不同重金属污染水平下生物结皮及下层土壤细菌群落的差异性分析
2024-01-25王融融丛士翔常文静余海龙黄菊莹
樊 瑾,王融融,丛士翔,常文静,余海龙,*,黄菊莹
1 宁夏大学地理科学与规划学院,银川 750021 2 西北农林科技大学水土保持研究所,杨凌 712100 3 宁夏大学生态环境学院,银川 750021
随着近些年矿产资源开发与工业进程的加快,带来巨大经济效益的同时也产生了大量重金属污染物,它们通过干、湿沉降进入周边土壤或被植物叶片直接吸收,引起土壤和植物中As、Cd等多种重金属含量超标,甚至进入食物链威胁人类健康[1-2]。由于重金属的隐蔽性高、易累积、难降解且毒性强,进行重金属污染修复工作显得亟不可待。目前常用的修复方法有客土法、土壤淋洗法、植物修复法、生物炭修复法等[3]。然而,这些方法在实际的修复效果中都存在成本高、单一手段修复效率低等问题。微生物对土壤中微量金属离子灵敏度高、特异性强,兼具环保、经济的优点,在环境监测和重金属污染治理中有着广泛的应用前景[4]。尤其是土壤细菌对土壤环境质量变化敏感[5],可通过土壤细菌群落结构及多样性的变化对土壤是否被污染、污染的程度、污染的效应和风险做出及时、科学的客观判断[6]。
生物结皮(Biocrusts)是由土壤颗粒与细菌、真菌、藻类和苔藓植物相互作用形成的有机复合体[7]。作为尾矿地植被恢复过程中的先锋拓殖生物体和重要的地表覆盖物,一方面生物结皮中的植物体藻、苔藓具有独特的生物构造被认为是良好的重金属富集材料[8-9],另一方面生物结皮通过吸附和截留大气降尘和溶解于降水中的矿物质,促进土壤颗粒细化,改善土壤结构,为微生物和土壤动物提供适宜的微生境[10],因此被视作潜在修复重金属污染的理想材料。
有研究表明,细菌能够通过胞内聚集、矿化沉淀、吸附溶解等方式,改变土壤重金属赋存形态,调控其生物有效性[11],影响植物对重金属的吸收和转运,直接或间接活化吸收或固化土壤中重金属[12]。然而,现有研究多聚焦于特定菌种-超积累植物的狭隘组合,成本高且植物的移栽过程繁琐,修复效果往往差强人意。目前应用生物结皮修复重金属污染物的相关研究较少,对其可能的修复效果和机制并不明确。此外,人们对于重金属污染过程如何驱动生物结皮及其下层土壤细菌群落变化知之甚少,对于不同重金属污染背景下生物结皮层及下层土壤细菌群落的异同及原因鲜见报道。这在一定程度上制约了人们对微生物在修复退化生态系统过程中的贡献及作用的科学评估。
基于此,本研究以宁夏东部荒漠草地和工矿区为研究区域,选取自然分布的2种生物结皮(藻结皮和苔藓结皮)为研究对象,探究生物结皮富集重金属效应,分析不同重金属污染程度下生物结皮层及下层土壤细菌群落的结构、组成、多样性,探讨哪些关键的环境因子驱动了细菌群落的变异。研究结果旨在揭示生物结皮及其下层土壤细菌群落多样性和结构对不同土壤重金属污染水平的响应及重金属污染的生物指示程度,为重金属污染的微生物修复提供数据支撑。
1 材料与方法
1.1 研究区概况
研究区地处宁夏东部,属典型温带大陆性气候,年均降水量190.4 mm,年均蒸发量2088.2 mm,年均温6.7-8.8℃。土壤类型主要为灰钙土和风沙土,质地偏砂,土层厚度约为50 cm,结构松散。受水分条件限制,植被以耐旱、耐寒、耐土壤贫瘠的灌木和草本植物为主,自然形成的生物结皮广泛分布,以藻类和地衣类结皮为主,藓类结皮大多以斑块状分布。
1.2 样地设置及样品采集
依据生产活动类型差异造成的重金属污染水平差异,本研究设置工矿区和封育区(图1)。其中工矿区位于宁东基地核心区的马莲台电厂周边(37°48′N,107°21′E)。马莲台电厂是宁夏发电集团公司独资建设的第一个大型火力发电厂,于2005年投产,设计规模为2×330MW+2×600MW+2×1000MW。在盛行风向(西北风)的下风向,选取地形相对平坦且无高大树木遮挡(为避免地形、植被对本研究的影响)的生物结皮分布区作为研究样地。依据电厂产生的燃煤烟尘排放距离、方向,将位于电厂东南方向100 km处杨寨子村的封育草地设为封育区(周边无工业活动)。两个样地均发育有大面积的生物结皮,且其组成和类型基本一致(表1)。依据生物结皮中优势类群的差异,选取发育良好的藻结皮和藓结皮。其中,藻结皮在自然状态下多为黑色,成片分布;藓结皮在自然状态下为深褐色,雨后为绿色,多呈斑块状分布。

表1 样地基本信息Table 1 Basic information of the two sample plots

图1 研究区位置Fig.1 Location of study area
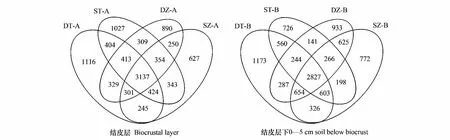
图2 生物结皮A、B层土壤细菌群落OTU的Venn图Fig.2 Venn diagram of the OTUs of the soil bacterial communities in A and B horizons of biocrustsSZ-A: 工矿区藻结皮层 Algal biocrustal layer in mining area; DZ-A: 封育区藻结皮层 Algal biocrustal layer in enclosed area; ST-A: 工矿区苔藓结皮层 Moss biocrustal layer in mining area; DT-A: 封育区苔藓结皮层 Moss biocrustal layer in enclosed area; SZ-B: 工矿区藻结皮层下土壤 Soil below Algal biocrust in mining area; DZ-B: 封育区藻结皮层下土壤 Soil below Algal biocrust in enclosed area; ST-B: 工矿区苔藓结皮层下土壤 Soil below Moss biocrust in mining area; DT-B: 封育区苔藓结皮层下土壤 Soil below Moss biocrust in enclosed area;本研究以0-1 cm为生物结皮层,标记为A层;生物结皮层下0-5 cm为结皮下层,标记为B层
据前期研究工作[13],2类生物结皮的厚度均小于1 cm(藻结皮2.54-3.08 mm,藓结皮6.13-7.23 mm),因此本研究以0-1 cm为生物结皮层,标记为A层;生物结皮层下0-5 cm为结皮下层,标记为B层。
采样工作于2019年10月进行,在工矿区和封育区各布设3个生物结皮的典型样地(40 m×40 m,间隔至少50 m),每个样地内设置3个典型的子样方(10 m×10 m,间隔至少15 m),子样方中同时包含藻结皮和藓结皮斑块。采用5点采样法,先用木铲铲取A层结皮,再挖取B层土壤,之后将同一土层的5个小斑块所采样本混匀并标记好。因结皮斑块面积大小不等,取样斑块至少为6 cm×6 cm。共采集24个样本(A层样本12个,B层土壤样本12个,n=3),所有样本置于冰盒内保存,迅速带回实验室进行后续分析。
1.3 样本测定
将采集的样本各取10 g置于-20℃冰箱,短暂保存进行DNA提取和后续研究。剩余样本风干,测定pH、电导率、有机碳等指标。土壤基本理化指标的测定参考《土壤农化分析》。重金属(Zn、As、Cd、Cr、Pb、Hg)含量的测定采用密封微波消解体系消解样本,利用电感耦合等离子体质谱仪(ICP-MS)检测消解液中重金属浓度。
1.4 重金属污染负荷指数分析
本研究运用重金属污染负荷指数(Pollution Load Index)法[14]对生物结皮A、B层重金属进行污染评价。
1.5 样本DNA提取、扩增和和高通量测序
根据Power SoilTM DNA试剂盒说明书提取土壤总DNA。之后利用1%琼脂糖凝胶电泳检测DNA的纯度和浓度,用无菌水稀释样本至1 ng/μL。以稀释后的基因组DNA为模板,采用通用引物515F(5′-GTGCCAGCMGCCGCGGTAA-3′)和806R(5′-GGACTACHVGGGTWTCTAAT-3′)对细菌16S rDNA-V4区域进行PCR扩增。每一样本的引物均含有特异的Barcode序列用于区分样本序列。测序由北京诺禾致源Illumina NovaSeq PE250测序平台完成。
1.6 数据处理和分析
采用FastQC软件对原始测序数据的进行质控,过滤去除低质量Reads。采用FLASH(V1.2.7)进行序列拼接、QIIME(V1.7.0)进行序列再过滤。采用UPARSE软件在97%的相似水平下进行可操作分类单元(OTU,Operational Taxonomic Units)的聚类。样品中的微生物多样性和各类微生物的丰度都是基于OTU的分析。采用RDP classifier(V2.2)对照Silva数据库以80%置信度对OTU代表序列进行物种注释。为避免各样本生物量的差异,按最小样本序列数进行样本序列的抽平处理,得到标准化数据进行后续统计分析。基于OTU结构分析α多样性、β多样性及优势菌群差异,评估不同重金属污染水平下两类生物结皮A、B层土壤细菌群落结构和多样性差异。其中,细菌群落的α多样性指标(Chao1、ACE、Shannon-Weiner和Simpson指数)通过Qiime平台计算。β多样性是基于两两样本间距离矩阵反映样本间的差异,通过PCoA进行可视化展示[15]。
采用Origin 2021绘图。在数据符合方差齐性的条件下,采用单因素方差分析(LSD,Duncan)进行不同处理间的多重比较,反之采用非参数检验Mann-Whitney法。采用Spearman相关系数分析优势细菌门与土壤理化性质的相关性。采用Canoco 5.0进行土壤理化因子和细菌群落变化的冗余分析。图表中数据为平均值±标准误差(n=3)。
2 结果与分析
2.1 生物结皮A、B层土壤理化性质
所有样本的pH值均大于7,土壤偏碱性(表2)。A层和B层的pH、电导率、有机碳、全氮、全磷、速效氮、速效磷、黏粒和粉粒都有显著差异,表现出随着生物结皮的演替,pH逐渐降低,有机碳、养分逐渐增高且A层始终高于B层(P<0.05)。其中有机碳和速效氮变化幅度最大。同一土层中,工矿区生物结皮的有机碳、全氮、全磷、速效氮、速效磷、黏粒和粉粒高于封育区,而pH、砂粒含量低于封育区。
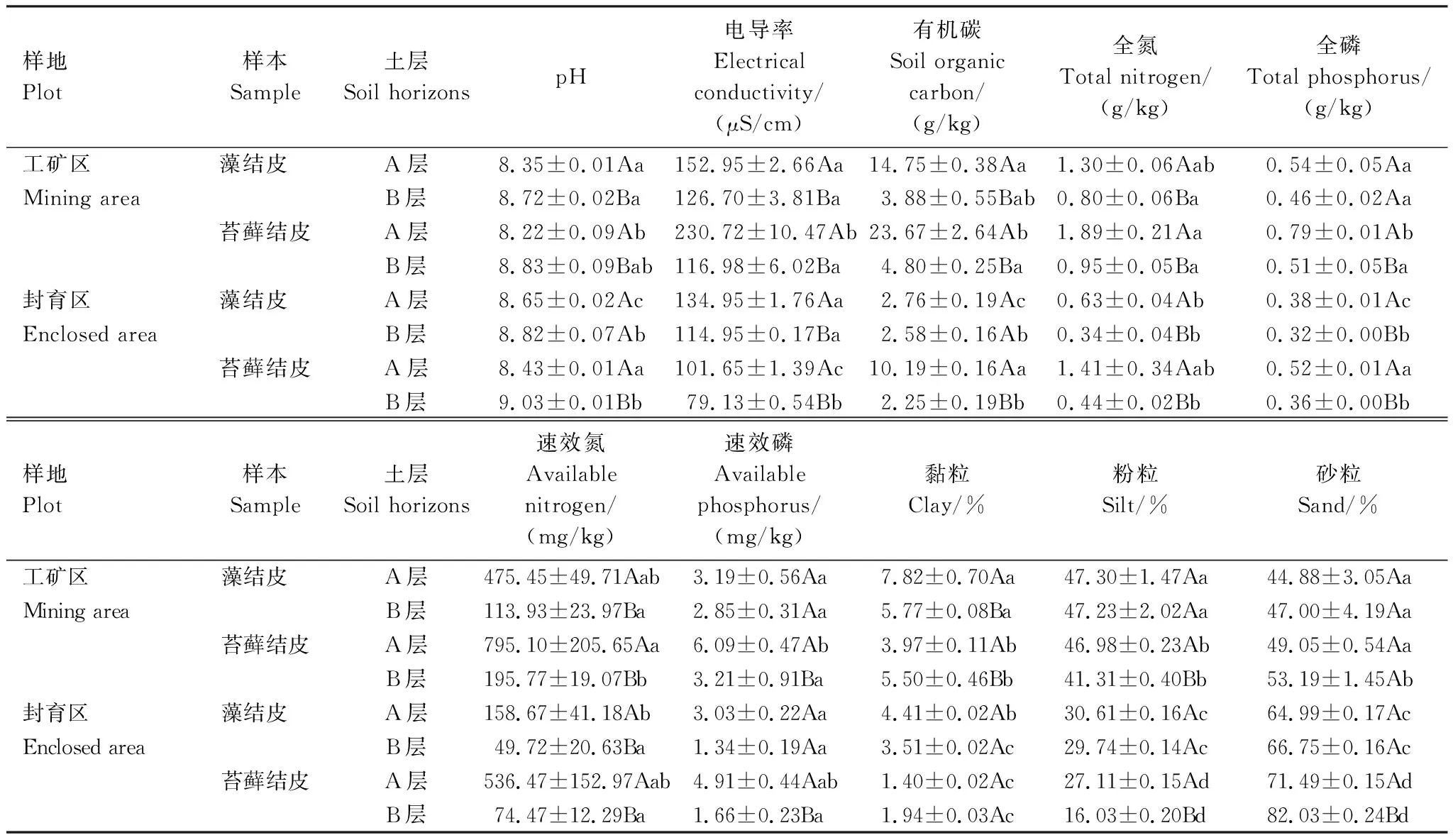
表2 生物结皮A、B层理化性质Table 2 Physical and chemical properties of A and B horizons of biocrusts
2.2 生物结皮A、B层土壤重金属及污染评价
工矿区和封育区生物结皮A、B层土壤中重金属含量见表3。总体表现为工矿区生物结皮A、B层土壤中重金属含量显著高于封育区,生物结皮A层和B层的Cd和Zn含量差异显著(P<0.05)。从藻结皮向藓结皮演替过程中,生物结皮富集重金属的能力呈增强趋势。本研究以宁夏土壤背景值[16]作为重金属污染程度评价标准进行参比评价。结果显示该区域生物结皮的6类重金属元素均存在不同程度污染(表4)。2个样地中,所有样本重金属元素的最高污染系数(CFi)表现出一致的规律:Hg>Cd>As>Cr>Zn>Pb,污染负荷指数(IPL)排序结果为苔藓结皮A层>苔藓结皮B层>藻结皮A层>藻结皮B层。说明重金属对生物结皮层及其下土壤均有影响。IPLZONE的结果表明工矿区为极强污染程度。

表3 生物结皮A、B层土壤重金属含量Table 3 Heavy mental contents of A and B horizons of biocrusts

表4 生物结皮A、B层土壤重金属污染评价Table 4 Evaluation of heavy metal pollution in A and B horizons of biocrusts
2.3 生物结皮及其下土壤细菌群落α多样性
每一个OTU代表一种微生物。根据细菌OTU数目类群划分,可区分不同生境中生物结皮中细菌的种群。α多样性表征微生物群落的丰富度和多样性,Chao1指数和ACE指数表征物种丰富度,Shannon指数和组成及分布。Venn图显示了不同供试样本中特异和共有OTU数量差异。结果显示(图3),A层OTU数量排序为:STA(6411) >DTA(6336) >DZA(5983) >SZA(5681);B层OTU数量排序为:DTB(6654) >SZB(6268)>DZB(5977) >STB(5565)。尽管生物结皮A层OTU数量低于B层,但共有OTU数量高于B层。工矿区生物结皮A、B层的特异OTU数量均低于封育区,苔藓结皮的OTU数量高于藻结皮。说明生物结皮通过富集大气降尘重金属能够影响生物结皮A、B层土壤细菌群落结构,并存在种间差异。

图3 生物结皮A、B层土壤细菌群落结构的主坐标分析Fig.3 Principal coordinate analysis (PCoA) of bacterial community structure of A and B horizons of biocrusts
α多样性表征微生物群落的丰富度和多样性,Chao1指数和ACE指数表征物种丰富度,Shannon指数和Simpson指数表征菌群的多样性。测序覆盖率用来评估测序深度。由表5可知,所有供试样本测序覆盖率均 >0.97,表明细菌群落置信度高,测序数据结果可靠。两个样地生物结皮A层的Chao1、ACE、Shannon和Simpson指数均低于B层土壤,表明A层细菌丰富度低于B层。工矿区2类生物结皮A、B层的Shannon指数和Simpson指数均大于封育区;工矿区藻结皮A、B层的Chao1和ACE指数亦大于封育区,但苔藓结皮A、B层的Chao1和ACE指数小于封育区。
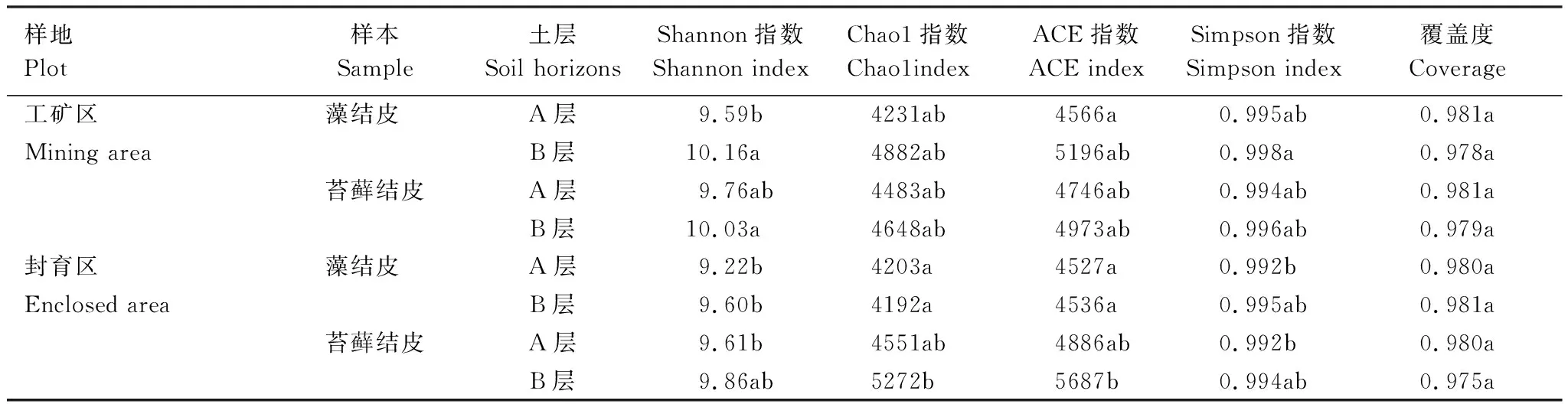
表5 生物结皮A、B层土壤细菌群落α多样性Table 5 Bacterial community α diversity in A and B horizons of biocrusts
2.4 生物结皮A、B层土壤细菌群落β多样性
为了进一步了解不同程度重金属污染程度下细菌群落结构的差异,通过基于Bray-Curtis距离矩阵的主坐标分析(PCoA)可以将两个样地间、结皮类型及层下土壤间的细菌群落相似度和差异性通过点与点间的距离体现出来。如图3所示,在不同生境(工矿区与封育区)或不同土层间(A、B层),生物结皮样点间的距离较远,表明不同样地和不同土层的细菌群落组成均存在明显差异;而在同一生境中,不论A层还是B层,2类生物结皮样点间的距离均较近,表明处于相同样地、相同土层的藻、藓结皮细菌群落结构相似,且同类结皮土壤细菌群落相似。由此推断,人为活动(如重金属污染引起土壤环境的变化)是细菌群落结构变异的主要驱动因素。PCoA1和PCoA2共同解释了40.99%的变异,其中,PCoA1解释了总变异的24.88%,而PCoA2解释了总变异的16.11%。
2.5 生物结皮A、B层土壤的细菌群落组成
生物结皮A、B层土壤中门水平细菌相对丰度结果显示(图4):放线菌门(Actinobacteria,16.64%-40.75%)、变形菌门(Proteobacteria,15.30%-26.03%)和酸杆菌门(Acidobacteria,8.49%-25.29%)为供试样本中的优势菌门,相对丰度最高。其他主要细菌有芽单胞菌门(Gemmatimonadetes,4.49%-11.45%)、绿弯菌门(Chloroflexi,3.71%-10.87%)、浮霉菌门(Planctomycetes,2.32%-7.91%)、拟杆菌门(Bacteroidetes,0.98%-7.48%)、厚壁菌门(Firmicutes,0.32%-3.98%)、奇古菌门(Thaumarchaeota,0.96%-2.81%)、疣微菌门(Verrucomicrobia,0.87%-1.93%)、装甲菌门(Armatimonadetes,0.77%-1.23%)、棒状杆菌门(Rokubacteria,0.27%-0.65%)、硝化螺菌门(Nitrospirae,0.16%-0.36%)、蓝细菌门(Cyanobacteria,0.02%-26.48%)、迷踪菌门(Elusimicrobia,0.02%-0.07%)。其中蓝细菌门为藻结皮的特有种,相对丰度显著高于苔藓结皮(P<0.01)。工矿区中的放线菌门、绿弯菌门和装甲菌门相对丰度大于封育区,厚壁菌门和迷踪菌门相对丰度小于封育区。就土层来看,A层的变形菌门、芽单胞菌门、绿弯菌门、厚壁菌门和硝化螺菌门相对丰度小于B层,而拟杆菌门和蓝细菌门相对丰度大于B层。

图4 生物结皮A、B层土壤中门水平优势细菌相对丰度Fig.4 Relative abundance of the bacterial phylum in A and B horizons of biocrustsAci: 酸杆菌门Acidobacteria; Act: 放线菌门 Actinobacteria; Pro: 变形菌门 Proteobacteria; Bac: 拟杆菌门 Bacteroidetes; Gem: 芽单胞菌门 Gemmatimonadetes; Chl: 绿弯菌门 Chloroflexi; Fir: 厚壁菌门 Firmicutes; Unc: 未分类unclassified; Pla: 浮霉菌门 Planctomycetes; Tha: 奇古菌门Thaumarchaeota; Ver: 疣微菌门 Verrucomicrobia; Arm: 装甲菌门 Armatimonadetes; Rok: 棒状杆菌门 Rokubacteria; Nit: 硝化螺菌门 Nitrospirae; Cya: 蓝细菌门 Cyanobacteria; Elu: 迷踪菌门 Elusimicrobia
2.6 土壤环境因子与生物结皮A、B层细菌群落的关系
对2个样地的优势细菌门与土壤环境因子进行Spearman相关性分析,旨在厘清不同重金属污染水平下生物结皮A、B层细菌群落多样性及优势细菌门相对丰度产生显著差异的原因。结果表明(图5):α多样性(Shannon指数、Chao1指数、ACE指数和Simpson指数)与Hg、As、Cr、pH呈正相关关系,与Cd、EC、SOC、TN呈负相关关系,但只有Shannon指数与Hg达到显著差异(P<0.05)。
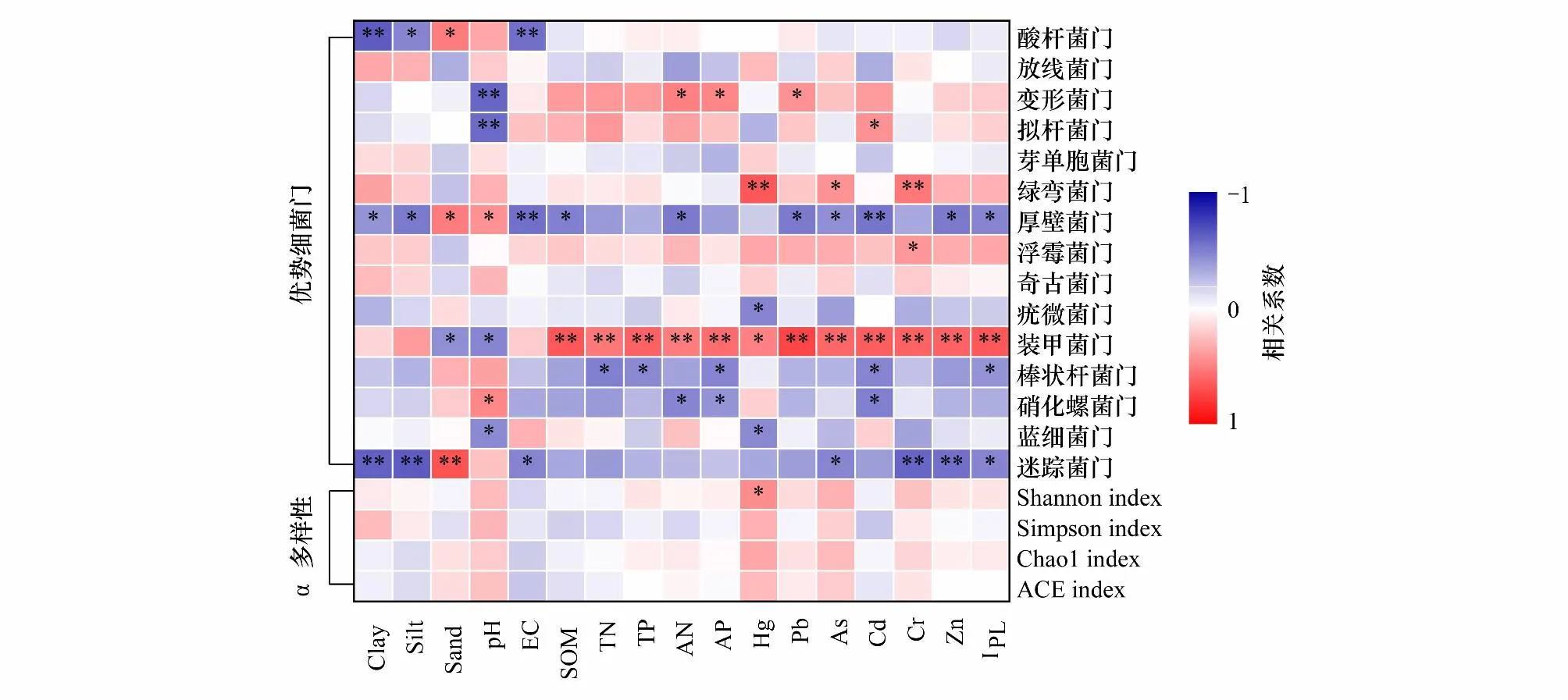
图5 生物结皮A、B层中优势细菌门相对丰度、α多样性与土壤环境因子的相关性Fig.5 Correlation of relative abundance and α diversity of dominant phyla in biocrust and subsoil with soil environmental factorsClay: 黏粒; Silt: 粉粒; Sand: 砂粒; EC: 电导率Electrical conductivity; SOM: 有机质Soil organic matter; TN: 全氮Total nitrogen; TP: 全磷Total phosphorus; AN: 速效氮Available nitrogen; AP: 速效磷Available phosphorus; IPL: 污染负荷指数 Pollution Load Index;*P<0.05;**P<0.01
在优势细菌门中,酸杆菌门与Clay相关性最强(图5,P<0.01);迷踪菌门与Sand相关性最强(P<0.01);变形菌门、拟杆菌门与pH相关性最强(P<0.01);厚壁菌门与EC相关性最强(P<0.01);芽单胞菌门与AP相关性最强;绿弯菌门(P<0.01)、疣微菌门、蓝细菌门与Hg相关性最强(P<0.05);装甲菌门与Pb相关性最强(P<0.01);棒状杆菌门、硝化螺菌门与Cd相关性最强(P<0.05);浮霉菌门与Cr相关性最强(P<0.05)。
为进一步厘清土壤理化性质变化对生物结皮及下层土壤细菌群落的影响,利用Canoco 5.0软件进行了门水平优势细菌与土壤环境因子的RDA分析。结果显示,对细菌群落变异贡献率前5的土壤因子有As(14.3%)> Clay(11.4%)> pH(9.7%)> Pb(8.3%)> Zn(7.7%),说明As、Pb、Zn是影响当地细菌群落结构的重金属元素,而黏粒含量和pH是影响当地细菌群落结构的关键土壤理化因子(表6,P<0.05)。

表6 冗余分析(RDA)中各因子的显著性检验结果Table 6 Significance of factors in redundancy analysis (RDA)
3 讨论
3.1 生物结皮基本理化性质及其对细菌群落结构的影响
大气降尘作为荒漠区成土母质的来源,其组成、通量会影响生物结皮的成土过程、方向和污染程度。本研究中,生物结皮A层具有明显富集大气降尘细颗粒物的能力,并随着生物结皮演替方向呈逐渐增强趋势,促进了生物结皮A层机械组成的细化和养分含量的增加,尤其是工矿区表现尤为突出。这是由于工矿区位于宁夏典型工矿企业富集区,氮沉降较高[17],大气降尘来源复杂,在富含重金属颗粒物的同时,还具有较高的养分和有机质含量[18],而封育区大气降尘主要以沙尘为主,粒径较粗[19]。生物结皮A层的pH值均略低于B层,可能是A层中的微生物能分泌酸性物质所致[20]。生物结皮具有显著改变土壤pH、土壤养分含量及有机碳含量的功能[21-22]。其中,藻结皮具有显著的固氮能力[23],而苔藓结皮具有更好的固碳作用[24]。本研究中,生物结皮捕获近地表风沙流、大气降尘和凋落物,促进了各类生物结皮A、B层土壤养分含量的提高,其中以藓结皮的氮和磷增加幅度最为显著(表6),这主要是由于苔藓结皮具有更高的生物量,且其凋落物是土壤氮和磷的重要来源[25-26]。
已有研究表明,细菌群落结构、多样性会受到外界因素如植被类型、土壤理化性质等的影响,并可以作为衡量土壤生态系统变化的敏感指标[27-28]。生物结皮具有富集及积累大气降尘重金属污染物的作用[2]。本研究中,生物结皮对大气降尘重金属元素具有表层富集效应,表现为A层污染程度高于B层。重金属污染会显著影响土壤细菌群落结构[29]。工矿区生物结皮A层重金属元素Hg、As、Cd、含量超过宁夏土壤背景值。特别是苔藓结皮A层污染负荷指数已达重度污染等级。由于细菌群落对重金属污染敏感,其组成、多样性和相对丰度会对重金属污染做出响应。本研究中,重金属污染水平更高的工矿区生物结皮A、B层的特异OTU数量均低于封育区(图2)。富集重金属能力较弱的藻结皮A、B层的细菌丰富度指数(Chao1和ACE指数)高于封育区,富集重金属能力较强的苔藓结皮A、B层细菌丰富度指数低于封育区,但所有供试样本中均表现为B层土壤细菌多样性和丰富度更高(表5)。这和唐凯[30]在浑善达克沙地生物结皮细菌多样性和群落结构的研究结论一致。可能是由于结皮层富集的重金属长期赋存于在土壤中,部分细菌对重金属产生抗性,造成一些对重金属毒性耐受性强的细菌增殖,相对丰度增大,并使一些耐受性差的细菌相对丰度减少,这和邓琳静[31]和Li等[32]关于重金属胁迫下土壤微生物群落结构和多样性变化规律一致。
植物物种对细菌群落具有重要影响[33]。基于Bray-Curtis距离矩阵的主坐标分析结果表明,同一生境的不同种生物结皮细菌群落结构组成相似,在图中聚类比较明显,但A层和B层的细菌群落离散程度较高,说明富集重金属更多的表层更易于发生细菌群落组成变异(图3)。本研究中,两个样地共存于同一类型生态系统,且无明显的地理隔离。可以推测细菌群落β多样性分布可能是由土壤重金属含量的空间异质性,使细菌群落结构发生分化。
土壤性状显著影响细菌群落结构[20],且这种影响随着重金属污染程度、重金属赋存状态[34]、地上植被类型和土壤基本理化性质(pH、有机碳、机械组成等)发生变化。而土壤性状与生物结皮类型密切相关。生物结皮能够固定碳、氮并活化磷,使表层碳、氮、磷含量均显著增加。本研究中,藓类植物作为生物结皮演替的高级阶段,相较于藻结皮,藓结皮能够显著增加A、B层有机碳和全氮、全磷等养分含量(表2),并提高表层土壤细菌α多样性(表5)。由于生物结皮A层具有滞蓄养分和减弱渗透的作用,使B层养分含量相对较低,但重金属污染也更易富集在A层,避免了B层细菌群落受到更强的重金属污染胁迫,因而表现为B层细菌群落多样性和相对丰度均高于A层。
土壤理化性质(如土壤质地、有机质和pH等)是决定重金属生物有效性的主要影响因子[35]。其中,pH是决定细菌群落结构和多样性变化的最显著的土壤因子。本研究中,α多样性指标均与pH呈正相关,而部分优势细菌门如拟杆菌门和变形菌门相对丰度则与之极显著负相关(P<0.01),可能是由于pH激发了重金属的生物毒性,造成部分细菌门相对丰度减少。放线菌门是土壤团聚体的重要组成部分,与团聚体的稳定性关系密切。本研究中,放线菌门相对丰度与黏粒、粉粒呈正相关关系,而与砂粒呈负相关。可能与生物结皮层具有明显富集细颗粒物的能力有关[36]。由于工矿区生物结皮富集大量富含养分和有机质的大气降尘,可为细菌群落提供更多的可用生态位,表现为试验区生物结皮具有更高的α多样性。
3.2 生物结皮细菌群落对重金属污染的响应
测序结果表明变形菌门、放线菌门、酸杆菌门为2个样地内生物结皮A、B层门水平上的优势细菌群落,这与荒漠区的研究结果相类似[37-38]。封育区和工矿区2类生物结皮的门水平优势菌群相似,但相对丰度存在差异,可能是重金属元素的直接干扰,亦或是重金属的入侵改变了生物结皮的物种组成和土壤性状。其中,放线菌门相对丰度最高且工矿区高于封育区,主要是由于放线菌门对贫瘠环境具有较高的耐受性和适应性[39],此外,放线菌门对金属毒性比较敏感,对重Cd污染耐受性强。研究区Cd严重超标,刺激了放线菌门的金属抗性[40]。其次为变形菌门,作为一种可在高污染水平环境中存活的重金属耐性细菌,其多样化的能量利用途径,有利于变形菌门在矿区广泛存在[41]。酸杆菌门适生于碳源贫瘠的寡营养生存环境,对高重金属浓度有强大的抵抗力并作为矿区的优势细菌门存在[42]。研究表明土壤的非生物性质是土壤细菌群落结构和组成的主要调节因子,如pH、电导率和有机碳等会影响土壤重金属的赋存状态和生物学毒性,从而使细菌群落发生变化[43]。而本研究中,2种生境中的2类生物结皮的A、B层土壤理化因子存在显著差异,受pH、电导率和有机碳的影响表现为重金属污染程度更高的A层细菌群落α多样性均低于B层,工矿区α多样性高于封育区。但分属于不同生境的同类结皮各层次细菌群落相似,这与高天鹏等[35]的研究结论相似。
细菌群落对重金属污染的响应因重金属种类而异。本研究中,工矿区的重金属污染属复合污染。而细菌对重金属污染的耐受性会因土壤环境因子、重金属类别、赋存状态等而异。优势细菌门相对丰度变化对各类重金属元素污染及综合污染的响应既有共性,也有不同。具体表现为绿弯菌门、厚壁菌门、装甲菌门、迷踪菌门是受重金属影响较大的微生物群落,与本研究涉及的重金属相关性较为显著,而其他优势细菌门相对丰度与各类重金属含量并无一致性相关关系,且相关性大多不显著。猜测这可能取决于有效态重金属的浓度和土壤本身的理化性质[44]。除重金属外,理化性质与优势细菌群落之间也存在相关性。机械组成对酸杆菌门、厚壁菌门、迷踪菌门影响较大,有机碳和养分显著影响装甲菌群落。冗余分析结果表明,重金属As、Pb、土壤黏粒含量和pH是影响当地细菌群落结构的主要土壤理化因子(P<0.05)。说明生物结皮在改善土壤理化性质的同时,会通过影响一些重金属的赋存状态和生物学毒性而对细菌群落结构和多样性产生影响。
生物结皮各层次土壤细菌群落结构和多样性可以反映其受重金属污染的程度,鉴于细菌群落种类众多且部分细菌对重金属具有耐受性和特异性,其相对丰度和多样性指标会因重金属污染程度发生相应变化。细菌群落多样性和丰度的变化既体现了细菌群落对重金属污染的适应与响应,也反映了土壤细菌群落在对重金属污染物的生物转化中随着污染物的浓度和种类发生的适应性进化。生物结皮是富集大气降尘重金属、修复污染土壤的潜在理想材料,而利用微生物活细胞吸附重金属是未来生物吸附领域的研究方向[45],因地制宜培育生物结皮中具有重金属抗性的细菌群落则可为工矿区的重金属污染土壤的生态修复提供天然原材料。
4 结论
本研究涉及的两类生物结皮分处于不同生境,大气降尘来源、组成的差异使生物结皮土壤理化性质差异显著。生物结皮对源自大气降尘的重金属元素均具有显著的表层富集作用。由于不同类型生物结皮富集重金属污染的能力不同,不同生物结皮各层次土壤重金属污染程度存在显著差异。变形菌门、放线菌门、酸杆菌门是研究区域的优势门类细菌,但受土壤理化性质和重金属污染程度的影响,不同生境优势细菌门相对丰度存在差异。相关性分析和冗余分析结果表明,重金属As、Pb、土壤黏粒含量和pH是影响当地细菌群落结构的主要驱动因子。
