Food-derived protein hydrolysates and peptides: anxiolytic and antidepressant activities, characteristics, and mechanisms
2024-01-24WenhuiLiYuXiJunruWangYinxiaoZhangHeLiXinqiLiu
Wenhui Li, Yu Xi, Junru Wang, Yinxiao Zhang, He Li, Xinqi Liu
Beijing Advanced Innovation Center for Food Nutrition and Human Health, Beijing Engineering and Technology Research Center of Food Additives,National Soybean Processing Industry Technology Innovation Center, Beijing Technology and Business University, Beijing 100048, China
Keywords: Anxiolytic Antidepressant Peptides Protein hydrolysates Neurotransmitter
ABSTRACT Globally, the prevalence of anxiety and depression has reached epidemic proportions. Food-derived protein hydrolysates and peptides delivered through dietary supplementation can avoid the negative risks associated with traditional pharmaceuticals while delivering superior anxiolytic and antidepressant effects. This review summarizes current research on food-derived anxiolytic and antidepressant protein hydrolysates and peptides,and subsequently analyses their physicochemical characteristics and elaborates on their mechanisms. The aim of this work is to contribute to the in-depth study and provide a theoretical foundation for the development of related products to better serve patients with anxiety and depression.
1. Introduction
Anxiety is an abnormal response to a dangerous or stressful situation that may arise in everyday life, however, the response can develop into an anxiety disorder when it is uncontrollable or happens without the presence of an actual threat[1]. Anxiety disorder is a prevalent mental illness and has been ranked by the World Health Organization (WHO) as the ninth leading health-related cause of disability[2]. There are numerous types of anxiety disorders, all of which cause a wide range of psychological and physiological symptoms, including fear, fatigue, worry or agitation, palpitations,becoming easily frightened and irritability, and others[3-6]. Moreover,anxiety and depression are inextricably linked. Despite having different diagnostic criteria, anxiety and depression are highly comorbid syndromes with a high prevalence and share many symptoms and presentations[7]. Depression is now cited as the leading cause of disability and death worldwide, with almost one in five people suffering a depressive episode at some point in their lives. As an emotional disorder characterized by persistent sadness and/or an inability to experience happiness, depression is also commonly accompanied by defects in daily functioning[8]. It can lead to a loss of capacity to experience pleasure, a sense of worthlessness, fatigue,insomnia, loss of weight or appetite, decreased attention, slow or excited movement, among other symptoms that, in severe cases,significantly increase the risks of suicidality and morbidity[9]. In addition, the currently ongoing global Corona Virus Disease 2019(COVID-19) pandemic has increased the prevalence of anxiety and depression in numerous populations, including children, the elderly,and healthcare workers[10-11].
Psychotherapy and pharmacotherapy are commonly combined in the treatment of anxiety and depression, however, numerous side effects are associated with such drugs, including weight gain, a dry mouth, nausea, and headaches[12]. Consequently, the quest to identify and develop anxiolytic and antidepressant alternatives generated from the natural components in raw foods has become an earnest research topic. Protein hydrolysates, obtained via enzymatic or microbial fermentation, among other methods, generally include nonenzymatic proteins, amino acids, and low molecular weight peptides(3–20 amino acid residues)[13]. Protein hydrolysates and peptides,in contrast to protein, not only maintain the benefits of rich protein nutrition and amino acid balance but also have beneficial physical and chemical qualities such as easy dissolution, absorption, and utilization[14]. Peptides have also been shown to enhance a variety of biological activities, including antioxidant[15], anti-inflammatory[16],antihypertensive[17], hypoglycemic[18], immune regulation[19],and others[20]. Furthermore, in terms of neuromodulation, it has been shown that protein hydrolysates and peptides have a role to play in psychiatric disorders, such as depression, anxiety, and bipolar disorder, as well as in neurodegenerative illnesses, such as cerebral ischemia, Alzheimer’s disease, Parkinson’s disease,among others[21-22].
Thus far, however, there are few articles about the various sources and anxiolytic/antidepressant mechanisms of protein hydrolysates and peptides and incompletely summarized. Therefore, this review investigated and summarized the protein hydrolysates and peptides that have been reported in recent studies to exhibit anxiolytic and/or antidepressant properties. Further analysis of the physicochemical characteristics of peptides can be summarized in general as having a low molecular weight, with ultraviolet absorption (A280nm), being hydrophobic and stable, and containing Leu, Tyr, Pro, and Trp amino acids, with aromatic amino acid leucine or YXXY sequence at N-terminal. In addition, in this paper, the mechanisms of the above mentioned differently sourced anxiolytic and antidepressant protein hydrolysates and peptides are described from the following 4 aspects: In neurotransmitter systems, these protein hydrolysates and peptides regulate the levels of neurotransmitters and act through different neurotransmitter receptors and in a certain order; In terms of neurotrophic functioning, they increase the expression of neurotrophic-related factors and activate the brainderived neurotrophic factor/tyrosine kinase B-extracellular signalregulated kinase/cyclic AMP-responsive-element-binding protein(BDNF/TrkB-ERK/CREB) signalling pathway; In neurons and nerves, they modulate neuronal expression in different regions of the brain, thereby increasing the proliferation of cells and immature neurons while restoring cellular damage; And, in the hypothalamicpituitary-adrenal (HPA) axis, they significantly downregulate the level of corticosterone (CORT), while regulating the expression of corticotropin-releasing hormone (CRH) and adrenocorticotropic hormone (ACTH), thus normalizing the HPA axis. Thereafter,the content of this review is summarized and prospects presented,together with an analysis of the issues involved in current research on anxiolytic and antidepressant protein hydrolysates and peptides. The aim of this work is, thus, to contribute to the discovery, investigation,and more effective application of food-derived antidepressant and anxiolytic protein hydrolysates and peptides, and the expansion of ideas for future research into ways in which these natural resources can assist patients with anxiety and depression.
2. Food-derived anxiolytic protein hydrolysates and peptides
Animal sources, such as milk, fish, eggs, and pig skin, as well as plant sources, including spinach and soybean, are currently used to obtain anxiolytic protein hydrolysates and peptides. Table 1 summarizes the current research on food-derived anxiolytic protein hydrolysates and peptides, among which the research on milk is the most extensive.

Table 1 Food-derived anxiolytic protein hydrolysates and peptides.
2.1 Milk
Milk is the main source of nutrition for mammalian newborns. In addition to the supplementation of essential nutrients, milk performs a variety of functions and biological activities, including calming nerves, inhibiting tumors, and promoting brain development[51].Furthermore, milk also has a certain anxiolytic effect, and studies have shown that milk collected at night (night milk) exerts a greater level of anxiolytic activity than milk collected during the day (day milk)[23]. Night milk treatment at various dosages (100, 200, and 300 mg/kg) all significantly decreased the distance and duration of movement in Institute of Cancer Research (ICR) mice in the open field test, which suggested that night milk is highly rich in Trp and melatonin[52]. In another study, mice that received night milk treatments at the same dose demonstrated an increased percentage of entries and time spent in the open arms in the elevated plus maze test,similarly confirming the anxiolytic effects of night milk[23]. Numerous studies have suggested that the anxiolytic effects of both day milk and night milk may be due to the contribution of rich protein or peptides obtained after digestion.
Milk protein is composed of 80% casein and 20% whey protein[53].Casein comprises four components, namelyαS1,αS2,β, andκ-casein,in whichαS1is the main representative[54-55]. A growing body of published work provides evidence for the anxiolytic activity ofαS1-casein hydrolysates and peptides. Using the conditioned defensive burying test, elevated plus-maze test, passive avoidance test, and conditioned place preference test, researchers have shown thatαS1-casein hydrolysates exert anxiolytic activity in Wistar rats[24-27].A total of three hydrolysates have reportedly been obtained via trypsin hydrolysis[24-25,27]. One anxiolytic peptide,α-casozepine(α-CZP) with the peptide sequence YLGYLEQLLR, corresponds to the 91–100 fragment from bovineαS1-casein, and studies using various neurobehavioral tests, such as the elevated plus-maze test,open field test, conditioned defensive burying test, and the light/dark box test, have demonstrated the anxiolytic activity ofα-CZP in animal models[27-30]. Furthermore, studies have found that the peptides generated by the enzymatic hydrolysis ofα-CZP also exhibit anxiolytic activities[28,30-31]. YLGYLEQ, which corresponds to the 91–97 fragment of bovineαS1-casein, accounted for the greatest proportion inα-CZPin vitrodigestion hydrolysate and was found to significantly improve the neurobehavior of Wistar rats in the elevated plus-maze, open field, conditioned defensive burying, and light/dark box tests[28]. Benoit et al.[30]discovered that anotherα-CZP-derived peptide, YLGYL, which corresponds to the 91–95 fragment from bovineαS1-casein, exhibited anxiolytic properties that resulted in neurobehavioral improvements in Swiss mice when administered via the light/dark box test. In an earlier study, Mizushige et al.[31]observed that YLG, which corresponds to the 91–93 fragment of bovineαS1-casein, exerted a potent anxiolytic-like activity in ddY mice during the elevated plus maze and open field tests.
The anxiolytic effects of whey protein-derived peptides were investigated when Zhu et al.[32]used trypsin, pancreatin, acid protease,and flavourzyme to hydrolyze whey protein isolate, thereby obtaining whey protein hydrolysate (WPH) with a high Trp/five large neutral amino acids ratio, and also synthesizedγ-[Glu]n-Trp (EW) via the enzymatic method. C57BL/6N mice were subjected to chronic restraint stress (CRS, 6 h/day) and then administrated with CORT for 3 weeks to induce anxiety and a depression-like state. Subsequently,the mice were administered WPH for four weeks, during which their percentage of time spent in the center in the open field test did not change but the percentage of entries into the open arms in the elevated plus maze test significantly increased. In addition, EW significantly improved the above two indicators. Zhu et al.[33]further compared the anxiolytic activity of Trp oligopeptides (VAGTWY, VAGTW,SAWY, WYS, and GTW) produced from whey protein and EW.Following identification by ultra-performance liquid chromatographyquadrupole-time of flight-tandem mass spectrometry (UPLC-QTOF-MS/MS), it was found that VAGTWY, VAGTW, and GTW are derived fromα-lactalbumin, whereas SAWY and WYS are derived fromβ-lactoglobulin. The same molding method as that used in the previous study was applied. Their results showed that after 4 weeks of gavage, the WPH and EW had increased the serotonin level, enhanced antioxidant capacity, normalized the HPA axis,and promoted the BDNF-TrkB signaling pathway in the C57BL/6 mice[33]. Aside from the study of the total whey protein, various kinds of research on its constituent proteins have been reported[34-35],such asβ-lactoglobulin, which can produce anxiolytic peptides. The anxiolytic-like activity ofβ-lactotensin (HIRL), a tetrapeptide isolated from a chymotryptic digest of bovineβ-lactoglobulin, was exhibited in ddY mice via the use of the elevated plus maze and open field tests after its intraperitoneal (i.p.) administration[34]. Another study found that thermolysin hydrolysate wheylin-1 (MH) and wheylin-2 (MKG),corresponding to the 145–146 and 7–9 fragments fromβ-lactoglobulin,respectively, showed anxiolytic activity in ddY mice during the elevated plus maze and open field tests. In addition, the anxiolytic activity of wheylin-1 was shown to be more potent than that of wheylin-2[35].
Researchers have, additionally, compared the anxiolytic effects of casein and whey protein in rodent models. In the study of Orosco et al.[36], casein in rat feed was replaced withα-lactalbumin and the rats were fed acutely (30 min) or chronically (3 or 6 days).Compared with their behavior on the casein diet, the Wistar rats spent an increased amount of time in the open arm of the elevated plus maze test and an increased number of visits to the center in the open field test 30 min after being fed with anα-lactalbumin meal. In another study, Vekovischeva et al.[37]replaced casein in mice feed withα-lactalbumin or whey (89.1% (m/m) of the final product in which 23.9% wasα-lactalbumin and 65.2% wasβ-lactoglobulin), and the mice had voluntary access to a running wheel to induce stress.The results showed that the C57BL/6J mice fed theα-lactalbumin meal showed improved neurobehaviors in the light-dark box, elevated plus maze, and open field tests, while those fed the whey diet showed improvement in the social behavioral test.
2.2 Fish
Fish protein and its hydrolysate are also known to provide certain anxiolytic properties. Fish is a plentiful and underutilized source of protein, obtained mostly from the muscle, skin, and waste parts (head,tail, fin, skeleton, viscera, and roe)[56-57]. Studies on the anxiolytic effects of fish protein have focused on fish protein commodities,mainly protein hydrolysates, however, a peptide sequence has yet to be identified.
Orally administered Gabolysat®, a natural fish protein extract product (15 mg/kg at 30 and 60 min), reduced the time spent by Wistar RjHan rats in burying the probe in a conditional defense burial test[26]. Another study investigated the putative effects of Gabolysat PC60 given to Wistar rats for 5 days as a nutritional supplement on their responsiveness to ether inhalation and restraint stressors. According to the findings, the Gabolysat PC60 considerably decreased the release of adrenocorticotropic hormone brought on by ether inhalation and restraint, as well as the amount ofγ-aminobutyric acid (GABA) present in the hippocampus and hypothalamus.Moreover, the PC60 mitigated the increase in noradrenaline and adrenaline induced by the restraint and in adrenaline induced by the ether inhalation[38]. Additionally, the anxiolytic effect of Peptidyss®,a hydrolysate of sardine byproducts (98% of which have a molecular weight below 3 000 Da), was demonstrated in the elevated plus maze and open field tests following its administration to BALB/c mice[39].Currently, however, few anxiolytic investigations of fish peptides have been reported.
2.3 Egg
Egg white is rich in many proteins, including ovalbumin, egg transferrin, egg mucin, and egg globulin[58]. Ovalbumin has thus far been the primary focus of research on egg anxiolytic peptides.The anxiolytic-like activity of the VYLPR peptide, obtained from ovalbumin trypsin digests and corresponding to the 280–284 fragment of ovalbumin, has been demonstrated in the elevated plus-maze and open field tests following its administration to ddY mice[40,59]. In another study, two peptides from egg white alkaline proteinase digests(TNGIIR, RVPSL) also showed anxiolytic effects in spontaneously hypertensive rats (SHR) during the elevated plus maze test[41].
2.4 Pig skin
Pig skin gelatin, the raw material most used in related research,is hydrolyzed from collagen, the most abundant protein in animal tissues and a source of protein in the human diet[60]. In one study,pig skin gelatin was hydrolyzed with a mixture of collagenase I and protease N to obtain two peptides with different molecular weights,one a low molecular weight peptide (LP) ranging from 300–2 000 Da with an average of 367.5 Da, and the other a high molecular weight peptide (HP) of approximately 30 000 Da. After oral administration of the two peptides to C57BL/6 mice for 4 weeks, the results showed that LP significantly reduced the time of mice in the closed arm of the elevated plus maze test compared with HP[42].
2.5 Spinach
D-Ribulose-1,5-diphosphate carboxylase/oxygenase (Rubisco),a crucial enzyme for carbon dioxide fixation and photorespiration,is present in spinach leaves. It is the most prevalent protein on earth and a significant component in green leaves[46]. In an elevated plus maze test, rubiscolin-6 (YPLDLF), a hexapeptide generated from the major subunit of Rubisco, demonstrated anxiolytic effects in ddY mice when administered intraperitoneally (10 mg/kg) and orally(100 mg/kg)[43]. In subsequent studies, Rubisco was digested to obtain a pepsin-pancreatin digestion product, from which Rubimetide(MRW) was extracted in vitro. Rubimetide has been shown to exert anxiolytic-like activity in ddY mice during the elevated plus maze test at a dose of 0.1 mg/kg (i.p.) or 1 mg/kg (oral administration, p.o.)[44-45].The gastrointestinal digestion of Rubisco was also simulated in another study, which conducted a comprehensive peptide analysis of the digests and the structure-activity relationship of known anxiolyticlike peptides, subsequently identified as rALP-1 (SYLPLTT),rALP-1 (1-7) (SYLPLT) and rALP-2 (YHIEPV). Using the elevated plus maze and open field tests, these three peptides were found to exhibit anxiolytic activity in the ddY mice[46].
2.6 Soybean
Soybeans typically comprise approximately 40%–50% protein.Storage proteins, which make up the majority of soybean protein and account for more than 70% of the overall protein, primarily include 7S globulin and 11S globulin (soybean globulin)[61]. At present,the anxiolytic peptides in research are drawn mainly from soybeanβ-conglycinin, which contains three subunits (α,α’, andβ). Three soybean anxiolytic peptides, namely soymorphins-5 (YPFVV),soymorphins-6 (YPFVVN), and soymorphins-7 (YPFVNA),are derived from the soybeanβ-conglycininβsubunit. Humanβ-casomorphin (YPFV) and soymorphins differ in sequence but share the same structure and may both have anxiolytic properties.Soymorphins-5, -6, and -7 all showed anxiolytic activity in ddY mice during the elevated plus maze test[47]. In addition, another soybean anxiolytic, undecapeptide βCGα 323–333 (FLSSTEAQQSY), was obtained when the soybeanβ-conglycininαsubunit was hydrolyzed with trypsin[48]. According to previous studies, the aromatic amino acid leucine dipeptide showed a strong anxiolytic effect, similar to that of diazepam, such as YL, FL, and WL, and the extension of C-terminal Leu also had anxiolytic activity[49-50]. Therefore, FLSSTEAQQSY was selected as the candidate peptide from the trypsin digest, it showed anxiolytic activity in ddY mice during the elevated plus maze and open field tests.
3. Food-derived antidepressant protein hydrolysates and peptides
Animal and plant sources of antidepressant protein hydrolysates and peptides include those from milk, fish, and bovine, as well as those from wheat, soybean, spinach, and mushroom. Table 2 summarizes the current research on food-derived antidepressant protein hydrolysates and peptides.

Table 2 Food-derived antidepressant protein hydrolysates and peptides.
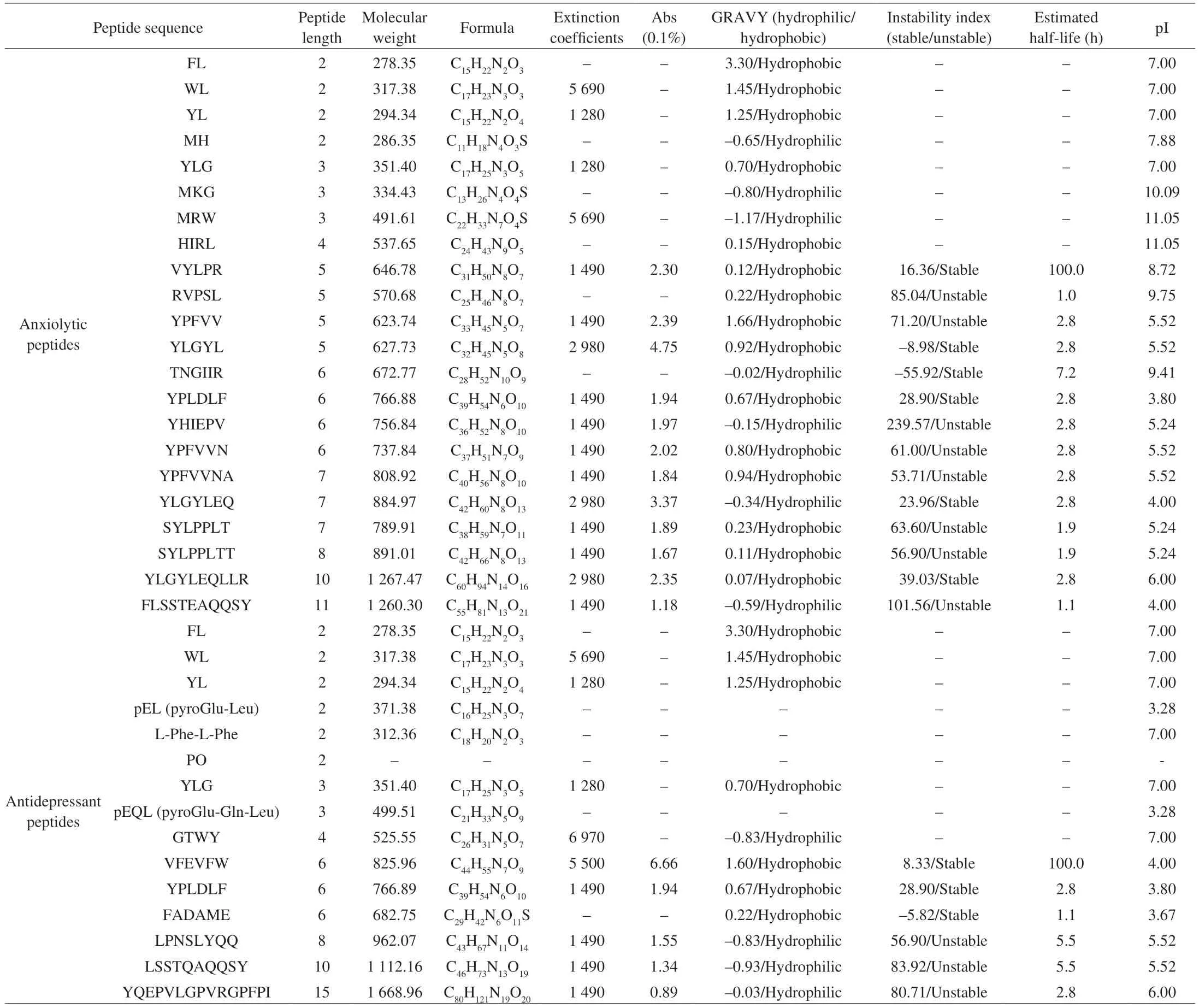
Table 3 Physicochemical characterization of food-derived anxiolytic and antidepressant peptides.
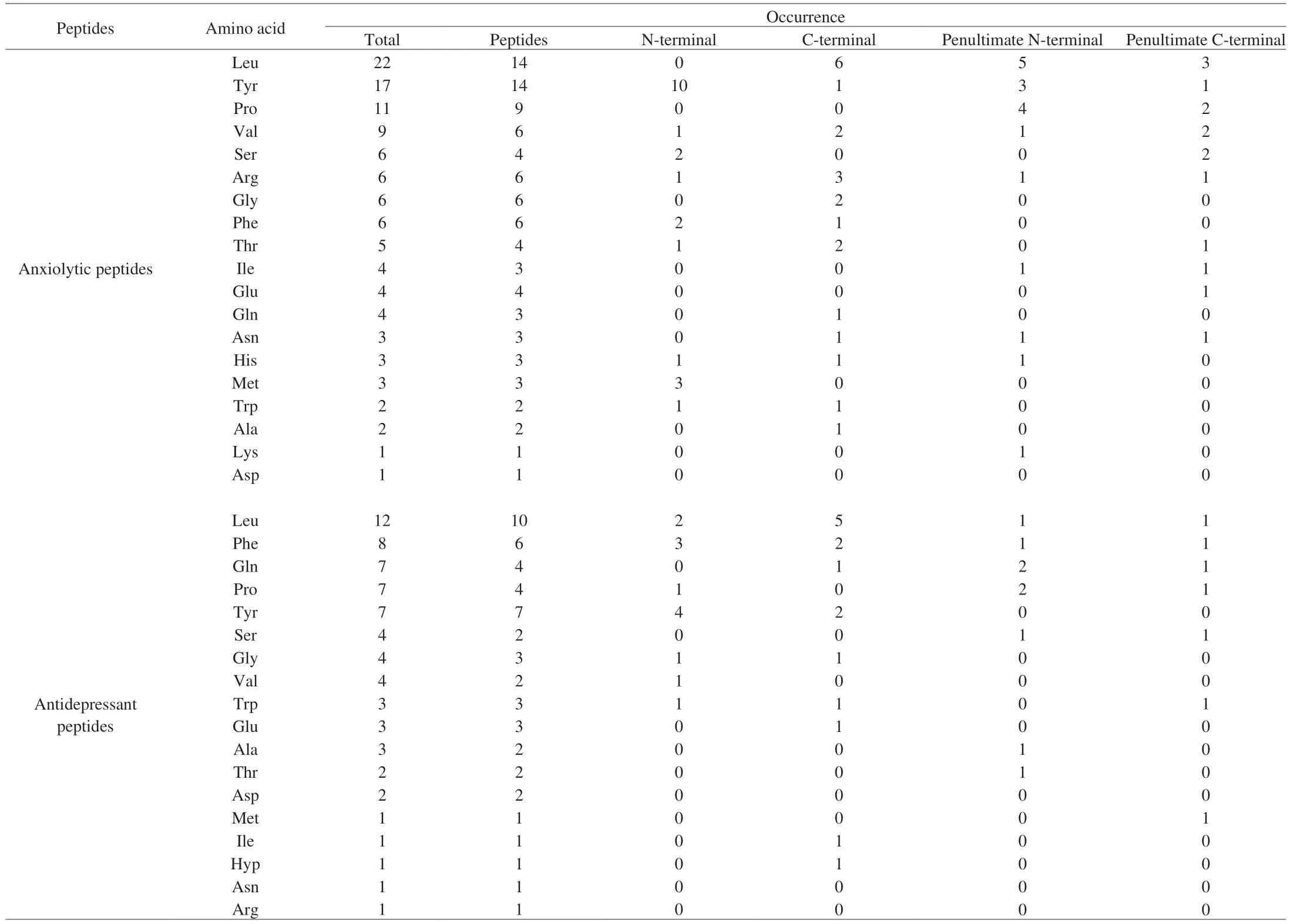
Table 4 Amino acid statistics in anxiolytic and antidepressant peptides.
3.1 Milk
The antidepressant activity of milk-derived peptides has been extensively studied. Aromatic amino acid-leucine dipeptides (YL, FL,and WL) and YLG peptide fragments fromαS1casein showed both anxiolytic and antidepressant activity in a study by Mizushige et al.[62].Following administration of YL, FL, WL (30 mg/kg, i.p.) and YLG(30 or 100 mg/kg, p.o.) to ddY mice, all four peptides significantly reduced their immobility time in the forced swim test. In particular,YL reduced the immobility time of the ddY mice in both the forced swim test (p.o., i.p., and intra-cerebroventricular injection, i.c.v.)and the tail suspension test (i.p.). Qi et al.[63]prepared a functional beverage using bovine milk casein hydrolysates, pepper oil, and Acantho-panax senticosus extract that was found to reduce acute stress after administration (7.50 and 15.00 mg/kg) to Kunming mice by demonstrably shortening their immobility time in the tail suspension and forced swimming tests, respectively. In a chronic unpredictable mild stress (CUMS) model (involving exposure to empty water bottles, tail pinching, overnight illumination, exposure to wet caging, cold water swimming, and tiled caging at 45°), mice reportedly developed various features of depression after 21 days,however, 7.50 and 15.00 mg/kg of the functional drink significantly improved various parameters (including sucrose preference, the number of lattices moved, stand-up times, immobility time, and the number of stool droppings) in the open field and sucrose preference tests. In the anxiolytic study of whey protein, Zhu et al.[32]found that 4 weeks of WPH administration slightly prolonged the immobility times and significantly reduced the total swimming distances of mice in the forced swim test. Another study found thatβ-lactolin (GTWY)obtained via the enzymatic hydrolysis of whey protein demonstrated antidepressant activity when a single oral administration (0.3 and 1.0 mg/kg) reduced the immobility time of ICR mice in the tail suspension test[64].
3.2 Fish
In a similar vein, studies on the antidepressant properties of fish protein have also tended to concentrate on fish protein commodities.Garum Armoricum®(GA), which is prepared via the enzymatic digestion of blue ling fish (Molva dypterygia) protein, has a high percentage of small molecular weight peptides (300–1 500 Da) and contains 30.7% total amino acids, the most important of which are Glu, Gly, Asp, Lys, Ala, Arg, Leu, Pro, Ser, and Ile. Research that observed Wistar Han rats in the conditioned defensive burying and forced swimming tests found that GA decreased the average score of anxiety and the latency of the first approach towards the probe after shock after p.o. dosages of 25, 50, and 100 mg/kg and reduced immobility time after p.o. dosages of 15 and 45 mg/kg[65]. Hairtail(Trichiurus japonicus) also contains antidepressant peptides. In one study, two hydrolysates were obtained using pepsin and pancreatin to simulate gastrointestinal digestionin vitro, denoted as gastrointestinal digestion (GIH) and pepsin digestion (PDH), respectively. The fractions (< 3 kDa) of GIH and PDH were found to have the highest monoamine oxidase A (MAO-A) inhibitory activity[66]. The MAO-A enzyme, located in the outer membrane of mitochondria, is primarily responsible for the metabolism of serotonin and dopamine. Selective inhibition of MAO-A has been shown to increase serotonin levels in the central nervous system and reduce depression symptoms[75].VVFFEW and FADAME peptides were subsequently identified from both the GIH and PDH using liquid chromatography-tandem mass spectrometry (LC-MS/MS). While they both showed the highest MAO-A inhibitory activity in the digested products from their respective sources, that of the VVFFEW peptide was higher and the half-maximal inhibitory concentration (IC50) was 0.405 mmol/L[66].
3.3 Bovine
Collagen hydrolysate with an average molecular weight of 816 Da was reportedly prepared from bovine bone gelatin using a ginger rhizome. The ginger-degraded collagen hydrolysate (GDCH)was administered to ddY mice (30 and 100 mg/kg) and was shown to significantly reduce their immobility time in the forced swim test[67]. In another study, after oral administration of GDCH, prolylhydroxyproline (PO) dipeptides were detected in the cerebrospinal fluid of Wistar-Imamichi rats, indicating that PO dipeptides can be absorbed into the blood from the gastrointestinal tract and transported to the brain. Moreover, PO dipeptides also exhibited antidepressant activity, with oral administration (10 mg/kg) significantly decreasing the immobility time of ddY mice in the forced swim test[68].
3.4 Wheat
Wheat contains approximately 13% protein, of which gluten can be described as the rubber-like substance remaining when the wheat dough is washed to remove starch particles and water-soluble components[76]. Gluten comprises mainly alcohol- and water-soluble glutenins and alcohol- and water-soluble gliadins, and is rich in Glu and Pro[77]. Using the characteristics of the amino acids in gluten,Yamamoto et al.[69]studied the antidepressant effects of wheatderived pyroglutamyl peptides, including pyroglutamyl leucine (pEL)and pyroglutamyl glutaminyl leucine (pEQL). Both pEL and pEQL reduced the immobility time of ddY mice in the tail suspension and forced swim tests, although the activity of pEQL was more effective.
3.5 Soybean
Antidepressant and anxiolytic peptides in soybean are derived fromβ-conglycinin with a similar peptide structure.β-Conglycinin was enzymatically hydrolyzed via thermolysin, subtilisin, and sumizyme, and an antidepressant peptide named soy-deprestatin(LSSTQAQQSY) was subsequently identified from the hydrolysate.The soy-deprestatin exhibited antidepressant-like effects in two different paradigms and two strains of mice when orally administered soy-deprestatin (0.3, 1.0, and 3.0 mg/kg), it decreased the immobility times of ddY mice in the tail suspension and forced swim tests, and similar results were also seen in C57BL/6J mice[70].
3.6 Spinach
The anxiolytic and antidepressant activities of rubiscolin-6 have been proven in numerous studies. Mitsumoto et al.[71]applied acute restraint stress on a group of ICR mice by placing them into a tapered plastic film tube for 2 h. During a subsequent tail suspension test, it was found that while the administration of 30 mg/kg rubiscolin-6 had no effect on non-restraint stressed mice, it significantly reduced the immobility time in the restraint stressed mice.
3.7 Mushroom
The popular culinary and health-promoting mushroom Ganoderma lucidum is used worldwide. As the plant grows, a tiny 4–6 μm ovarian germ cell is known as a Ganoderma lucidum spore (GLS) is released from the body’s folds[78]. Numerous active substances identified in GLS, including polysaccharides, fatty acids, and trace essential elements, provide it with a range of biological activities, including anti-inflammatory[79], anti-tumor[80],and immunity-boosting effects[81]. However, some researchers have also extracted antidepressant peptides from GLS. Zhao et al.[72]used hot water extraction and ethanol precipitation to extract crude polysaccharide peptides from GLS and purified them to obtain a 28 kDa polysaccharide peptide (PGL). After the administration of 200 and 400 mg/kg PGL, the immobility time of C57BL/6 mice in both the tail suspension and forced swimming tests decreased significantly.In addition, CUMS was used to simulate depression in C57BL/6 mice for eight weeks. The stressors included food and water deprivation,exposure to an empty bottle, a soiled cage, light, and darkness in rotation every 2 h, space restriction, a 45° tilt of the cage, overnight illumination, and the sound of predators. Oral administration of PGL was performed for the latter four weeks and the results indicated that PGL dramatically raised the percentage of sucrose preference in the sucrose preference test and decreased immobility time in the forced swimming test.
3.8 Other sources
Antidepressant peptides have also been found in commercially available goods like chicken essence powder and kefir. An antidepressant peptide identified as the diketopiperazines (DKP)loop (L-Phe-L-Phe) was purified from chicken essence.In vitro, by establishing the blood-brain barrier (BBB) model, it was found that DKP can enter the brain through the BBB.In vivo, oral administration of DKP significantly reduced escape latency in the water maze test in depressed C57BL/6N mice that had been previously subjected to repeated open-space swimming tasks[73].
Kefir, a traditional fermented dairy product, is prepared at an artisanal level by adding kefir grains (5%–10%) as a fermenting agent to the whole, semi-skimmed or skimmed and pasteurized milk of goat, sheep, camel, or buffalo[82]. Chen et al.[74]prepared kefir peptides(KFP) with a molecular weight < 3 000 Da and, after administration(150 mg/kg) to CD-1 mice, found that they could reduce the duration of immobility in the forced swimming and tail suspension tests, increase the time spent in the open arm and center zones in the elevated plus maze test, and raise the total distance traveled,average speed and time spent in the central area in the open field test. Subsequently, the KFP was further isolated and KFP-1, -3, -5 were identified. It was found that among the three peptides, KFP-3(YQEPVLGPVRGPFPI) had the best antidepressant effect, with an oral dose of 10 mg/kg to mice significantly reducing their immobility time in the forced swim test and increasing their mobility time in the tail suspension test.
4. Physicochemical characterization of food-derived anxiolytic and antidepressant peptides
The food-derived anxiolytic and antidepressant peptides summarized above were subsequently further analyzed in this study to identify additional peptides with similar activities. As shown in Tables 3 and 4, the analysis included both peptides (peptide length,molecular weight, formula, extinction coefficients, Abs 0.1%,hydrophilic/hydrophobic, stable/unstable, estimated half-life and pI)and amino acids (occurrence and number of total peptides, N-terminal,C-terminal, penultimate N-terminal and penultimate C-terminal).
It was found that the food-derived anxiolytic peptides ranged from dipeptides to decapeptides, with molecular weights of 278.35 to 1 267.47 Da. In addition, 72.7% of the peptides included extinction coefficients and, due to the presence of Trp, Tyr and Cys absorbed light at 280 nm. The ratio of hydrophobic peptides to hydrophilic peptides was 15:7, while that of stable peptides to unstable peptides was 6:8. The estimated half-lives of the peptides were mostly concentrated at 2.8 h or less, with only two peptides (TNGIIR and VYLPR) exhibiting longer half-lives. Leu, Tyr, Pro, and Val were found to be the key amino acids, occurring at frequencies of 22, 17,11, and 9, respectively. These amino acids made up 63.6%, 40.9%,and 27.3% of the total peptides. The N-terminal region was noted to have a higher abundance of different amino acid species, with 45.5% of peptides having Tyr at the ultimate N-terminal position,while the proportion of the remaining amino acids, Met, Phe, and Ser, was small. Leu, Pro, and Tyr made up the majority of the amino acids in the penultimate N-terminal position, with Tyr being the most prevalent. In the ultimate C-terminal, the amino acid species were more dispersed, with Leu (27.3%) and Arg (13.6%) most prevalent.Leu was generally present in the penultimate C-terminal amino acid position. It is worth noting that peptides with aromatic amino acid leucine dipeptides (YL, WL, and FL) exhibited anxiolytic activity and were typically located at the N-terminal of the peptide. And peptides with a YXXY pattern also have anxiolytic activity because the positions of the two Try are similar to the anxiolytic medication(nitrazepam). In addition to individual peptides, tryptophan-rich mixed peptides (VAGTWY, VAGTW, SAWY, WYS, and GTW)could alleviate anxiety by regulating serotonin metabolism.
The food-derived antidepressant peptides varied from dipeptides to pentadecapeptides, with molecular weights of 278.35–1 668.96 Da.Most of the peptides absorbed light at 280 nm (64.3%). The ratio of hydrophobic peptides to hydrophilic peptides was 7:4, while that of stable peptides to unstable peptides was 3:3. The estimated halflives of the peptides were evenly distributed, with VFEVFW found to have a longer half-life (100 h). Leu, Phe, Tyr, Pro, and Gln were the key amino acids in these peptides, occurring 12, 8, 7, and 7 times,respectively, and comprising 66.7%, 40.0%, 46.7%, 26.7%, and 26.7% of the total peptides, respectively. The amino acids found in the N-terminal (Tyr, Phe, and Leu) were more scattered, while in the C-terminal they were more concentrated (Leu, 33.3%). The amino acids were mainly Pro and Gln in the penultimate of the N-terminal,while more species at a lower proportion of each were found in the penultimate of the C-terminal. In addition, the antidepressant peptides contained Hyp (PO), and there were two modified peptides at the N terminus (pEQL and pEL). The above analyses of all the food-derived anxiolytic and antidepressant peptides revealed that they share the following characteristics: low molecular weight, ultraviolet absorption(A280nm), hydrophobic and stable, contains Leu, Tyr, Pro, and Trp amino acids with aromatic amino acid leucine or YXXY sequence at N-terminal. Based on the qualities listed above, more food-derived anxiolytic and antidepressant peptides can be identified in future investigations.
5. Mechanisms of anxiolytic and antidepressant protein hydrolysates and peptides
Based on the comprehensive information reported in related research, the mechanisms of the anxiolytic and antidepressant protein hydrolysates and peptides were categorized into four aspects.As shown in Fig. 1, first, in terms of the neurotransmitter system,protein hydrolysates and peptides can regulate the content of neurotransmitters and act through various neurotransmitter receptors in a certain order to exert their anxiolytic and antidepressant effects;Second, in terms of neurotrophic function, protein hydrolysates and peptides can increase the expression of neurotrophic-related factors and activate the BDNF/TrkB-ERK/CREB signal pathway, so as to regulate the survival, growth and differentiation of neurons and,thereby, alleviate depression and anxiety; Third, in terms of neurons and nerves, protein hydrolysates and peptides can alleviate depression and anxiety by regulating the expression of neurons in different brain regions, increasing proliferating cells and immature neurons,and restoring cell damage; Finally, in terms of the HPA axis, protein hydrolysates and peptides can significantly down-regulate the level of CORT and the expression of CRH and ACTH, so as to regulate the HPA axis to alleviate anxiety and depression.
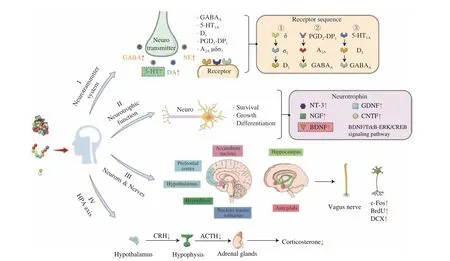
Fig. 1 Mechanisms of anxiolytic and antidepressant protein hydrolysates and peptides.
5.1 Neurotransmitter system
5.1.1 Neurotransmitter
Neurotransmitters are molecules that, along with electrical signals, transmit and amplify signals between nerves, as well as between nerves and other cells[83-84]. Neurotransmitters are excitatory, inhibitory, or both and they play roles in numerous and diverse functions, from mood and movement to sleep. The main types of neurotransmitters include acetylcholine, biogenic amines,amino acids, neuropeptides, purines, dissolved gases, and lipids[84].Anxiolytic and antidepressant protein hydrolysates and peptides act mainly on 5-hydroxytryptamine (5-HT), GABA, dopamine, and norepinephrine.
5.1.1.1 5-HT
5-HT is synthesized from Trp via a short metabolic pathway, and its main functions are feeding, sleeping, dreaming, mood regulation,thermoregulation, and pain transmission[85]. 5-HT is an inhibitory neurotransmitter involved in the control of circuits in anxiety and fear[86]. It has been found that increasing its levels can reduce the symptoms of patients with anxiety disorders[87], and many different substances can inhibit the reuptake of 5-HT, thus alleviating anxiety and depression.
α-Lactalbumin, GLS polysaccharides peptides, DKP cyclopeptides, a functional beverage (including active peptides, menthol,and eleutheroside), and KFP have all been reported to increase the 5-HT levels in mice and rat brains[36,63,72-74]. Zhu et al.[32-33]conducted two studies on the mechanism of EW and WPH in anxious-depressed mice. One study focused on the relationship between 5-HT synthesis and antioxidant indicators, while the other investigated the relationship between 5-HT synthesis and anti-inflammatory factors. In combining,the results of the two studies showed that EW and WPH affect the synthesis and utilization of 5-HT in 5 main ways, thereby alleviating anxiety and depression, as shown in Fig. 2: 1) EW and WPH act as direct sources of Trp to increase the concentration of Trp in the brain;2) EW and WPH can increase 5-HT precursors by inhibiting the kynurenine pathway, including reducing the activity of indoleamine 2,3 dioxygenase (IDO), which is responsible for the conversion of Trp to kynurenine; 3) EW inhibits mitogen-activated protein kinase(MAPK) enzymes, which otherwise reduce the synaptic utilization of 5-HT; 4) EW enhances the utilization of tetrahydrobiopterin (BH4),a coenzyme factor that promotes the conversion of Trp to 5-HT;and 5) EW and WPH indirectly reduce the levels of oxidative stress indicators such as reactive oxygen species (ROS) and nitric oxide synthase (NOS), thereby attenuating the damage of 5-HT neurons by malondialdehyde (MDA).
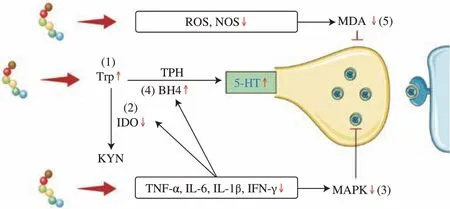
Fig. 2 γ-[Glu]n-Trp and whey protein hydrolysates regulate 5-HT synthesis and utilization by (1) increasing Trp concentration, (2) decreasing IDO activity, (3) inhibiting MAPK enzymes, (4) enhancing BH4 utilization, and(5) attenuating the damage by MDA.
5.1.1.2 Dopamine
Dopamine plays important roles in cognition, learning, mood,and sleep, but its activity as either an excitatory or inhibitory neurotransmitter depends on which receptor subtype is activated.The metabolism of dopamine’s involvement in anxiety regulation is complex, however, studies show that an increase in dopaminergic signals does reduce anxiety and depression to a certain extent[88].
β-Lactolin, PO dipeptides, GLS polysaccharides peptides, DKP cyclo-peptides, a functional beverage (active peptides, menthol, and eleutheroside), and KFP have all been shown to increase dopamine levels in mouse and rat brains[63-64,68,72-74]. One study of PO explored the changes it exerted on dopamine metabolites in the brains of ddY mice and found that, following the oral administration of PO dipeptides, dopamine concentrations in the prefrontal cortexes of the treated mice were higher than those in the control mice, whereas there were no significant changes in the hippocampus. Furthermore, the concentrations of 3,4-dihydroxybenzeneacetic acid, homovanillic acid,and 3-methoxytyramine in the prefrontal cortex and hippocampus were not significantly different[68].
5.1.1.3 Norepinephrine
In the cytoplasm, norepinephrine is produced from Tyr and is closely related to the components of the fear circuit[86]. GLS polysaccharides peptides, DKP cyclo-peptides, a functional beverage(active peptides, menthol, and eleutheroside), and Gabolysat PC60 have all been found to significantly elevate norepinephrine levels in the brains of mice and rats[38,63,72-73]. The study on Gabolysat PC60 examined circulating catecholamines and found that in the absence of stress and ether inhalation, administration of PC60 (300 or 1200 mg/(kg·day)) and diazepam did not significantly alter the plasma norepinephrine or epinephrine in rats. In contrast, treatment of rats with PC60 (1 200 mg/(kg·day)) or diazepam under restraint stress resulted in elevated circulating norepinephrine and epinephrine[38].
5.1.1.4 GABA
GABA, a glutamate-derived inhibitory neurotransmitter, is principally important for promoting sleep and preventing arousal[89].Stress can increase the GABA levels in the hippocampus and hypothalamus of the rat brain. Rats gavaged with glutamate-rich Gabolysat PC60 after ether inhalation or restraint stress displayed a minor decrease in GABA concentrations in the hippocampus but a notable decrease in the hypothalamus[38].
5.1.2 Neurotransmitter receptors
After neurotransmitters have been secreted by synaptic vesicles,they bind to their respective receptors and perform different functions,depending on the binding receptor. Studies on the anxiolytic and antidepressant effects of protein hydrolysates and peptides have shown that neurotransmitter receptors typically use antagonists to bind to the receptor and inhibit it from binding to other substances.Utilizing animal behavior tests, researchers have subsequently tried to determine whether it is through their binding receptors that these protein hydrolysates and peptides exert their anxiolytic/antidepressant effects. Thus far, studies have been reported involving 5-HT receptors, GABA receptors, dopamine receptors, prostaglandin receptors, adenosine receptors, and opioid peptide receptors.
5.1.2.1 5-HT receptors
5-HT receptors are widely distributed in the brain and are located mainly on the cytosol and dendrites of 5-hydroxytryptaminergic neurons in the middle suture nucleus. At least 16 different types of 5-HT receptors have been identified, with 5-HT1Abeing one of the most distinctive[90]. The 5-HT1Areceptor is crucial for neural activity and is closely linked to anxiety and depression. Research using animal behavioral tests has demonstrated that 5-HT1Areceptor agonists alleviate anxiety and depression in rodents[91].
Five anxiolytic peptide studies involving 5-HT receptors all utilized the 5-HT receptor antagonist WAY100135. Comparative analyses of the effects of peptide intervention both with and without WAY100135 have found that YLG, rALP-1, rALP-1 (1–7), βCGα,FL, WL, and YL all increased the percentage of time that rodents spent in the open arm of the elevated plus maze test in the absence of WAY100135, but decreased the time when administered together with WAY100135, indicating that all of those peptides activated 5HT1Areceptors and, thus, exerted anxiolytic activity[31,46,48-50]. Two studies of antidepressant protein hydrolysates and peptides presented different results: YL dipeptide and GDCH were both found to reduce the immobility time of mice in the forced swimming test, however,the antidepressant effect of the YL dipeptide was reversed with WAY100135, while the antidepressant effect of the GDCH remained present. These results suggest that YL dipeptides, but not GDCH, can exert antidepressant effects via 5-HT receptors[62,67].
5.1.2.2 Dopamine receptors
Dopamine receptors are widely present in the central nervous system, as well as in peripheral tissues such as blood vessels, kidneys,and adrenal glands. Typically, dopamine receptors belong to one of two families, namely D1-like (D1and D5) and D2-like (D2, D3,and D4),with D1and D2expressed most strongly in the brain[92].
Six dopamine receptor-related anxiolytic peptide studies used the dopamine D1receptor antagonist SCH23390, some of which also used another dopamine D1receptor, antagonist SKF83566, and D2receptor antagonist raclopride. YLG,β-lactotensin, rubiscolin-6, βCGα, FL,WL, and YL have all been found to increase the percentage of time spent in the open arm of the elevated plus maze test without the antagonists, but to significantly reduced the time when administered together with the antagonist[31,34,43,48-50]. The findings indicate that all these peptides can exert an anxiolytic effect via the activation of the dopamine D1receptor and thatβ-lactotensin and rubiscolin-6 do not act through the dopamine D2receptor[34,43]. The results for antidepressant protein hydrolysates and peptides are reportedly the same as those for anxiolytic peptides. In the absence of SCH23390,YL and GDCH significantly reduced the immobility time of mice in the forced swim test, whileβ-lactolin and whey digest significantly reduced the immobility time of mice in the tail suspension test.In contrast, following the administration of SCH23390 to mice,the abovementioned protein hydrolysates and peptides showed no antidepressant effects. The results suggest that YL, GDCH,β-lactolin,and whey digest exert antidepressant activity through the dopamine D1receptor[62,64,67].
5.1.2.3 GABA receptors
GABA receptors, the primary inhibitory neurotransmitter receptors in the mammalian brain, are present mostly in the central nervous system but can also be found in the liver, lungs, and other cells[93]. They include two main types of receptors, namely GABAAand GABAB, both of which inhibit neuronal activity in the brain by opening the transmembrane channels and making them permeable to chloride that binds to GABA. Should these channels become dysfunctional, anxiety and depression can result[94].
Seven anxiolytic peptide studies involving GABAAreceptors used the GABAAreceptor antagonist bicuculline.α-Casozepine, YLG,ovolin, rubimetide, βCGα, FL, WL, and YL have all been found to increase the percentage of time spent in the open arm in mice utilized in the elevated plus maze test, but to significantly reduce this time when administered with bicuculline. According to the findings,each of the peptides listed above has shown anxiolytic effects via the activation of GABAAreceptors[27,31,40,45,48-50]. Interestingly, one of these studies onα-casozepine found that it had a 10 000 lower affinity for the benzodiazepine site of GABAAthan diazepamin vitro,and was 10 times more effective than diazepamin vivowhen used in the conditioned defense burial test[27]. The antidepressant activity of YL remained consistent with that found in studies of anxiolytic peptides. YL greatly reduced the immobility time in mice in a forced swimming test but, in contrast, could no longer reduce immobility time after the use of bicuculline and flumazenil. The findings indicate that YL activates the benzodiazepine and GABA sites of the GABAAreceptor to produce its antidepressant effects[62]. Flumazenil is an antagonist of the benzodiazepine site, whereas benzodiazepine is an antidepressant[95].
5.1.2.4 Opioid receptors
Opioid receptors are secreted by central and peripheral neurons and nerves (the pituitary and adrenal glands) and are expressed by immune and ectodermal cells. There are three main types of opioid receptors in the central nervous system: μ, δ, and κ[96]. When drugs bind to opioid receptors, they can reduce pain, regulate mood and relieve anxiety symptoms[97].
Studies using the guinea pig ileum and mouse vas deferens tests to investigate the μ opioid receptor reported that soymorphins (YPFVV,YPFVVN, and YPFVVNA) showed a dose-dependent inhibition of guinea pig ileal contractions and higher opioid activity than in the mouse vas deferens test, suggesting that soymorphins-5, -6 and -7 are selective for μ opioid receptors[47]. The δ opioid receptor and opioid receptor antagonists were examined in two anxiolytic peptide studies and one antidepressant peptide study of spinach-derived rubiscolin-6 both used naltrindole, an antagonist of δ receptors. It was found that naltrindole blocked the increase in the percentage of time and number of visits in the open arms induced by rubiscolin-6 in ddY mice in the elevated plus maze test, the increase of time in the elevated plus maze test open arms by ddY mice induced by rALP-2, and the decrease of the immobility time induced by rubiscolin-6 in restraint stressed mice in the tail suspension test. The data, thus, revealed that rubiscolin-6 and rALP-2 exert anxiolytic and antidepressant effects via δ opioid receptors[43,46].
5.1.2.5 Prostaglandin receptors
Prostaglandin (PG) receptors include several classifications,among which PGD2is widely distributed and abundant in the central nervous system and is involved in the regulation of several functions, such as platelet activation, vasodilation inhibition and sleep regulation, thereby improving symptoms of anxiety[98]. Two anxiolytic peptide studies administered the PGD2-PGD receptor1 (PGD2-DP1)receptor antagonist BW A868C intraperitoneally to ddY mice and found that it blocked the anxiolytic-like activity of rubimetide and ovolin, suggesting that the anxiolytic-like activity of both peptides is mediated by the PGD2-DP1receptor[40,44].
5.1.2.6 Adenosine receptors
Adenosine, an important intermediate metabolite, is ubiquitously present in the extracellular mediators of all tissues and acts as a local regulator via the activation of G protein-coupled receptors,also referred to as adenosine receptors. There are four main types of adenosine receptors, namely A1, A2A, A2B,and A3, all of which are extensively expressed and involved in a variety of physiological and pathological functions, including cardiac rhythm and circulation,sleep regulation, and neurodegenerative diseases[99]. Two anxiolytic peptide studies, both involving adenosine A2Areceptors, showed that rubimetide and ovolin increased the time spent in the elevated plus maze test open arms by mice without SCH58261, but this time was significantly reduced with the use of SCH58261. Both rubimetide and ovolin can activate adenosine A2Areceptors, thereby exerting anxiolytic activity[40,44].
5.1.2.7 Sigma receptors
The σ receptor, previously categorized as an opioid receptor but now classed as a separate group, is a relatively new receptor type that is still poorly understood. It is located mostly in the central nervous system and peripheral organs and is separated into two subtypes, σ1and σ2[100]. σ Receptors are closely associated with a variety of neurological disorders and have been linked to anxiety[101].Hirata et al.[43]conducted a study using two σ1receptor antagonists,BMY14802 and BD1047, and found that both blocked the increase of time spent in the elevated plus maze test open arms by ddY mice induced by rubiscolin-6, which suggested that the anxiolytic effect of rubiscolin-6 is mediated via σ1receptors.
5.1.3 Neurotransmitter receptor sequence
The anxiolytic and antidepressant effects of protein hydrolysates and peptides are related not only to the type of neurotransmitter receptor, but also to the order of the neurotransmitter receptor binding.Hirata et al.[43]found that rubiscolin-6 exerts anxiolytic effects through δ opioid receptors, σ1receptors, and dopamine D1receptors.The combined findings of previous studies suggest that the order of rubiscolin-6 activating receptors is: δ opioid receptor→endogenous σ1ligand→σ1receptor→dopamine→D1receptor[43]. Another spinachderived anxiolytic peptide, rubimetide, acts through the PGD2-DP1receptor, adenosine-A2Areceptor, and GABA-GABAAreceptor, and the sequence of rubimetide activated receptors is hypothesized to be: PGD2-DP1receptor→Adenosine-A2Areceptor→GABA-GABAAreceptor[45]. In another study, the YL anxiolytic peptide was found to act through 5-HT1A, D1and GABAAas well as through the use of agonists (5-HT1Areceptor agonist 8-OH-DPAT, D1receptor agonist SKF38393, GABAAreceptor agonist muscimol) and antagonists,and the sequence of YL-activated receptors was detected as 5-HT1Areceptor→D1receptor→GABAAreceptor[50].
5.2 Neurotrophic function
Neurotrophic peptide factors are structurally related and are essential for neuronal survival, growth, and differentiation. Among them, BDNF, nerve growth factor (NGF), neurotrophin-3 (NT-3), and neurotrophin-4/5 (NT-4/5) form the core of the family[102].
Studies of anxiolytic and antidepressant protein hydrolysates and peptides have focused mainly on BDNF, with more research conducted in relation to antidepressants. As an important member of the neurotrophic factor family, BDNF can promote neurogenesis,neuronal survival, and differentiation, while also maintaining the function of mature neurons[102]. Stress significantly reduces BDNF levels in the mice hippocampus and accelerates depression-like symptoms, whereas several studies have shown that different protein hydrolysates and peptides (EW, Trp oligopeptides derived from whey protein, GDCH, GLS polysaccharides peptides, KFP, and PO dipeptides) can increase the expression of BDNF and, thus, alleviate anxiety and depressive symptoms[33,67-68,72,74]. In addition, EW, Trp oligopeptides derived from whey protein and KFP can further promote the BDNF/TrkB-ERK/CREB signaling pathway[33,74]. As shown in Fig. 3, herein of the BDNF/TrkB-ERK/CREB signaling pathway, BDNF binds to its downstream receptor, TrkB, to induce Tyr phosphorylation, then activates the ERK/CREB pathway to transcribe into pro-BDNF mRNA, which undergoes translation to generate pro-BDNF, resulting in BDNF. EW and WPH have been found to increase the mRNA expressions of TrkB and CREB, thus promoting the BDNF/TrkB signaling pathway[33]. Furthermore, KFP activates the BDNF signal by elevating the phosphorylation of TrkB and ERK1/2, and raising the mRNA and protein levels of BDNF to relieve depression-like behavior[74].

Fig. 3 Anxiolytic and antidepressant protein hydrolysates and peptides promote the BDNF/TrkB-ERK/CREB signaling pathway.
In addition, neurotrophic factors NGF and NT-3 are also involved in regulating the proliferation and differentiation of neural stem cells.Two studies have found that GLS polysaccharides peptides and PO dipeptides significantly increase the expression levels of NGF and NT-3 in the brains of ddY mice, thereby alleviating anxiety and depression symptoms[68,72]. Mizushige et al.[67]examined multiple factors with neurotrophic functions. In addition to BDNF, NGF,and NT-3, glial cell-derived neurotrophic factor (GDNF), ciliary neurotrophic factor (CNTF), vascular endothelial growth factor α(VEGFα), fibroblast growth factor 2 (FGF2) and insulin-like growth factor 2 (IGF2) were also included in their study of the antidepressant activity of GDCH in mice. The results revealed that GDCH upregulated the expressions of GDNF and CNTF in the mice under both normal and stress conditions, which resulted in neurotrophic effects on neural stem cells, astrocytes, and neurons. However, GDCH did not markedly alter the mRNA expressions ofBDNF,NGF,NT-3,VEGFα,FGF2, orIGF2in the hippocampus of the mice.
5.3 Neurons and nerves
5.3.1 Neuronal activity
Anxiety and depression cause the activation of nerve cells in different brain regions, however, protein hydrolysates and peptides can regulate neuronal expression in various ways to alleviate anxiety and depression symptoms. The expressions of c-Fos in different regions of the mouse brain, including the prefrontal cortex,hippocampal regions, accumbens nucleus, hypothalamus, amygdala,and nucleus tractus solitarius (NTS) have all been studied[29-30,70].An indicator of neuronal activation, c-Fos is a proto-oncogene that is expressed in response to external stimuli. It has been used to identify the neuronal circuits behind stress-induced neuroendocrine,autonomic and behavioral responses[103]. Benoit et al.[29]compared the modulation of neurons in the male Swiss mouse brain usingα-CZP and diazepam. After pressure stimulation of the mice with the light/dark box model, it was found that c-Fos expression was upregulated in different regions of the brain (prefrontal cortex, hippocampus,accumbens nucleus, and hypothalamus), whereas after the i.p.administration ofα-CZP, c-Fos expression was down-regulated in the hippocampus, accumbens nucleus, and hypothalamus, but unchanged in the prefrontal cortex and up-regulated in the amygdala, indicating a different regulation of neuronal activity compared to that of diazepam. Benoit et al.[30]further compared the modulation of neurons in the male Swiss mouse brain using YLGYL,α-CZP, and diazepam.Both YLGYL andα-CZP dramatically increased the number of c-Fos positive neurons in the amygdala, however, the two peptides demonstrated different modes of action in different regions of the amygdala. Theα-CZP was more effective in the medial nucleus,basolateral nucleus, and basomedial nucleus, whereas YLGYL was more valid in the anterior cortical nucleus and posterolateral cortical nucleus. On the other hand, diazepam proved more valuable in the NTS and prefrontal cortex, substantially increasing the number of c-Fos positive neurons in the NTS and amygdala central nucleus and greatly decreasing them in the prefrontal cortex.
Low molecular weight peptides have been shown to dramatically increase the density of proliferating cells and immature neurons in the hippocampus. Kakoi et al.[42]used 5-bromo-2-deoxyuridine (BrdU)to label proliferating cells in the hippocampus and further detected BrdU-positive cells via immunofluorescence staining and stained immature neurons in the hippocampus with doublecortin. It was found that lower molecular weight peptides enhanced hippocampal neurogenesis in C57BL/6J mice compared to higher molecular weight pig skin gelatin peptides. Another study detected ki67-positive cells in the hippocampal dentate gyrus via immunofluorescence staining and found that PO dipeptides significantly promoted the proliferation of neural progenitor cells, bothin vitroandin vivo[68].
In addition, in a study involving neurons, CRS and CORT injection were used to cause anxiety and depression-like behavior in C57BL/6 mice, which resulted in damage to the hippocampus and hypothalamus. However, subsequent treatment with EW and Trp oligopeptides derived from whey protein was able to counteract the negative effects of the CRS and CORT injection on the hippocampal CA3 tissue and neuropathy in the hypothalamus, consequently restoring the cells to a near-normal appearance. The EW was the more efficient of these two treatments[33].
5.3.2 Vagus nerve
Few studies have investigated the anxiolytic and antidepressant roles of nerve protein hydrolysates and peptides. Thus far, only two studies have found that peptides can exert any effect through the vagus nerve[48,70]. The vagus nerve is the main part of the parasympathetic nervous system, and is the tenth pair of brain nerves,extending from their origin in the brain stem through the neck and chest to the abdomen. The most important function of the vagus nerve is afferent, in which it transmits information from internal organs (such as the intestine, liver, heart, and lung) to the brain[104].
The peptides in the abovementioned studies were derived from soybean. In one, the researchers performed vagotomies on the mice and found that in the elevated plus maze test, oral administration of βCGα (323–333) increased the percentage of time and number of visits to the open arms in sham-treated mice, but showed no anxiolytic-like effects in the vagotomized mice[48]. Similarly,oral administration of soy-deprestatin shortened the immobility time of sham-treated mice in the tail suspension test, however, no antidepressant effect was demonstrated in the vagotomized mice[70].To further verify the soybean peptides’ activation of the vagus nerve,the two studies also detected the expression of c-Fos in the nucleus of the solitary tract (which receives vagus nerve input) and discovered that the two peptides enhanced the number of c-Fos positive cells in the NTS of mice. These results suggest that βCGα (323–333) and soy-deprestatin can exert anxiolytic and antidepressant-like effects,respectively, via the vagus nerve[48,70].
5.4 HPA axis
The HPA axis is a crucial stress system in the body, the fundamental mechanism of which is the release of CRH from the hypothalamus, which further stimulates the release of ACTH from the pituitary gland. ACTH, in turn, stimulates the adrenal glands to secrete CORT. Additionally, there is a negative feedback phenomenon in which the hypothalamus and pituitary gland are inhibited by the release of CORT, a glucocorticoid that is associated with mood,stress, anxiety, and cognitive effects[105-106]. Numerous studies have shown that hyperactivity of the HPA axis and the release of cortisol are associated with anxiety and depression[107-108].
Trp oligopeptide, Peptidyss, YL dipeptides, and GLS polysaccharides peptides have all been found to reduce the CORT levels in mice under stressful situations, thereby modulating the HPA axis to alleviate anxiety and depression[33,39,62,72]. The i.p. injection of YL dipeptides reduced serum CORT concentration and hypothalamicCRHmRNA expression in forced swim stressed mice[62], while the gavage of GLS polysaccharides peptides for 4 weeks significantly reduced serum CORT levels in mice under CUMS for 8 weeks[72].In addition to these findings, some studies have measured changes in CORT levels at different time points during stress. Dinel et al.[39]measured CORT levels in the plasma of BALB/c mice before the stress protocol (10 min in the open field test and 5 min in the elevated plus maze test), 30 min thereafter and 60 min thereafter and found no significant difference between the groups at 0 min and 60 min.However, at 30 min, the CORT levels were markedly higher in the stressed group than in the non-stressed group, while those in the Peptidyss-supplemented stressed group was significantly lower than those in the non-stressed group. When Zhu et al.[33]used a combination of CORT injection and restraint stress to establish an anxiety and depression model in mice, they found that plasma CRH and ACTH levels were higher in the model group, while CORT levels were lower. The reason for these results may have been that prolonged CORT injection leads to adrenal cortical atrophy and secretory dysfunction and, in turn, a decrease in CORT levels usually modulates the HPA system through a negative feedback mechanism.EW and WPH have been shown to ameliorate this phenomenon by reducing CRH and ACTH levels while elevating CORT levels in the plasma, thereby promoting the normalization of the HPA axis.
6. Conclusion and prospects
This review has shown comprehensively that anxiolytic and antidepressant protein hydrolysates and peptides derived from milk,fish, eggs, pig skin, bovine, spinach, soybean, wheat, and mushrooms can exert effects through neurotransmitter systems, neurotrophic functions, neurons, nerves, and the HPA axis. Furthermore, they share the following physicochemical traits: low molecular weight,ultraviolet absorption (A280nm), hydrophobic and stable, and contain Leu, Tyr, Pro, and Trp amino acids with aromatic amino acid leucine or YXXY sequence at N-terminal. However, it is also evident that the existing research has the following problems:
1) Most of the current studies used behavioral tests to assess the anxiolytic and antidepressant activities of animals, this method has the disadvantages of large individual differences, poor reproducibility,cumbersome operation, and susceptibility to influence. Subsequent studies should focus on identifying key indicators and combining behavioral tests to better evaluate the anxiolytic and antidepressant effects.
2) In addition to cell and animal experiments, there are also some studies showing the potential anxiolytic and antidepressant activity of protein hydrolysates and peptides in humans. However, a large number of human clinical trials are necessary to confirm the safety and efficacy of these peptides.
3) Protein hydrolysates and peptides may be affected by proteases as they pass through the gastrointestinal tract, requiring verification of their stability in the human body or optimization of delivery vehicles.Besides, attention should be paid to the anxiolytic and antidepressant effects of unabsorbed peptides in the intestine, such as changes in the intestinal flora, metabolite composition, and enteric nervous system.
4) The anxiolytic and antidepressant mechanisms of protein hydrolysates and peptides have not been fully and thoroughly elucidated. Endogenous biomarkers, pathways of action, and target genes need to be further explored. It can be screened and verified by reference to the targets of herbal medicine and other drugs, such as the reuptake of monoamines, the activity of affecting neuroreceptor binding and channel transporter protein, etc.
5) Studies on the relationship between the structure and activity of anxiolytic and antidepressant peptides are still lacking and incomprehensive. Molecular docking and molecular dynamics can be used to screen anxiolytic and antidepressant protein hydrolysates and peptides and determine the conformational relationships.
6) There are still many barriers to break through from basic research to product application, and most research is currently limited to laboratory production and has not yet been extended to the industrial level.
Overall, anxiolytic and antidepressant protein hydrolysates and peptides with high activity and few side effects are promising prospects, but there are still many challenges until they are applied to clinical patients.
Conflict of interest
Xinqi Liu is an editorial board member forFood Science and Human Wellnessand was not involved in the editorial review or the decision to publish this article. The authors declare no conflict of interest.
Acknowledgment
This work was supported by the National Key Research and Development Program of China (2021YFD2100402), the National Natural Science Foundation of China (81903275) and the Fund of the Cultivation Project of Double First-Class Disciplines of Food Science and Engineering, Beijing Technology & Business University(BTBUYXTD202203).
杂志排行
食品科学与人类健康(英文)的其它文章
- Betalains protect various body organs through antioxidant and anti-inf lammatory pathways
- Effects of Maillard reaction and its product AGEs on aging and age-related diseases
- Characterization of physicochemical and immunogenic properties of allergenic proteins altered by food processing: a review
- Polyphenol components in black chokeberry (Aronia melanocarpa)as clinically proven diseases control factors—an overview
- Recent advances in the study of epitopes, allergens and immunologic cross-reactivity of edible mango
- Learning about good nutrition with the 5-color front-of-package label“Nutri-Score”: an experimental study