼠的肝脏、肺脏以及四肢出现苍白僵硬的状态为止。快速在冰上断头取脑,将脑组织完整取出来。用模具从中间切成前后两部分,将包含有海马的后半部分用4%多聚甲醛固定后于4 ℃保存。脑组织脱水、包埋制备石蜡切片。切片脱蜡至水,抗原修复,双氧水、血清封闭后,将Caspase-1(1∶500)滴入切片,将切片置于4 ℃下的湿盒中孵育一夜;按顺序添加5 μL的TdT酶、CY3-TSA(1∶500);将50 μL的TUNEL反应液滴于切片上,37 ℃避光培养60 min;DAPI复染细胞核,抗荧光淬灭封片剂封片。显微镜下观察并拍摄4只大鼠海马区同一位置。计算细胞共表达率(三色重叠细胞/蓝色荧光细胞)为细胞焦亡百分率。
1.6.3 Western blot法检测NLRP3、Caspase-1和GSDMD-N蛋白 干预结束后每组取4只大鼠,待大鼠在相应时间点吸入异氟烷进行麻醉后,快速断头取脑,迅速分离海马组织。脑组织样本取出后,置于冷冻管中,将其放入液氮中速冻15~20 s。上述完成后,将冷冻管放于-80 ℃冰箱,等待后续检测。测定样品的质量,加入相应体积的裂解液提取脑组织总蛋白,BCA法蛋白定量。用40 μg的蛋白煮沸变性,SDS-PAGE后,在37 ℃下5%的脱脂奶粉下封闭1 h。滴加稀释I抗(NLRP3∶1∶1 000;Caspase-1∶1 000;GSDMD-N∶1∶2 000)和抗β-actin I抗(1∶1 000),于4 ℃下培养过夜。次日用TBST冲洗5次后,加入羊抗兔IgG-HRP(1∶5 000),在37 ℃下孵育45 min。清洗后ECL显影,用凝胶图像处理系统分析计算目标蛋白质的吸光度,用NLRP3、Caspase-1和GSDMD-N的吸光度与β-actin的吸光度之比来定量目标蛋白的表达。
1.7 统计学处理
采用SPSS26.0进行分析,采用均值±标准差表示;用单因素方差分析进行多组间比较,用LSD-t检验进行两两比较;以P<0.05为差异有统计学意义。
2 结果
2.1 各组运动功能缺损评分
参照网屏实验,在各个时间点,对5组新生大鼠分别进行运动功能缺损评分。其中,相较于A组,其他4组新生大鼠在各个时间点的网屏实验评分显著升高,差异具有统计学意义(P<0.05);相较于B组,各治疗组的网屏实验评分在各个时间点均明显降低,差异具有统计学意义(P<0.05);在各时间点,E组的网屏实验评分与C组和D组相比较均降低,差异具有统计学意义(P<0.05)。见表1。
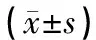
表1 各组新生大鼠网屏实验评分
2.2 各组海马区神经细胞焦亡情况
对各组大鼠的海马区同一位置进行TUNEL+Caspase-1免疫荧光双染,计算在各个时间点的阳性细胞焦亡率,A组可见少量焦亡细胞;相较于A组,其余4组新生大鼠在各个时间点,焦亡细胞率显著增加,差异具有统计学意义(P<0.05);相较于B组,各治疗组在各个时间点神经细胞焦亡率明显降低,差异具有统计学意义(P<0.05);相较于C组和D组,E组神经细胞焦亡率显著降低,差异具有统计学意义(P<0.05)。见图1、表2。
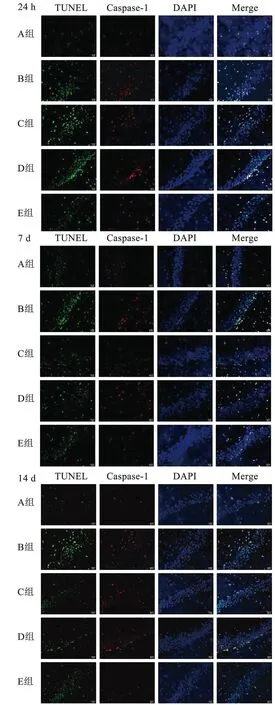
图1 新生大鼠海马区细胞焦亡比较(20×)

表2 各组新生大鼠细胞焦亡的比较
2.3 各组脑组织海马区NLRP3、Caspase-1及GSDMD-N蛋白表达情况
对各组大鼠的大脑海马区进行Western blot检测,观察在各个时间点NLRP3、Caspase-1及GSDMD-N表达情况,相较于A组,其余各组大鼠在各时间点的NLRP3、Caspase-1及GSDMD-N表达量明显增加,差异具有统计学意义(P<0.05);相较于B组,各治疗组在各个时间NLRP3、Caspase-1及GSDMD-N表达量明显减少,差异具有统计学意义;相较于C组和D组,E组的NLRP3、Caspase-1及GSDMD-N表达量在各个时间点显著减少,差异具有统计学意义(P<0.05)。见图2~4、表3~5。
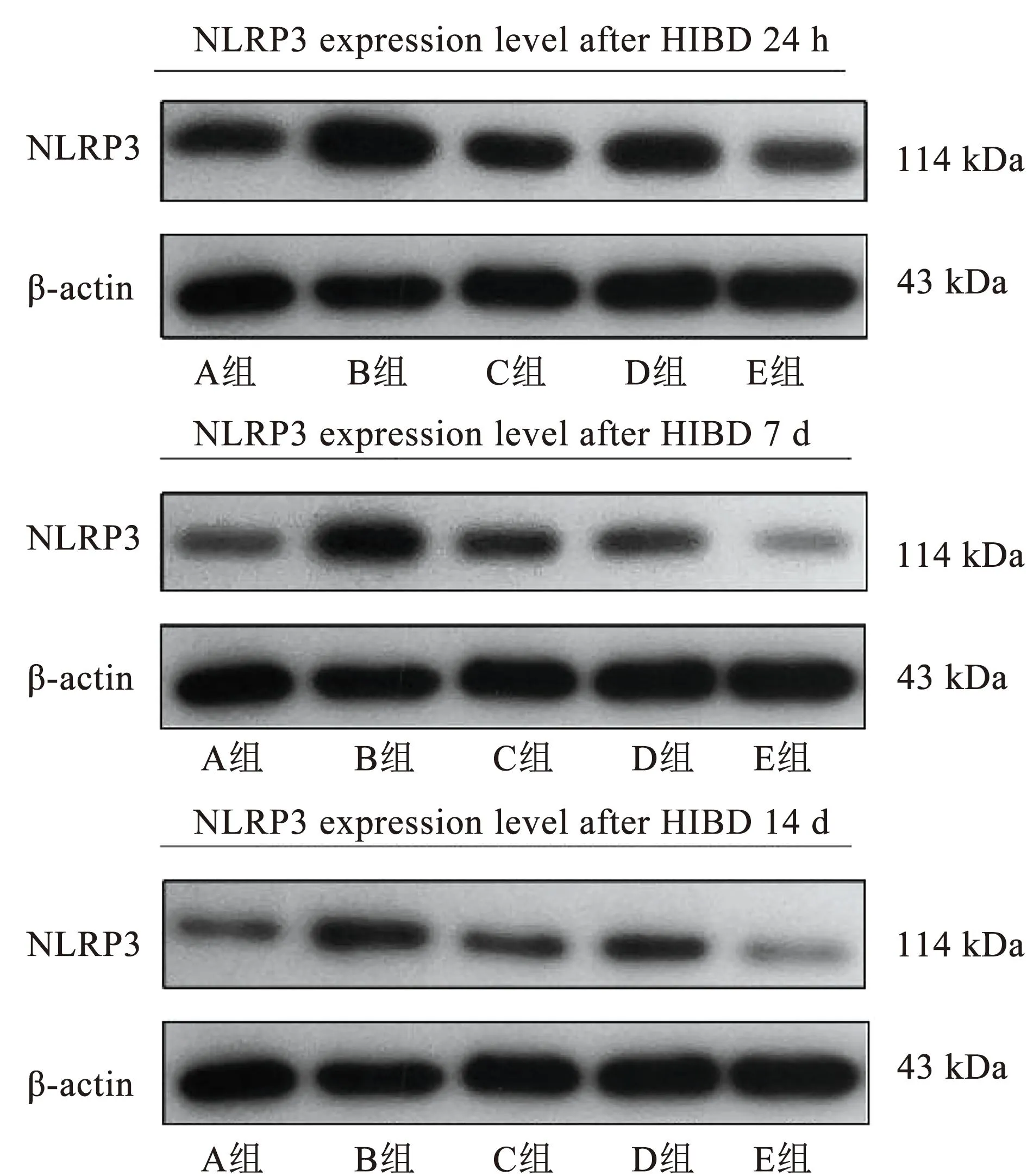
图2 各组新生大鼠NLRP3的表达情况

图3 各组新生大鼠Caspase-1的表达
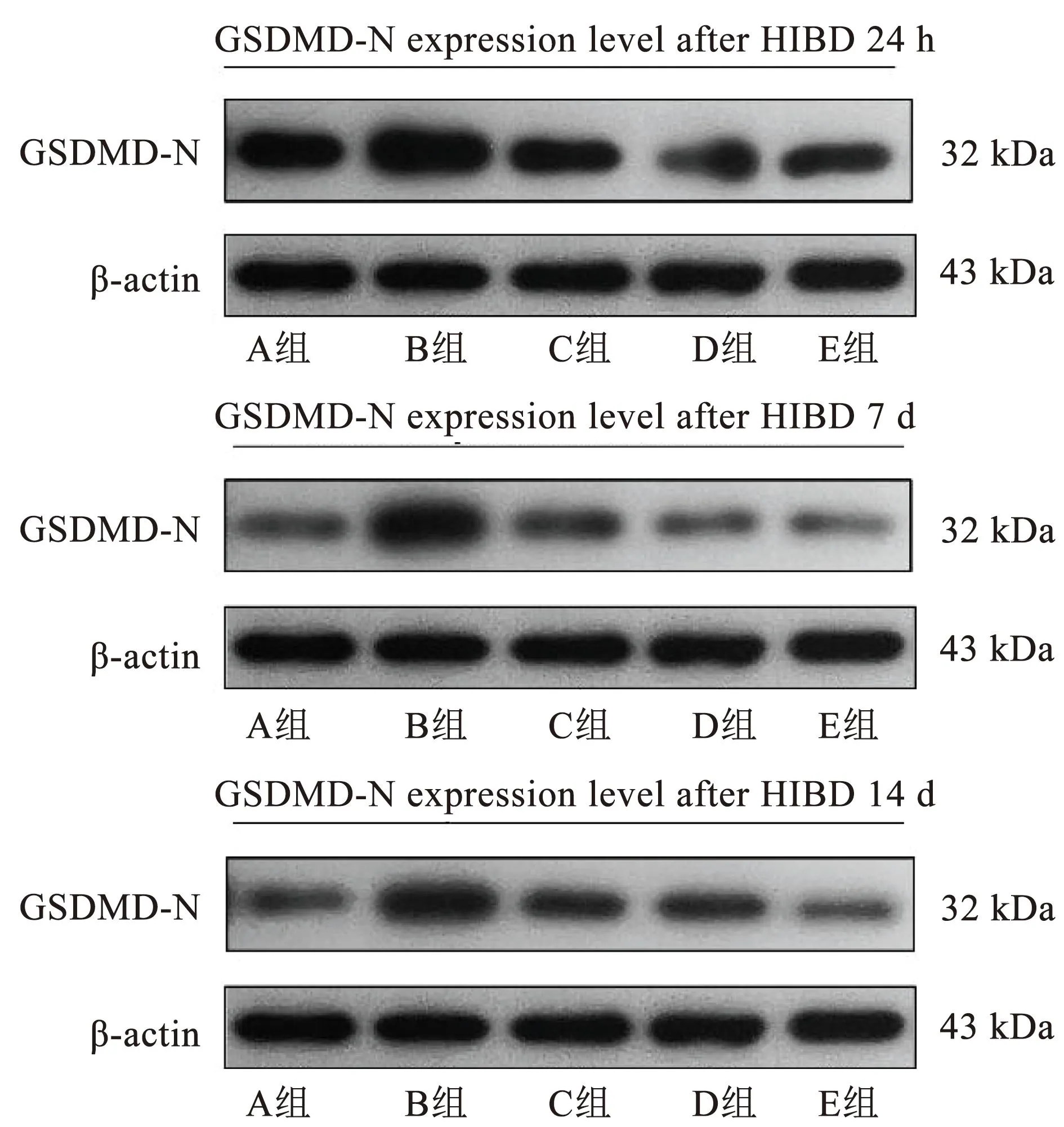
图4 各组新生大鼠GSDMD-N端的表达
表3 各组新生大鼠NLRP3的表达情况
表4 各组新生大鼠Caspase-1的表达情况
表5 各组新生大鼠GSDMD-N的表达情况
3 讨论
新生儿缺氧缺血(Hypoxic ischemia,HI)可导致不同程度的脑损伤,未成熟脑组织对HI的刺激尤为敏感[9]。HIBD可引起神经系统后遗症,如患儿可有运动障碍、发育迟缓、癫痫发作、认知功能障碍及感觉障碍等神经系统的后遗症[10],在严重的情况下可导致死亡。目前,亚低温治疗被认为是在围产期治疗新生儿脑损伤的唯一标准的疗法,只有50%的治疗是有效的,炎性敏感的HIBD新生儿的效果较差。因此,为减轻发病后新生儿脑损伤程度和功能障碍,降低永久性神经系统损伤发生率,仍然有必要探索新的治疗方法。
《灵枢·经脉》载:“人始生,先成精,精成而脑髓生”,阐明了要将肾之精华转化为髓进而填充大脑才能发挥功能,而脑髓的生成也与脾的精气息息相关,故中医学者认为本病病位在脑,其基本病因与先天胎禀不足、肝肾亏虚和脏腑气血功能失调密切相关,而脑髓损伤等后天因素则进一步促进了本病的发生和发展[11-12]。近年来,中医对缺血性脑损伤的治法逐渐趋于多样化,包括针灸、方药、中医传统功法和中西医结合的综合疗法等。唐强教授通过对各种针灸方法与现代康复技术结合治疗缺血性脑损伤的联合时机、联合点等问题的理解创新后,提出了“针康同步、动态治疗与整体康复”的中西医结合康复理念,创立了针康法治疗技术[13],即在患者头穴丛刺长留针期间进行康复训练,以达到最佳的治疗效果。
近年来,针康法在临床缺血性脑损伤的治疗中已经展现出较好的疗效,可以显著地改善缺血性脑损伤后遗症的各种功能异常,从而提升患者的生活质量。前期实验研究发现[14-17],针康法可以通过促进血管新生、抑制细胞自噬、降低细胞凋亡、调节肠道菌群和减轻脑部炎症损伤,起到了神经保护的作用。研究发现,细胞焦亡参与了HIBD发生发展的整个过程。而对于针康法是否具有拮抗细胞焦亡作用尚未报道,问题一直处于搁置状态。为了深化针康法治疗HIBD具体的作用机理,于是本课题进行了此次有针对性的研究。
炎症小体是细胞浆内识别系统参与的组装多蛋白的复合体,是介导HIBD后中枢神经系统炎症反应和细胞焦亡的关键因素之一[18]。其中,作为炎症反应核心的NLRP3炎性小体是被研究的最透彻的一种炎性小体。NLRP3作为受体蛋白通过其效应结构域招募ASC,并由ASC的CARD结构域的NLRs募集Caspase-1的前体,从而形成NLRP3炎性小体[19]。NLRP3炎症小体切割Caspase-1诱导其活化,触发炎症因子pro-IL-1β、pro-IL-18各自剪切成IL-1β及IL-18并释放到胞外发挥促炎作用[20]。在脑缺血缺氧后,会产生大量的活性氧,对细胞结构和线粒体功能造成损害,进而导致脂质过氧化反应增强。损伤的线粒体和残留的细胞碎片或分解的蛋白质,可引发NLRP3炎症小体的活化,导致炎症级联反应。细胞焦亡是Caspase-1依赖的一种细胞炎性死亡模式[21],这种死亡模式可能会引起机体炎症级联反应。经典细胞焦亡途径主要依靠激活的炎症小体活化Caspase-1切割GSDMD,促使GSDMD-N端片段与质膜内层结合,形成膜孔并诱导焦亡[22]。NLRP3是触发细胞焦亡的关键蛋白,能够与 ASC产生相互作用,并募集pro-Caspase-1,再经过上述Caspase-1经典焦亡途径,将焦亡信号传递给下游的焦亡蛋白GSDMD,GSDMD-N打孔从而导致细胞溶胀破裂,释放出大量的炎性介质,扩大炎症反应,诱导更多细胞损伤[23]。欧阳昕等[24]发现针刺脑缺血大鼠 “百会”“大椎”“足三里”穴,可降低大脑皮质NLRP3、Caspase-1的表达和IL-1β含量,抑制神经细胞焦亡。此外,杨小惠发现[25]对脑缺血大鼠给予丰富环境刺激干预,发现通过上调缺氧诱导因子-1α的表达可抑制NLRP3炎症小体激活,进而调控细胞焦亡的发生和发展。综上,针刺或康复训练都可抑制脑缺血大鼠抑制NLRP3炎症小体的激活,从而调控细胞焦亡,促进脑损伤后神经修复。
细胞触发焦亡时,形态上既有坏死性又有凋亡的特点,其过程类似于凋亡,但也伴随着核聚集、染色质DNA破碎等病理改变。TUNEL法检测细胞凋亡和细胞焦亡都呈阳性,而细胞焦亡则是Caspase-1所介导。故当Caspase-1及 TUNEL免疫荧光染色均呈阳性时(核内红/绿双阳性),可判定为细胞焦亡[26]。因此,本研究采用此方法对HIBD大鼠脑组织海马区进行免疫荧光双染,发现缺氧缺血处理后的大鼠海马区神经细胞焦亡率增加。此外,各治疗组相较于B组细胞焦亡率降低,E组最为显著。WB检测发现C组、D组和E组在各时间点与B组相比较, NLRP3、Caspase-1和GSDMD-N端蛋白的表达水平降低,其中E组的疗效最明显,这与荧光显微镜观察到的结果一致。网屏实验结果显示,相较于B组,C组、D组和E组的网屏实验评分明显降低,提示针康法能促进HIBD大鼠运动功能恢复。以上结果表明针康法可拮抗NLRP3炎症小体激活,调控细胞焦亡的发生,从而改善缺氧缺血性脑损伤后运动功能障碍。有研究称甲基莲心碱通过抑制NLRP3炎症小体途径对HIBD大鼠发挥拮抗细胞焦亡作用[27],与本研究结果一致。但针康法对NLRP3炎症小体的具体抑制通路及对其他炎性小体的调控机制仍需进一步探索。
