外源γ-氨基丁酸对芥蓝生长及品质的影响
2024-01-22高晓萍常有麟刘思恬韩康宁颉建明
刘 阳, 张 婧, 高晓萍, 常有麟, 刘思恬, 杨 滟, 韩康宁, 颉建明
(甘肃农业大学园艺学院,甘肃 兰州 730070)
芥蓝(BrassicaalbograbraL.H.Bailey)是十字花科芸薹属蔬菜,富含硫代葡萄糖苷、游离氨基酸、类胡萝卜素、维生素C和酚类化合物等多种营养物质,并且含有少量有机碱及金鸡纳霜,有益于消化,还具有解毒清热的功效[1-3]。
γ-氨基丁酸(γ-Aminobutyric acid, GABA)是细胞游离氨基酸库中重要的组成部分[4-5],其广泛存在于植物的各器官中,可为植物提供营养元素、调节碳氮代谢,影响植物生长发育、形态建成、物质积累和作物产量形成等各种生理生化过程[6-9]。Hijaz等[10]发现,GABA与植物激素协同作用,GABA的外源性应用增加了柑橘类植物中大多数植物激素的含量,促进植株生长。Li等[11]将GABA作用于低温弱光下的辣椒幼苗,发现其显著提高光合能力和内源GABA含量,增强抗氧化酶活性。研究结果表明,外源添加GABA可显著提高番茄植株中酚类物质和内源GABA含量[12]。Nayyar等[13]发现,添加GABA可以在一定程度上减轻光合损伤,缓解高温胁迫对玉米幼苗的伤害。外源GABA处理显著促进番茄幼苗体内可溶性蛋白质和可溶性糖的积累,降低活性氧产生速率,从而缓解干旱胁迫对幼苗生长的抑制作用[14]。此外,Ma等[15]发现,外源GABA诱导大麦幼苗合成内源GABA和其他游离氨基酸。近年来GABA对植物生理代谢影响的研究多数集中在提高植物对盐胁迫、低温胁迫等抗逆性方面,且多数是针对苗期开展试验研究,而对GABA在芥蓝生长和品质等方面作用的研究鲜有报道。本试验研究外源GABA对芥蓝植株生长及产品品质的影响,探索芥蓝高产高质栽培的新途径。
1 材料与方法
1.1 试验材料
供试芥蓝品种为农宝,其抗逆性和抗病性强,丰产性好,品质优。芥蓝种子于50~55 ℃浸种15 min后,用蒸馏水漂洗3~5次,将消毒处理后的种子置于恒温摇床(摇床转速为180 r/min),在28 ℃黑暗条件下振荡培养2 d,每12 h更换1次超纯水,待80%的种子露白后播于穴盘中。待芥蓝幼苗长到五叶一心,选取生长一致且无病虫害的植株移入花盆,每盆1株。
1.2 试验设计
试验采用随机区组设计,设置6个不同浓度的GABA溶液,分别为:0 mmol/L(CK)、25 mmol/L(T1)、50 mmol/L(T2)、75 mmol/L(T3)、100 mmol/L(T4)、125 mmol/L(T5),其中CK以等量清水替代,所有处理中均加入0.1%吐温80。芥蓝幼苗定植15 d后,于9:00对芥蓝叶片正反面均匀喷施GABA溶液,每3 d处理1次,共4次。处理结束后,第6 d对芥蓝的可食用器官(叶片和花茎)进行取样,测定相关指标。每处理9株,重复3次。
1.3 测定指标及测定方法
1.3.1 形态指标测定 测定植株茎粗(用游标卡尺测量)、株高(用直尺测量)、地上和地下部干质量及鲜质量。
1.3.2 品质指标测定 可溶性糖含量测定采用蒽酮比色法,维生素C含量测定采用2,6-二氯酚靛酚钠比色法,可溶性蛋白质含量测定采用考马斯亮蓝比色法,硝酸盐含量测定采用磺基水杨酸比色法,游离氨基酸含量测定采用茚三酮比色法,总酚含量测定采用福林酚比色法,类黄酮含量测定采用比色法[16],总硫代葡萄糖苷含量测定采用氯化钯比色法[17]。
1.3.3 叶绿素含量测定 参照李静[18]的方法测定植株叶片叶绿素含量。
1.3.4 光合参数测定 用CIRAS-2便携式光合仪(美国PP SYSTEM公司产品),于晴天9:00-11:30测定植株的净光合速率(Pn)、胞间CO2浓度(Ci)、气孔导度(Gs)和蒸腾速率(Tr)。
1.3.5 叶绿素荧光参数测定 用IMAPING-PAM调制叶绿素荧光成像系统(德国WALZ公司产品)测定植株叶片的PSⅡ最大光化学效率(Fv/Fm)、实际光能转换效率(ΦPSⅡ)、光化学淬灭系数(qP)和非光化学淬灭系数(NPQ)。
1.3.6 内源激素测定 参照Mao等[19]的方法并稍作修改,用Aglient series 1100高效液相色谱仪测定植株叶片玉米素(ZT)、赤霉素(GA3)、生长素(IAA)和脱落酸(ABA)含量。色谱条件:ZORBAX SB-C18色谱柱(色谱柱直径和长度分别为4.6 mm和250.0 mm;进样通道的直径:5 μm);流动相为甲醇和0.1%磷酸溶液(1∶9,体积比);流速为0.1 ml/min;检测波长为254 nm;柱温为30 ℃;进样量为10 μl。
1.3.7 内源GABA相关指标测定 GABA含量测定参照Hu等[20]的方法并稍加修改,谷氨酸含量测定参照Kennedy等[21]的方法,谷氨酸脱羧酶(GAD)活性测定参照Bartyzell等[22]的方法并稍加修改。
1.4 数据统计分析
采用Excel 2021进行数据处理与作图,试验数据的统计分析采用SPSS 20.0,并采用Duncan’s检验对显著性进行多重比较(α=0.05)。
2 结果与分析
2.1 外源GABA对芥蓝生长的影响
从表1可以看出,与CK相比,大部分浓度GABA处理均可增加芥蓝生物量。T3、T4、T5处理芥蓝的株高均显著高于CK,分别增加21.19%、12.78%和6.43%。T1~T5处理均显著增加芥蓝的茎粗,较CK分别增加14.48%、21.41%、41.59%、23.34%和16.72%。T2、T3、T4处理的地上部鲜质量和干质量显著高于CK,其中T3处理最高,较CK分别增加38.57%和50.21%,T1~T3处理的地下部鲜质量和干质量均显著高于CK,T3处理最高,较CK分别增加97.87%和63.16%。

表1 外源γ-氨基丁酸对芥蓝生长的影响
2.2 外源GABA对芥蓝品质的影响
2.2.1 外源GABA对芥蓝可溶性糖和维生素C含量的影响 外源GABA处理后芥蓝叶片和花茎中可溶性糖含量呈先增加后降低趋势,维生素C含量表现出相似的趋势(图1)。芥蓝叶片和花茎中可溶性糖含量在GABA处理下均显著高于CK,其中T3处理可溶性糖含量分别提高59.37%和52.84%。与CK相比,T2、T3处理显著增加芥蓝叶片和花茎中维生素C 含量,T3处理含量最高,分别提高13.83%和15.61%。

CK、T1、T2、T3、T4、T5见表1注。不同小写字母表示各处理之间差异显著(P<0.05)。图1 外源γ-氨基丁酸对芥蓝可溶性糖和维生素C含量的影响Fig.1 Effect of exogenous γ-aminobutyric acid on soluble sugar and vitamin C contents of Chinese kale
2.2.2 外源GABA对芥蓝可溶性蛋白质、游离氨基酸和硝酸盐含量的影响 适宜浓度的GABA处理可促进芥蓝叶片和花茎中可溶性蛋白质和游离氨基酸的积累,叶片中可溶性蛋白质和游离氨基酸含量均显著高于花茎。如图2A所示,T1~T5处理芥蓝叶片中可溶性蛋白质含量均显著高于CK,T3处理最高,较CK提高24.73%,部分处理间差异显著;T2、T3和T4处理花茎的可溶性蛋白质含量均显著高于CK,处理间无显著差异。T2、T3、T4、T5处理芥蓝叶片和花茎中游离氨基酸含量均显著高于CK,其中T2、T3、T4、T5处理叶片游离氨基酸含量较CK分别增加16.58%、42.22%、38.04%、19.21%,T2、T3、T4、T5处理花茎游离氨基酸含量较CK分别增加20.07%、53.67%、44.42%和31.32%(图2B)。
外源GABA处理均能降低芥蓝叶片和花茎中硝酸盐含量(图2C)。与CK相比,T2、T3、T4、T5处理均显著降低了芥蓝叶片和花茎中硝酸盐含量,其中T3处理分别降低24.69%和17.11%。

CK、T1、T2、T3、T4、T5见表1注。不同小写字母表示各处理之间差异显著(P<0.05)。图2 外源γ-氨基丁酸对芥蓝可溶性蛋白质、游离氨基酸和硝酸盐含量的影响Fig.2 Effect of exogenous γ-aminobutyric acid on soluble protein, free amino acid and nitrate contents of Chinese kale
2.2.3 外源GABA对芥蓝总硫代葡萄糖苷含量的影响 不同浓度GABA处理均能促进芥蓝叶片和花茎中总硫代葡萄糖苷的积累,且随GABA处理浓度的增加呈先上升后下降的趋势(图3)。与CK相比,T2、T3、T4处理芥蓝叶片和花茎中总硫代葡萄糖苷含量均显著增加,其中T3处理总硫代葡萄糖苷的含量最高,分别增加28.15%和35.79%。

CK、T1、T2、T3、T4、T5见表1注。不同小写字母表示各处理之间差异显著(P<0.05)。图3 外源γ-氨基丁酸对芥蓝总硫代葡萄糖苷含量的影响Fig.3 Effect of exogenous γ-aminobutyric acid on total glucosinolate content of Chinese kale
2.2.4 外源GABA对芥蓝总酚和类黄酮含量的影响 由图4可看出,芥蓝总酚和类黄酮含量随GABA浓度的增加呈先增后减趋势。T2、T3处理芥蓝总酚含量均显著增加,叶片中总酚含量在T3处理最高,较CK增加12.44%,花茎中总酚含量在T2处理最高,较CK增加43.06%(图4A)。如图4B所示,T3处理显著增加芥蓝叶片和花茎中类黄酮含量,较CK分别增加29.55%和38.03%,其他处理间无显著差异。

CK、T1、T2、T3、T4、T5见表1注。不同小写字母表示各处理之间差异显著(P<0.05)。图4 外源γ-氨基丁酸对芥蓝总酚和类黄酮含量的影响Fig.4 Effect of exogenous γ-aminobutyric acid on total phenolics and flavonoid contents of Chinese kale
2.3 外源GABA对芥蓝生理特性的影响
2.3.1 外源GABA对芥蓝叶片光合色素含量的影响 不同浓度GABA处理均可不同程度地提高芥蓝叶片光合色素含量,且随GABA浓度的增加呈先增后减的趋势(图5)。T1、T2、T3和T4处理下芥蓝叶片的叶绿素a、叶绿素b和叶绿素a+叶绿素b含量均显著高于CK,其中T3处理的含量最高,较CK分别增加32.62%、37.25%和34.05%。
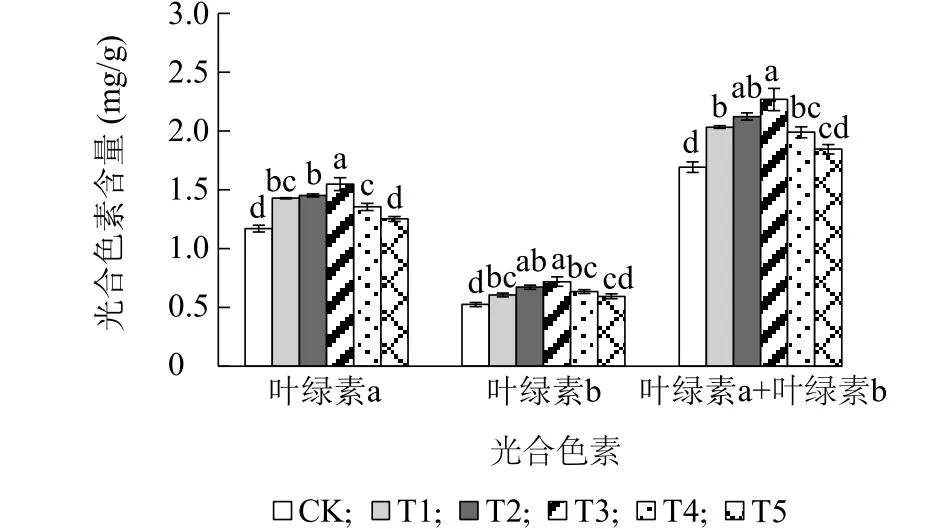
CK、T1、T2、T3、T4、T5见表1注。不同小写字母表示各处理之间差异显著(P<0.05)。图5 外源γ-氨基丁酸对芥蓝叶片光合色素含量的影响Fig.5 Effect of exogenous γ-aminobutyric acid on photosynthetic pigment contents of Chinese kale leaves
2.3.2 外源GABA对芥蓝叶片光合气体交换参数的影响 外源GABA对芥蓝叶片光合特性的影响如图6所示,Pn、Gs和Tr随GABA处理浓度的增加呈先增后减的趋势,而Ci呈现降低的趋势。T1、T2、T3、T4处理叶片Pn、Gs和Tr均显著高于CK,其中T3处理下芥蓝叶片的Pn和Tr最高,较CK分别增加52.06%和51.11%。叶片Gs在T2处理下最大,较CK增加92.75%。不同浓度GABA处理均显著降低了叶片的Ci,其中T3处理最低,较CK降低了15.41%。
2.3.3 外源GABA对芥蓝叶片叶绿素荧光参数的影响 不同浓度GABA处理对芥蓝叶片叶绿素荧光参数的影响如表2所示,Fv/Fm随GABA浓度增加呈现先上升后下降的趋势,ΦPSⅡ和qP在一定范围内呈逐渐上升的趋势,而NPQ则在一定范围内呈现下降的趋势。T1~T5处理芥蓝叶片中Fv/Fm、ΦPSⅡ和qP均显著高于CK,Fv/Fm以T3处理最大,较CK提高4.35%,ΦPSⅡ和qP分别以T4、T2处理最大,较CK分别增加24.34%和21.40%。T2、T3、T4、T5处理均可显著降低芥蓝叶片NPQ,较CK分别降低了15.84%、25.51%、29.91%和17.89%。
2.3.4 外源GABA对芥蓝叶片内源激素的影响 芥蓝叶片中ZT、GA3、IAA和ABA等内源激素含量随GABA处理浓度的增加呈先增后减的趋势。如图7A所示,T1、T2、T3处理均显著增加叶片中ZT含量,较CK分别增加16.11%、66.31%和72.41%。如图7B、图7C所示,T1~T4处理叶片中GA3和IAA含量均显著高于CK,其中T3处理GA3和IAA含量最高,较CK分别增加98.68%和84.37%。不同浓度GABA处理叶片的ABA含量均显著高于CK,其中T3处理较CK提高38.72%(图7D)。
2.3.5 外源GABA对芥蓝GABA代谢的影响 外源GABA处理对芥蓝的内源GABA含量、谷氨酸含量的影响均呈先增后减的趋势。如图8A所示,T2、T3、T4、T5处理下芥蓝叶片和花茎内源GABA含量显著高于CK,其中T3处理最大,较CK分别增加39.26%和60.29%,各处理表现为T3>T4>T5>T2>T1>CK,且叶片中内源GABA含量显著高于花茎。T2、T3、T4、T5处理下芥蓝叶片和花茎中谷氨酸含量均显著高于CK,其中T3处理谷氨酸含量最高,较CK分别增加32.41%和14.28%(图8B)。芥蓝叶片和花茎中谷氨酸脱羧酶活性随GABA浓度的增加呈现先增加后降低的趋势,T2、T3、T4和T5处理GAD活性均显著高于CK,在T3处理下GAD活性达到最大值,较CK分别增加56.13%和66.53%(图8C)。

CK、T1、T2、T3、T4、T5见表1注。不同小写字母表示各处理之间差异显著(P<0.05)。图6 外源γ-氨基丁酸对芥蓝叶片光合参数的影响Fig.6 Effect of exogenous γ-aminobutyric acid on photosynthetic parameters of Chinese kale leaves
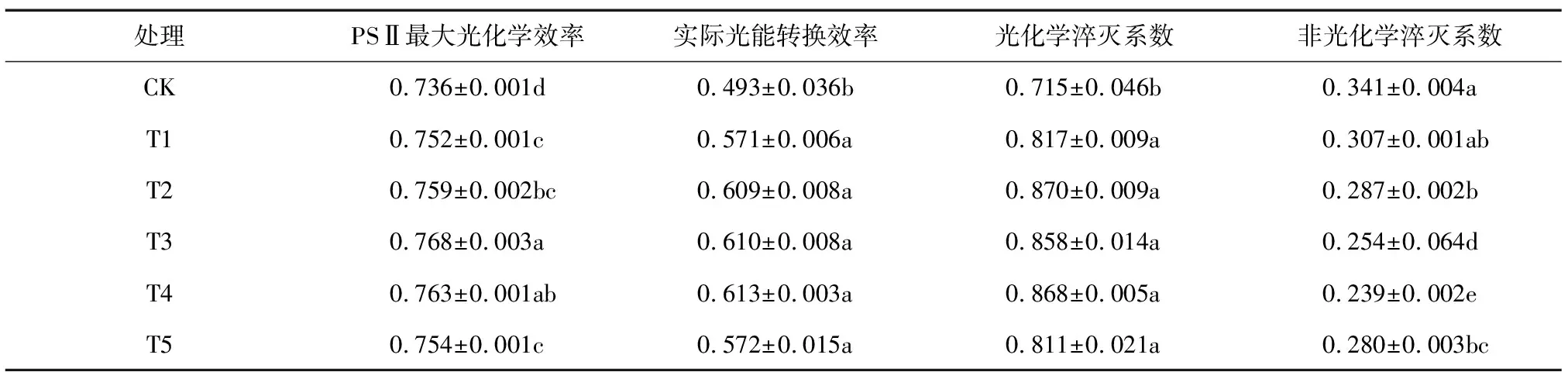
表2 外源γ-氨基丁酸对芥蓝叶片叶绿素荧光参数的影响

CK、T1、T2、T3、T4、T5见表1注。不同小写字母表示各处理之间差异显著(P<0.05)。图7 外源γ-氨基丁酸对芥蓝叶片内源激素水平的影响Fig.7 Effect of exogenous γ-aminobutyric acid on endogenous hormone level in leaves of Chinese kale

CK、T1、T2、T3、T4、T5见表1注。不同小写字母表示各处理之间差异显著(P<0.05)。图8 外源γ-氨基丁酸对芥蓝内源γ-氨基丁酸代谢的影响Fig.8 Effect of exogenous γ-aminobutyric acid on endogenous γ-aminobutyric acid metabolism of Chinese kale
2.4 芥蓝生长、品质及生理特性的相关分析
由表3可以看出,除Ci和硝酸盐含量外,其他生长、品质及生理特性指标之间均表现出正相关关系,且部分指标间达到极显著正相关。鲜质量与可溶性糖含量、可溶性蛋白质含量、总硫代葡萄糖苷含量、谷氨酸含量、Pn、Fv/Fm、IAA含量和ABA含量均呈显著正相关(P<0.05);Pn与可溶性蛋白质含量、总硫代葡萄糖苷含量、总酚含量、谷氨酸含量、叶绿素含量、Gs、Tr、Fv/Fm、ZT含量、GA3含量和IAA含量均呈显著正相关;内源激素与生长生理指标之间的相关性较强;内源GABA含量与可溶性糖和游离氨基酸含量呈极显著正相关(P<0.01),与株高、Fv/Fm和ABA含量均呈显著正相关。
3 讨 论
GABA是一种新发现的参与植物生长、发育和多种生理代谢的信号分子[23]。黄娟等[24]的研究结果表明,外源GABA能促进高温胁迫下黄瓜幼苗的生长,增强黄瓜叶片的光合作用。本研究中,外源喷施不同浓度GABA均能增加芥蓝的株高、茎粗、鲜质量和干质量,表明GABA处理能促进芥蓝生长和生物量的积累。光合作用能将光能转化为有机物并放出能量,是植物干物质积累和产量形成的基础,提高光合能力有助于植物生长发育及生物量积累。叶绿素含量与光合能力密切相关,是反映植物生长状况的重要指标,还可以反映蔬菜的色泽等外观品质。叶绿素荧光参数反映不同环境下植株光合电子转换过程的变化[25]。叶面喷施GABA可改善生菜的光合功能,调控植株的生长[26]。本研究发现,不同浓度GABA均可在不同程度上提高芥蓝叶片的叶绿素含量,从而提高光合效率。同时,GABA处理能有效提高芥蓝叶片的Pn、Tr和Gs,增强芥蓝叶片的光合能力,显著降低Ci,提高叶片对CO2的利用率,促进光合物质的积累,这与李武等[27]的研究结果相似。此外,外源GABA处理能够提高芥蓝叶片Fv/Fm、ΦPSⅡ和qP,显著降低NPQ值,说明GABA能提高芥蓝对光能的捕获和利用效率,减少能量的耗散,有利于把所捕获的光能尽可能地用于光合作用[28],从而促进芥蓝生长。植物激素对植物的生长发育具有重要的调控作用。GABA与生长素、脱落酸、赤霉素、细胞分裂素(CTK)之间具有相互调控的关系[29]。一般情况下,赤霉素和生长素能促进植株的生长和发育[30-31];玉米素能抑制叶绿素分解,促进气孔开放、叶片生长及叶绿体发育,延缓植株衰老[32]。在植物生长发育的不同时期,各激素相互作用,共同调节植物生长发育及生理过程[29]。研究结果表明,外源GABA可显著提高植物IAA、茉莉酸(JA)、ABA、玉米素核苷(ZR)等内源激素的含量[33]。本研究中,经GABA处理的芥蓝叶片中ZT含量、ABA含量和IAA含量均有不同程度的增加,说明外源GABA处理可以增加芥蓝内源激素含量,从而促进芥蓝生长;而125 mmol/L GABA处理的GA3含量显著低于CK,表明较高浓度的GABA处理抑制芥蓝内GA3的积累。

表3 芥蓝生长、品质及生理特性指标的相关性
GABA也是植物体内普遍存在的一种氨基酸,能直接被吸收利用,可为植株提供氮源,调节体内氮代谢,减少硝酸盐含量。GABA通过降低氨基酸合成所用能量,提高可溶性糖、可溶性蛋白质和维生素C等的含量,从而改善蔬菜的营养品质[29, 34]。本研究发现,外源GABA处理可明显提高芥蓝的品质,但存在一定的浓度效应。可溶性糖、可溶性蛋白质、维生素C和总硫代葡萄糖苷含量等是衡量芥蓝品质的重要指标。可溶性糖是植物光合作用的直接产物[35],通过检测植株可溶性糖含量的变化,可以部分了解叶片光合产物的积累情况。外源GABA处理可显著增加芥蓝叶片和花茎中可溶性糖含量,随着处理浓度的增加,可溶性糖含量呈先增后减趋势,但均高于CK,与张换换等[36]的研究结果相似。同时,植株叶片中约有50%的可溶性蛋白质是光合作用的关键酶[35]。在本研究中,外源GABA喷施处理可提高芥蓝可溶性蛋白质含量,可溶性蛋白质含量提高时,可间接增强光合关键酶的活性,增强植株的光合能力。芥蓝叶片和花茎中维生素C含量在50 mmol/L和75 mmol/L GABA处理下均明显提高。外源GABA处理也可促进游离氨基酸在芥蓝叶片和花茎中的积累,芥蓝叶片中的游离氨基酸含量均高于花茎,且75 mmol/L和100 mmol/L GABA处理下游离氨基酸积累量相对其他处理较高,弓瑞娟等[37]研究发现外源GABA促进芥蓝中游离氨基酸的积累,与本研究结果相似。此外,不同浓度GABA处理降低芥蓝叶片和花茎中硝酸盐含量,这与李敬蕊等[38]的研究结果相似。本研究还发现芥蓝中硝酸盐含量与游离氨基酸含量变化趋势相反,这主要是因为GABA加速了硝态氮的还原与转化,将植株体内积累的硝态氮转化为铵态氮以合成氨基酸[39]。硫苷是一组含氮含硫的次级代谢物,其降解产物不仅与蔬菜的风味形成有关,还能降低癌症发生率[40]。外源氨基酸处理能显著提高芥蓝中总硫代葡萄糖苷含量,促进维生素C和可溶性蛋白质的积累[41]。本研究中,外源GABA处理显著提高芥蓝叶片和花茎中总硫代葡萄糖苷含量,75 mmol/L和100 mmol/L GABA处理效果显著优于其他处理,这与周芷亦[42]的研究结果相似,氨基酸是硫苷的合成前体,因此GABA中的碳氮骨架有助于硫苷的形成,但随GABA浓度的增加,硫苷含量逐渐下降,表明高浓度GABA可能抑制了芥蓝叶片和花茎中硫苷的积累。总酚和类黄酮是评价蔬菜品质的重要指标,但也能够起到清除自由基的作用[43]。植物中多酚含量越高,自由基清除能力越强[44]。在本研究中,随GABA浓度增加,芥蓝中类黄酮和总酚含量先增加后减少,不同浓度GABA处理均能不同程度地增加类黄酮和总酚含量,表明GABA处理能有效促进总酚和类黄酮的合成,多酚含量的增加可能是由于GABA促进了植物对必需营养元素吸收[45]。
GABA主要是谷氨酸在GAD催化下合成的,GABA可通过分流进入三羧酸循环,作为替代呼吸底物参与植物能量供应[46]。研究发现,内源性的GABA水平可以通过外源性GABA的应用来增加,但取决于GABA的浓度[15]。在本研究中,外源GABA显著增加芥蓝叶片和花茎中内源GABA的浓度,75 mmol/L GABA处理的芥蓝内源GABA浓度达到最大值。芥蓝中谷氨酸含量在多数浓度GABA处理下均显著高于对照,且75 mmol/L GABA喷施处理的效果较为显著。GAD是GABA合成的关键酶,植株体内GABA水平受GAD活性的调节,外源GABA处理显著提高芥蓝叶片和花茎中GAD活性,随着处理浓度的增加,GAD活性有所降低,可能是由于高浓度的GABA抑制GAD活性,进而抑制谷氨酸转化为GABA,此外,GAD活性的增加促进芥蓝叶片和花茎中内源GABA的合成和积累,这与王贺[47]的研究结果相似。因此,外源GABA条件下,GABA支路中谷氨酸含量显著上升,提高GAD的活性,促使更多的谷氨酸合成GABA,增强GABA支路的效率,进而调节植物的生长发育。此外,植物吸收的外源GABA通过GAD催化谷氨酸生成α-酮戊二酸,从而为植物提供碳骨架,碳骨架与氮素合成氨基酸,从而提高蔬菜品质。
4 结 论
外源喷施适宜浓度GABA(75 mmol/L)可促进芥蓝生物量的积累,显著增加芥蓝叶片光合色素含量,增强光合作用,提高ZT、GA3、IAA、ABA等内源激素含量,促进植株生长发育;显著促进叶片和花茎中维生素C、可溶性糖、游离氨基酸及总硫代葡萄糖苷等的积累,促进酚类化合物的合成,降低硝酸盐含量,提高芥蓝的产量和品质;显著促进GABA和谷氨酸的积累,增强GAD活性,从而调控内源GABA代谢。
