升温、光周期和氮添加变化对兴安落叶松幼苗叶黄期的影响
2024-01-25张宇钦周广胜吕晓敏武炳义
张宇钦,周广胜,吕晓敏,武炳义
1 复旦大学大气与海洋科学系,上海 200048 2 中国气象科学研究院,北京 100081 3 中国气象科学研究院郑州大学生态气象联合实验室,郑州 450001 4 河北固城农业气象国家野外科学观测研究站,保定 072656
物候是生物长期适应自然环境出现的以年为周期的周期性生长发育节律[1],反映了植物的季节性生长周期。植物物候包括发芽、展叶、开花、叶变色和落叶期等,其变化直接影响到植物的生存繁衍和生物多样性维持[1],进而影响陆地生态系统的生产力、碳平衡和水文循环。政府间气候变化专门委员会(IPCC)第五次评估报告指出,气候系统的变暖是明确无疑的[2]。全球气候变化背景下,物候研究不仅能够更好地理解植被对气候变化响应的过程,而且能够提高大气与植被之间物质与能量交换、陆地生态系统的生产力与碳收支的模拟准确性。正因为如此,物候的研究成为气候学家和生态学家共同关注的热点[3-4]。
植物物候受多生态因子共同影响,分为直接影响和间接影响,其中温度是植物物候的直接影响因子,其他生态因子对物候的影响也是以主导因子-温度的节律性变化为实质改变温度得以实现的[5]。温度升高可提高植物光合酶的活性,促进植物的光合作用,从而加快植物的物候进程[2,6]。另外,植物物候的温度敏感性也受光周期调控。光周期可以通过调整木本植物物候积温需求和冷激需求的非线性关系,进而调整其物候的发生时间,以达到最优的生长策略[7]。最近研究发现,植物叶片的温度敏感性在过去几十年间显著降低[8],这可能与光周期有关[9-10]。同时,氮素是陆地生态系统重要的营养元素,在植物的生长发育和新陈代谢中起着重要作用[11],土壤氮含量的增加不仅影响植物物候,也影响生态系统的生产力和碳平衡。
物候在碳循环、水循环等生态过程中扮演重要角色,特别是秋季物候期,对生长周期的影响比春季物候期更大[12]。然而,现有物候研究侧重于植物春季物候,对秋季物候研究相对较少。研究表明,温度是影响植物春季物候的主要因子,全球变暖背景下气温升高将导致温带和亚寒带植物的春季物候期明显提前。春季1℃的升温会使植物春季的展叶和开花时间提前2.5-6.7d[13]。在秋季物候中,温度同样起着重要的作用,但是影响秋季物候的因子更复杂,温度与其它因子交互作用控制秋季物候[18-20],且在不同物种和实验条件下影响秋季物候的主导因子差异较大[14-17]。研究发现,9月平均气温每升高1℃,兴安落叶松完全变色期推迟4.7d[14];秋季降水量每增加1mm,黄土高原苹果叶变色末期将提前0.07-0.40d,落叶末期将推迟0.12-0.55d[16];也有研究发现,日照增加有利于叶黄素的生成,促进叶色变黄,使叶黄期提前[17]。
另外,现有研究对植物物候响应气候单因子或双因子变化的研究较多[21-24],并已经建立了一些叶片枯黄或凋落的经验模型[25-27]。但是,关于多环境因子交互作用影响物候的研究尚不充分。因此,迫切需要开展多环境因子交互作用影响植物秋季物候的研究,以增进气候变化对植物物候影响的认知,并为物候模型的建立提供依据。
东北地区以兴安落叶松为建群种、集中分布在寒温带针叶林区北端的地带性植被对气候变化较为敏感,且兴安落叶松作为欧亚北方森林的优势树种,在全球变化和区域碳平衡研究中具有重要指示作用[23,28-29]。但是,目前针对兴安落叶松的物候期及其对气候变化响应的研究仍较少。为此,本研究试图以兴安落叶松幼苗叶黄期为研究对象,在河北固城农业气象国家野外科学观测研究站大型人工气候室采用控制实验方法研究兴安落叶松幼苗叶黄期对升温、光周期和氮添加变化及其交互作用的响应,以增进环境变化对兴安落叶松叶黄期影响的理解,助推物候模型的发展,为准确估算生态系统固碳提供依据。
1 材料和方法
1.1 实验设计
实验所选兴安落叶松幼苗产自黑龙江省齐齐哈尔市拜泉县林场,树龄为3年。将生长良好、发育基本一致的幼苗移栽至直径和高度均为20cm的塑料盆中。每盆1株,所用土壤为当地黑土。2019年3月中旬对产地的兴安落叶松幼苗进行装土,在大型人工气候室适应15d后开展模拟实验。实验于2019年4月至11月在河北固城农业气象国家野外科学观测研究站(39°08′N,115°40′E,海拔高度15.2m)开展。
实验共设3个光周期处理(L3短日照10h,L2对照,L1长日照18h)、3个温度处理(T1对照、T2 +1.5℃、T3 +2.0℃)和4个氮添加处理(尿素施用量分别为+0.0%(N1对照0g N m-2a-1)、+50.0%(N2低氮5g N m-2a-1)、+100.0%(N3中氮10g N m-2a-1)、+200.0%(N4高氮20g N m-2a-1)),共36个处理,每个处理4个重复,共144盆。其中,每月对照温度为兴安落叶松幼苗生长地近30年该月的平均气温(表1),增温处理是在对照温度基础上分别升温1.5℃和2.0℃;对照光照14h为兴安落叶松生长地近30年生长季的每日平均日长,光周期处理是在对照光周期基础上分别增加4h和减少4h;对照氮添加量为0,氮添加处理是在对照氮添加量基础上分别增加5g N m-2a-1、10g N m-2a-1、20g N m-2a-1。每月空气相对湿度为兴安落叶松生长地近30年该月的平均相对湿度,CO2浓度为(400±20)μmol/mol,每3天浇水一次,每月灌溉量依据生长季近30年该月的平均降水量确定,确保幼苗存活。每个环境因子均在每月初进行调整,从上一个月的平均值调整为新一个月的平均值。

表1 人工气候室的温度、湿度和灌溉量Table 1 Settings of temperature,humidity,irrigation amount in the artificial climate chamber
1.2 物候观测方法
按照《中国物候观测方法》[30]进行人工观测并记录兴安落叶松幼苗达到各物候的日期,将记录的日期转化为序日(Julian days),观测的物候期包括展叶始期、展叶盛期、叶黄始期、叶黄普期和完全变色期。当观测幼苗伸出的小叶开始分开时,认为其到达展叶始期;当新叶长出的长度达到老针叶一半的时候记录为展叶盛期。当针叶开始明显变黄时认为其到达叶黄始期;叶黄普期和完全变色期分别以50%叶子变色和所有的叶子完全变色作为判断标准。对物候变化较快的阶段(如叶黄普期-完全变色期)每天上午和下午各观测一次,物候变化较慢的阶段(如展叶始期-展叶盛期)每两天观测一次。
1.3 数据处理
使用Microsoft Excel软件对实验数据进行预处理,使用DPS数据分析软件和IBM SPSS Statistics 24软件对相关实验数据进行Pearson相关分析、Spearman相关分析、偏相关分析,登录SPSSAU网站绘制各环境因子不同梯度下兴安落叶松幼苗叶黄各时期平均值及其95%置信区间的条形图,并用Adobe Acrobat DC软件对条形图进行PDF图片编辑,在Matlab软件上采用多元线性回归法对兴安落叶松幼苗叶黄期各物候期和温度、光周期、氮添加量等环境因子进行回归关系拟合,使用IBM SPSS Statistics 24分析软件分析温度、光周期、氮添加量变化及其交互作用对兴安落叶松幼苗叶黄各时期的影响,并对数据进行三因素方差分析和多重比较。
2 结果与分析
2.1 环境因子对兴安落叶松幼苗叶黄始期的影响
兴安落叶松幼苗的叶黄始期与温度和光周期均呈极显著的负相关关系(表2),且与光周期相关性更强,表明升温和光周期延长均使兴安落叶松幼苗叶黄始期显著提前,兴安落叶松幼苗叶黄始期与氮添加量的相关性不显著。

表2 兴安落叶松幼苗叶黄始期与单环境因子的相关性Table 2 Correlation between leaf yellow initial stage of Larix gmelini seedlings and single environmental factor
根据不同升温、光周期、氮添加水平下兴安落叶松幼苗叶黄始期的平均值及其95%置信区间统计(图1)可知,升温和光周期延长均使兴安落叶松幼苗叶黄始期显著提前,而氮添加量增加对兴安落叶松幼苗叶黄始期影响不显著,与相关性分析得出的结论一致。

图1 不同升温、光周期、氮添加水平下兴安落叶松幼苗叶黄始期的平均值及其95%置信区间Fig.1 Mean values and 95% confidence intervals of leaf yellow initial stage of Larix gmelini seedlings under different levels of temperature rise,photoperiod and nitrogen addition
基于多元线性回归拟合的叶黄始期与温度(T)、光周期(L)、氮添加量(N)的多元回归多项式为Y=-0.294T-0.682L+0.050N,其中除N因子外,其它因子回归系数显著性均小于0.01,模型R2=0.565,F=50.245,P<0.001。在全部120个样本中,除个别数据外,残差均接近于零点,且残差的置信区间均包含零点,说明模型拟合效果较好。温度、光周期和氮添加量的变化可解释叶黄始期变化的56.5%。
该回归关系达到极显著水平,且兴安落叶松幼苗叶黄始期对温度、光周期、氮添加量变化的响应关系与相关分析的结果一致,表明升温和光周期延长均使兴安落叶松幼苗叶黄始期显著提前,且兴安落叶松幼苗叶黄始期与光周期的相关性更强;但兴安落叶松幼苗叶黄始期与氮添加量的相关性不显著。
兴安落叶松幼苗叶黄始期对温度、光周期、氮添加量交互作用的响应极显著(表3)。

表3 兴安落叶松幼苗叶黄始期与多环境因子交互作用的三因素方差分析Table 3 Three-factor analysis of variance between leaf yellow initial stage of Larix gmelini seedlings and interactive environmental factors
光温交互作用对叶黄始期影响极显著(表3)。不同光周期下,升温使叶黄始期的提前程度不同,且在光周期14h时最显著;不同温度下,光周期延长对叶黄始期的提前程度无显著差异,且均达到极显著水平。温氮交互作用对叶黄始期影响极显著。不同氮添加下,升温使叶黄始期的提前程度不同,且在施高氮(20g N m-2a-1)时最显著。不同温度下,氮添加使叶黄始期的推迟程度不同,且在升温1.5℃时最显著。光氮交互作用对叶黄始期影响极显著。不同氮添加下,光周期延长对叶黄始期的提前程度不同,且在施低氮(5g N m-2a-1)或高氮(20g N m-2a-1)时最显著。不同光周期下,氮添加对叶黄始期的推迟程度不同,且在光周期10h时最显著。光温氮交互作用对叶黄始期的影响达到极显著水平。
综上所述,升温、光周期和氮添加双因子交互作用对兴安落叶松幼苗叶黄始期影响极显著且存在极值。升温与光周期延长交互作用使叶黄始期提前,且在光周期14h时最显著。光周期延长和氮添加交互作用使叶黄始期提前,且在施低氮(5g N m-2a-1)或高氮(20g N m-2a-1)、光周期10h时最显著。升温与氮添加交互作用使叶黄始期提前,且在施高氮(20g N m-2a-1)、升温1.5℃时最显著。升温、光周期和氮添加变化交互作用对叶黄始期影响极显著。
2.2 环境因子对兴安落叶松幼苗叶黄普期的影响
兴安落叶松幼苗叶黄普期与温度和光周期均呈极显著的负相关关系(表4),且与光周期相关性更强,表明升温和光周期延长均使兴安落叶松幼苗叶黄普期显著提前。兴安落叶松幼苗叶黄普期与氮添加量呈较弱正相关,表明氮添加使兴安落叶松幼苗叶黄普期不显著推迟。
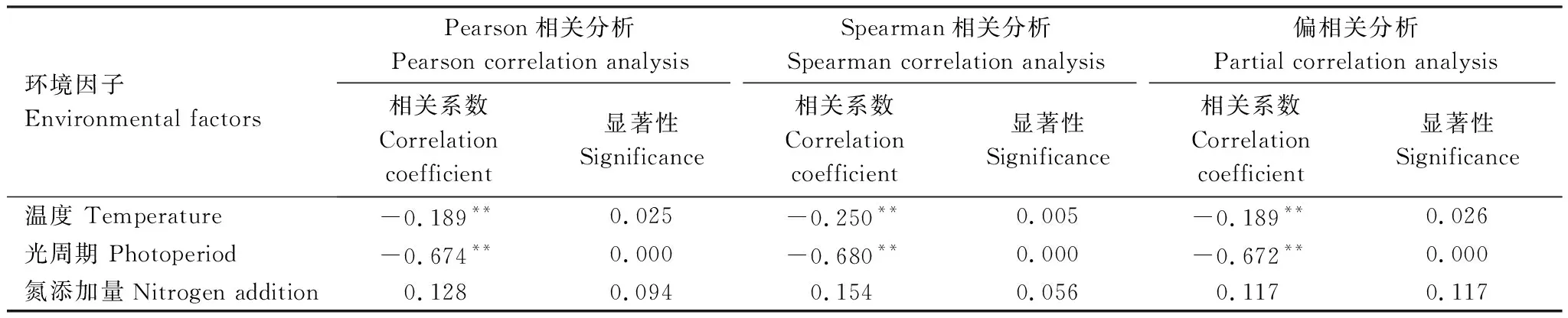
表4 兴安落叶松幼苗叶黄普期与单环境因子的相关性Table 4 Correlation between leaf yellow common stage of Larix gmelini seedlings and single environmental factor
根据不同升温、光周期、氮添加水平下兴安落叶松幼苗叶黄普期的平均值及其95%置信区间统计(图2)可知,升温1.5℃时,兴安落叶松幼苗叶黄普期无明显变化,升温2.0℃时显著提前,光周期延长使兴安落叶松幼苗叶黄普期显著提前,氮添加量增加使兴安落叶松幼苗叶黄普期不显著推迟,与相关性分析得出的结论一致。

图2 不同升温、光周期、氮添加水平下兴安落叶松幼苗叶黄普期的平均值及其95%置信区间Fig.2 Mean values and 95% confidence intervals of leaf yellow common stage of Larix gmelini seedlings under different levels of temperature rise,photoperiod and nitrogen addition
基于多元线性回归拟合的叶黄普期与温度(T)、光周期(L)、氮添加量(N)的多元回归多项式为Y=-0.139T-0.657L+0.085N,其中光周期的回归系数显著性小于0.01,温度的回归系数显著性略小于0.05,模型R2=0.480,F=31.991,P<0.001。在全部108个样本中,除个别数据外,其余数据的残差均接近于零点,且残差的置信区间均包含零点,说明模型拟合效果较好。这表明,温度、光周期和氮添加量变化可解释叶黄普期变化的48.0%。
该回归关系达到极显著水平,且兴安落叶松幼苗叶黄普期对温度、光周期、氮添加量各因子的响应关系与相关分析的结果一致,表明升温和光周期延长均使兴安落叶松幼苗叶黄普期显著提前,且兴安落叶松幼苗叶黄普期与光周期的相关性更强;但氮添加量增加使兴安落叶松幼苗叶黄普期不显著推迟。
兴安落叶松幼苗叶黄普期对温度、光周期、氮添加量交互作用的响应极显著(表5)。

表5 兴安落叶松幼苗叶黄普期与多环境因子交互作用的三因素方差分析Table 5 Three-factor analysis of variance between leaf yellow end stage of Larix gmelini seedlings and interactive environmental factors
光温交互作用对叶黄普期的影响极显著(表5)。不同光周期下,升温对叶黄普期的提前程度不同,且在光周期14h时最显著;不同温度下,光周期延长对叶黄普期的提前程度不同,且在升温1.5℃时最显著。温氮交互作用对叶黄普期影响极显著。不同氮添加下,升温对叶黄普期的提前程度不同,且在施高氮(20g N m-2a-1)时最显著;不同温度下,氮添加对叶黄普期的推迟程度不同,且在升温1.5℃时最显著。光氮交互作用对叶黄普期影响极显著。不同氮添加下,光周期延长对叶黄普期的提前程度不同,且在施低氮(5g N m-2a-1)时最显著;不同光周期下,氮添加对叶黄普期的推迟程度不同,且在光周期10h时最显著。光温氮交互作用对叶黄普期影响极显著。
综上所述,升温、光周期和氮添加变化双因子交互作用对兴安落叶松幼苗叶黄普期影响极显著且存在极值。升温与光周期延长交互作用使叶黄普期提前,且在光周期14h、升温1.5℃时最显著。光周期延长与氮添加交互作用使叶黄普期提前,且在施低氮(5g N m-2a-1)、光周期10h时最显著。升温与氮添加交互作用使叶黄普期提前且在施高氮(20g N m-2a-1)、升温1.5℃时最显著。升温、光周期和氮添加变化交互作用对叶黄普期影响极显著。
2.3 环境因子对兴安落叶松幼苗完全变色期的影响
兴安落叶松幼苗完全变色期与温度呈较弱的正相关关系(表6),升温使兴安落叶松幼苗完全变色期不显著推迟。兴安落叶松幼苗完全变色期与光周期呈极显著的正相关关系,表明光周期延长使兴安落叶松幼苗完全变色期显著推迟。兴安落叶松幼苗完全变色期与氮添加量相关性不显著。

表6 兴安落叶松幼苗完全变色期与单环境因子的相关性Table 6 Correlation between complete discoloration stage of Larix gmelini seedlings and single environmental factor
根据不同升温、光周期、氮添加水平下兴安落叶松幼苗完全变色期的平均值及其95%置信区间统计(图3)可知,升温、氮添加量增加均使兴安落叶松幼苗完全变色期不显著推迟,光周期延长使兴安落叶松幼苗完全变色期显著推迟。

图3 不同升温、光周期、氮添加水平下兴安落叶松幼苗完全变色期的平均值及其95%置信区间Fig.3 Mean values and 95% confidence intervals of complete discoloration stage of Larix gmelini seedlings under different levels of temperature rise,photoperiod and nitrogen addition
基于多元线性回归拟合的完全变色期与温度(T)、光周期(L)、氮添加量(N)的多元回归多项式为Y=0.099T+0.458L+0.009N,其中仅光周期的回归系数显著性小于0.01,模型R2=0.219,F=13.119,P<0.001。在全部144个样本中,除个别数据外,其余数据的残差均接近于零点,且残差的置信区间均包含零点。温度、光周期和氮添加量的变化只能解释完全变色期变化的21.9%。综上所述,升温、氮添加量增加均使兴安落叶松幼苗完全变色期不显著推迟,光周期延长使兴安落叶松幼苗完全变色期显著推迟。
兴安落叶松幼苗完全变色期对光周期变化的响应达极显著水平,对温度与氮添加量交互作用的响应达到极显著水平,对其他因子交互作用的响应不显著(表7)。

表7 兴安落叶松幼苗完全变色期与多环境因子交互作用的三因素方差分析Table 7 Three-factor analysis of variance between leaf yellow complete discoloration stage of Larix gmelini seedlings and interactive environmental factors
光温交互作用对完全变色期影响不显著(表7)。不同光周期下,升温对完全变色期均无显著影响;不同温度下,光周期延长对完全变色期的推迟程度不同,且在不升温时最显著。温氮交互作用对完全变色期影响较显著。不同氮添加下,升温对完全变色期的推迟程度不同,且在施中氮(10g N m-2a-1)时最显著;不同温度下,氮添加对完全变色期均无显著影响。光氮交互作用对完全变色期影响不显著。不同氮添加下,光周期延长对完全变色期的推迟程度不同,且在不添加氮时最显著;不同光周期下,氮添加对完全变色期影响均无显著影响。光温氮交互作用对完全变色期影响不显著。
3 讨论
物候观测已经由标准株的定点人工观测发展到现在利用数字相机法、涡度相关法、遥感光学法等观测[31],并基于观测物候开展了物候与环境因子的关系研究[32]。但是,自然界中的气温、光照、降水、氮沉降等环境因子变化缓慢且幅度微弱[33-34]。本研究采用模拟实验方法,通过人工设置相差幅度较大的多梯度环境因子,使环境因子对植物物候的影响在短时间内更显著[30,35]。
现有物候研究侧重于植物春季物候,且结果相对一致,即温度是影响植物春季物候的主要因子[25],关于秋季物候的研究则相对较少,且不同物种和实验条件下的物候影响因子差异较大[14-17]。研究表明,在秋季物候中,温度同样起着重要作用,但是影响秋季物候的因子更加复杂,温度与其它因子交互作用控制秋季物候[18-20]。在影响兴安落叶松等树种秋季物候的主导因子中,温度或热量因子的影响最显著,光照因子次之[14,22-24]。本研究基于兴安落叶松幼苗叶黄期响应多环境因子的模拟实验表明,光周期对兴安落叶松幼苗叶黄各时期的影响均达到极显著水平,且较温度的影响更显著。究其原因,可能是叶片衰老的开始受光周期调控,升温不会明显推迟叶黄期的开始,但会延缓叶片衰老的速率,使完全变色期推迟[36]。本研究中的光周期设置梯度较大,其中光周期最大达每日18h,最短光周期仅为每日10h,前者较后者多80%,较大的光周期梯度在生长季内维持,使兴安落叶松幼苗叶黄期对光周期的响应被充分展现。
同时,本研究采用多元线性回归方法分析了兴安落叶松幼苗叶黄始期、叶黄普期、完全变色期分别与温度、光周期、氮添加量之间的关系,但拟合模型的解释率并不高,表明叶黄期除受到温度、光周期、氮添加量影响外,其本身的生物学特征甚至其他环境因子可能对兴安落叶松幼苗叶黄期产生重要影响。尽管如此,回归关系的显著性均达到极显著水平,可以解释兴安落叶松幼苗叶黄期对温度、光周期、氮添加量等环境因子的响应,且相关结论与Pearson相关分析、Spearman相关分析、偏相关分析得到的结果具有一致性。
根据交互作用的定义,一个自变量的效果在另一个自变量每一水平上表现不一致的现象,即某一因素的真实效应随着另一因素的改变而改变的现象,称为交互作用[37]。本研究利用三因素方差法和方差分析的多重比较,探究了兴安落叶松幼苗叶黄期对多环境因子交互作用的响应,结果表现为某一个环境因子在不同水平下,另一个环境因子对叶黄始期、叶黄普期的提前(推迟)程度不同,且存在一个水平,提前(推迟)程度最显著,这表明升温、光周期、氮添加变化双因子交互作用对兴安落叶松幼苗叶黄始期、叶黄普期有显著影响并存在极值点,且双因子交互作用对兴安落叶松幼苗叶黄始期、叶黄普期的影响和极值点基本一致。这是对多环境因子交互作用影响物候的补充完善,不仅给出了不同环境因子交互作用对兴安落叶松幼苗叶黄期的影响,而且给出了不同环境因子交互作用影响物候的极值点。另外,三因素方差及多重比较的结论表明,升温、光周期延长和氮添加将延长兴安落叶松幼苗叶黄持续时间。植物叶片变色后,仍然具有一定的同化功能和固碳潜力,只是随叶黄程度加重而不断减弱,到完全变色期时光合能力完全消失[38]。本研究通过叶黄期的研究揭示其变化特征与影响因素,一方面解释物候的变化机制,另一方面为更准确地估算生态系统固碳提供依据。
尽管如此,本研究仅基于兴安落叶松幼苗一年的模拟实验资料,没有考虑前期热量、光能积累等的影响以及植物的生理生态机制[39];另外,本研究的模拟实验采用的是拜泉的三年生幼苗,幼苗和成熟树由于其本身生理条件的不同,在响应环境因子变化上可能出现分化;同时,盆栽到森林存在空间尺度的跨越,控制盆栽研究对于森林生态系统从幼苗到成年树的过程和机制的解析存在不确定性,受控盆栽研究的结果不能直接推广到树木[40]。原位控制实验的研究可以实现从人工实验到推广至树木的效果,然而其成本高,难以实现多环境因素控制[41]。因此,对以兴安落叶松为建群种、集中分布在寒温带的兴安落叶松林研究固然重要,但通过苗木开展物候研究揭示其变化机制有助于增进兴安落叶松林对气候变化响应的理解。国内外研究均提出了盆栽苗研究与实际野外观测结果有一定差异,但仍可以揭示其变化机制,有助于增进植物物候响应多环境因子变化的机理与过程的理解[42]。气候变化对兴安落叶松的影响机制复杂[43],未来仍需开展更多环境因子交互作用、更长时间的模拟实验研究,同时加强原位控制实验研究,以完善兴安落叶松各物候期对气候变化响应的理解[44-45]。
4 结论
本研究基于大型人工气候室的物候对多环境因子响应模拟实验资料,分析研究了升温、光周期和氮添加变化及其交互作用对兴安落叶松幼苗叶黄期的影响。主要结论如下:
(1)升温对兴安落叶松幼苗叶黄期的影响较显著,升温使兴安落叶松幼苗叶黄始期和叶黄普期显著提前,使完全变色期不显著推迟;
(2)光周期变化对兴安落叶松幼苗叶黄期的影响极显著,光周期延长使兴安落叶松幼苗叶黄始期和叶黄普期显著提前,并使完全变色期显著推迟;
(3)氮添加变化对兴安落叶松幼苗叶黄期影响不显著;
(4)升温、光周期和氮添加变化双因子交互作用对叶黄始期和叶黄普期的影响均极显著且均存在极值,但对完全变色期的影响均不显著。升温与光周期延长交互作用使叶黄始期和叶黄普期提前,且在升温1.5℃、光周期14h时最显著。光周期延长与氮添加交互作用使叶黄始期和叶黄普期提前,且在施低氮(5g N m-2a-1)、光周期10h时最显著。升温与氮添加交互作用使叶黄始期和叶黄普期提前,且在施高氮(20g N m-2a-1)、升温1.5℃时最显著;
(5)升温、光周期和氮添加变化交互作用对叶黄始期和叶黄普期影响极显著,但对完全变色期的影响不显著。这表明,升温、光周期延长和氮添加将延长兴安落叶松幼苗叶黄期,延长植物固碳时间。研究结果可为物候模型发展以及森林生态系统碳估算提供依据。
致谢:感谢参与物候模拟实验的周怀林、宋兴阳、周梦子、周莉、刘二华、于鸿莹、顾文杰。
