基于优化MaxEnt模型的中国紫堇属植物生境适宜性研究
2024-01-17陈程浩龙主多杰陆徐伟宋扎磋孙芳慧索南吉
陈程浩,龙主多杰,陆徐伟,宋扎磋,苗 琪,孙芳慧,索南吉,2,3,*
1 青海师范大学生命科学学院,西宁 810016 2 青海省人民政府-北京师范大学 高原科学与可持续发展研究院,西宁 810016 3 青海省青藏高原生物多样性形成机制与综合利用重点实验室,西宁 810016
全球气候变化直接影响着物种的栖息环境,如生态系统结构和功能[1],植物的春季物候[2],水热环境[3]等,进而对植物的地理分布格局造成深刻的影响[4]。青藏高原地区对气候变化尤为敏感[4],许多学者认为水热条件是影响青藏高原地区植物分布的关键因素[5],由于气候变化对青藏高原水热条件的影响[3],使该地区植物在未来将向更高纬度和更高海拔的地区迁移[6]。在全球气候变化的背景下,青藏高原地区植物的生存受到严重的威胁[7],了解气候变化对植物地理分布格局的影响可为青藏高原地区的物种保护奠定基础[8]。
物种分布模型是评价气候变化对物种分布影响的有力工具之一[9],其中最大熵(Maximum Entropy, MaxEnt)模型具有仅依靠物种分布点及环境数据便能准确可靠地模拟物种的潜在适生区[10],运算时间少,且在低样本量条件下模拟精度高等优点[11]。近年来,MaxEnt模型因在预测气候变化对植物(如高山栎组、喜旱莲子草等)分布影响的研究中展现出了优秀的预测能力而得到了广泛应用[12-13],如国际热带农业中心(CIAT)曾利用该模型预测了全球气候变化下全球主要产区小粒咖啡适宜性的变化[14]。
紫堇属(Corydalis)是罂粟科(Papaveraceae)中最大的属[15],主要分布于横断山脉及青藏高原地区[16]。横断山区地形结构复杂,海拔高差大,复杂的环境为紫堇属植物应对第四纪冰期提供了避难所,使紫堇属植物在此地区具有较高的物种多样性[17],是该地区物种组成中非常重要的一部分。为应对复杂的地形地貌环境,紫堇属植物形成了复杂的物种分化及形态变异,是分类学上最有挑战性的类群[18]。另外,紫堇属部分物种具有较高的药用价值[19-21]和观赏价值[22]。近年来,较多学者的研究集中在紫堇属植物的系统发育、分类[23]以及药用价值上[24],但对紫堇属物种分布格局对气候变化的响应鲜有报道。
为探索不同时期紫堇属物种分布格局,本文运用生态位模型(ecological niche models, ENMs)中的MaxEnt模型对末次盛冰期(last glacial maximum,LGM)、当代(Current)和两种温室气体排放场景下的未来4个时期,共6个时期的环境数据对紫堇属植物的潜在适生区进行模拟,并对不同时期间两种温室气体排放模式下的潜在适生区的变化、质心转移及保护现状进行了分析。以期回答以下三个科学问题:(1)影响中国紫堇属植物分布的主要环境因素是什么;(2)气候变化下紫堇属植物的潜在适生区如何变化;(3)中国紫堇属植物的保护状况如何?
1 材料与方法
1.1 数据与材料
1.1.1紫堇属物种名录及分布数据
本研究所使用的紫堇属物种名录主要来自于物种2000中国节点 《中国生物物种名录》2022年版,并参考《中国植物志》FRPS(Flora Reipublicae Popularis Sinicae)电子版(http://www.iplant.cn/frps2019/),依据接受名和异名确定了中国紫堇属植物的接受名录[25]。最终得到中国境内共有紫堇属植物共403种。
根据以上所确定的紫堇属植物的物种名,在全球生物多样性信息网络数据库(https://www.gbif.org/zh/)[26]、iNaturalist(https://www.inaturalist.org/)[27]和中国植物标本馆(https://www.cvh.ac.cn/)[28]中查找紫堇属植物位于中国的分布数据,通过ArcGIS10.4剔除分布于中国范围以外的数据,然后通过R4.2.3中的“CoordinateCleaner”包清洗重复、分布于海洋及围绕首都和国家中心的数据[29],清洗后的分布记录为3963条。为了避免人为采集标本的偏向性造成物种分布记录集群效应的误差,通过EMTools工具设置了2.5`分辨率的栅格,在每个栅格内筛选出离中心点最近的一个分布点进行保留[30]。最终得到中国紫堇属植物的有效分布点共1910个(图1),并获取了由太阳辐射量所确定的气候分区区划。

图1 筛选后紫堇属分布记录Fig.1 Distribution record of Corydalis after screening
1.2 环境数据
1.2.1环境数据获取、异常值处理及筛选
本研究共获取了分辨率均为2.5′的22个环境数据(表1),其中19个为气候因子数据,3个为地形因子数据,即海拔(Elevation)、坡向(Aspect)和坡度(Slope)。LGM数据选自IPCC第五次耦合模型比较项目(Coupled Model Intercomparison Project 5,CMIP5)中模拟中国气候较好的CCSM4大气环流模型所构建的在WorldClim1.4(https://worldclim.org/data/index.html)上的数据;Current数据以及未来环境数据均选取自IPCC第六次耦合模型比较项目(Coupled Model Intercomparison Project 6,CMIP6)构建的在WorldClim2.1(https://worldclim.org/data/index.html)上的数据[26]。未来环境数据选取了用共享社会经济路径(Shared Socio-economic Pathways, SSP)来表示的两种温室气体排放场景,即低浓度温室气体排放场景(SSP126)和高浓度温室气体排放场景(SSP585)下的四个时期,分别为2021-2040(2040s)、2041-2060(2060s)、2061-2080(2080s)、2081-2100(2100s)[31-32]。

表1 影响紫堇属物种分布环境变量的贡献率及重要性Table 1 Contribution rate and importance of environmental variables affecting species distribution of Corydalis
通过R包“outlisers”对环境异常值数据进行了纠正。WorldClim1.4(https://www.worldclim.org/data/v1.4/formats.html)中为了减少栅格数据下载体积,对温度相关数据进行了正常温度数值乘10的处理,因此本文对LGM的温度相关数据,即年平均气温(bio1)、平均气温日较差(bio2)、等温性(bio3)、气温季节性变动系数(bio4)、最暖月的最高温度(bio5)、最冷月份的最低温度(bio6)、气温年较差(bio7)、最湿季的平均温度(bio8)、最干燥季度的平均温度(bio9)、最暖季的平均温度(bio10)、最冷季的平均温度(bio11),用ArcGIS10.4的“栅格分析-除”工具对栅格数据值进行了除以10的处理,使其可以与WorldClim2.1中数据同时进行分析。
为避免环境数据间多重共线性造成模型的过度拟合[33],本研究根据模型预实验得出的贡献率(表1)以及通过ArcGIS10.4的“数据管理工具-栅格-栅格处理-重采样”工具提取的数据,进一步对环境数据的空间自相关性进行了Spearman分析。根据分析结果(图2),剔除了相关性系数大于0.8以及贡献率较小的环境数据[34]。最终筛选出具有统计学和生物学意义的共14个环境数据进行建模(表2)。

表2 MaxEnt模型各主导环境变量的重要性Table 2 The importance of dominant environmental variables in MaxEnt model

图2 22个环境变量的空间自相关性Fig.2 Spatial autocorrelation of environmental variablesbio1:年平均气温Annual Mean Temperature; bio2:平均气温日较差Mean Diurnal Range; bio3:等温性Isothermality; bio4: 气温季节性变动系数Seasonal variation coefficient of temperature; bio5: 最暖月的最高温度Max Temperature of Warmest Month; bio6: 最冷月份的最低温度Min Temperature of Coldest Month; bio7:气温年较差Temperature Annual Range; bio8: 最湿季的平均温度Mean Temperature of Wettest Quarter; bio9: 最干燥季度的平均温度Mean Temperature of Driest Quarter; bio10: 最暖季的平均温度Mean Temperature of Warmest Quarter; bio11: 最冷季的平均气温Mean Temperature of Coldest Quarter; bio12: 年降水量Annual Precipitation; bio13: 最湿月的降水量Precipitation of Wettest Month; bio14: 最干燥月份的降水量Precipitation of Driest Month; bio15: 季节性降水Precipitation Seasonality; bio16: 最湿季降水Precipitation of Wettest Quarter; bio17: 最干燥季的降水Precipitation of Driest Quarter; bio18: 最暖季降水Precipitation of Warmest Quarter; bio19: 最冷季降水Precipitation of Coldest Quarter; bio20:海拔Elevation; bio21: 坡向 Aspect; bio22: 坡度 Slope
1.3 模型优化、构建及精度评估
MaxEnt模型的默认参数是其开发者在对鸟类、哺乳类、爬行类和植物分布的研究中,根据其模型结果与现实分布的比较而设置的[35]。但由于MaxEnt模型是机器学习中的复杂模型,其对采样偏差敏感,且模型的复杂度与特征函数(Feature combination,FC):LQPTH(L为线性liner、T为阈值性threshold、Q为二次型quadratic、H为片段化hinge、P为乘积型threshold)和倍频(Regularization multiplier,RM)显著相关[36]。为了避免过度拟合对模型迁移能力的影响,本文运用R4.2.3中的”Kuenm”包对模型的RM(0-4)从0开始每0.1向上递增到4与FC(LQPTH)分别进行交叉组合验证,最终选择具有显著性,数据遗漏率(Omission rates)<5%,且delta AICc值(校准后的最优模型与当前模型的赤池信息量准则的差值)较小的组合作为最优参数组合来运行最终的模型[37]。
MaxEnt模型的其余参数设置为:输出方式(Output format)选择为Logistic、勾选随机种子(Random seed)、随机测试百分比(Random test percentage)设为25%、重复次数(Replicates)设为10次[38]。用ArcGIS10.6中的“栅格分析工具-重分类”工具,通过自然断点法将MaxEnt模型输出的结果划分为:非潜在适生区(0-0.135)、低潜在适生区(0.135-0.306)、中潜在适生区(0.306-0.477)、高潜在适生区(0.477-0.880)[26]。
采用受试者特征工作曲线(receiver operating characteristic curve,ROC曲线)中的AUC值验证模型的精度,AUC数值在0-1之间,且数值越大模型可信性越高[39]。通常认为AUC值在0.5-0.7之间,表明模型的预测能力一般;0.7-0.9之间,表明模型的预测能力较好;0.9-1之间,表明模型的预测能力极好[40]。
1.4 潜在适生区变化及质心转移
通过ArcGIS10.6中SDMTools中的“Quick Reclassify to Binary”工具[41],根据MaxEnt模型输出结果中的栖息地适宜度指数(habitat suitability index,HIS),将HIS大于最大测试敏感性(Maximum test sensitivity)和特异性逻辑阈值(maximum test sensitivity plus specificity logistic threshold)的区域划分为潜在适生区,将HIS小于最大测试敏感性和特异性逻辑阈值的区域划分为非适宜区,从而获得只有潜在适生区与非潜在适生区的二值化栅格文件[42]。通过SDMTools中的“Distribution Change Between Binary SDMs”工具,计算出不同时期紫堇属植物潜在适生区的变化,并将其值定义为:0为不适生、-1为扩张、1为稳定区和2为收缩,计算不同时期潜在适生区的变化面积[43]。
根据以上不同时期紫堇属植物潜在适生区的变化,运用SDMTools工具箱,计算不同时期紫堇属潜在适生区的几何中心的变化,得出质心迁移的状况,从而得知紫堇属植物在不同时期的潜在适生区的变化趋势[44]。
1.5 紫堇属植物保护现状与保护关键区
从中科院资源环境数据中心(https://www.resdc.cn/data.aspx?DATAID=272)获取中国自然保护区分布数据。通过ArcGIS10.6的Spatial Analyst中的掩膜工具,对不同时期的分布数据进行了掩膜提取,并统计面积,从而得知不同时期紫堇属植物的潜在适生区在保护区内的面积及变化。
2 结果
2.1 模型优化及精度评估结果
根据模型优化结果,FC与RM交叉验证了1240个模型参数组合,其中符合Omission rates<5%且delta AICc较小的模型参数组合共有1个,即 FC选择LQPT、RM为1.4。此时的Omission rates为4.8%且delta AICc较小,表明此参数组合预测结果较好(图3)[45]。本模型得到的平均测试AUC值为0.793,表明模型预测能力较好[40]。

图3 最优模型参数组合选择图Fig.3 Optimal model parameter combination selectiondelta _AICc值: 校准后的最优模型与当前模型的赤池信息量准则的差值 The difference between the calibrated optimal model and the Akaike information criterion of the current model
2.2 影响紫堇属物种分布格局的主要环境因子
本研究通过MaxEnt模型的输出结果并基于刀切法(图4)的正则化训练增益贡献率(Regularized training gain)、测试增益率(Test gain)、置换贡献率(Contribution rate of substitution)以及单因子响应曲线(图5)揭示了影响紫堇属植物地理分布的主导环境因子,从表2的贡献率和置换贡献率可以看出,气温年较差(bio7)、气温季节性变动系数(bio4)、年降水量(bio12)、平均气温日较差(bio2)的贡献率分别为30.5%、29%、20.8%,5.8%,累计贡献率达90.4%;置换重要性分别为2.6%、4%、11.2%、2.8%累计达20.6%,表明以上4个环境因子是影响紫堇属植物分布的主导环境因子。

图5 主导环境因子响应曲线Fig.5 Response curve of dominant environmental factors
2.3 当代气候下紫堇属潜在适生区
由图6可知,紫堇属植物分布的经纬度范围为3°51′-53°33′N、73°33′-135°05′E,分布范围几乎涵盖整个中国。在气候分区上,从北亚热带、南亚热带、中温带、南温带、高原气候区均有分布,其中西北端的中温带及东北端的北温带地区也有少量分布。紫堇属植物的总潜在适生区面积占549.99×104km2。高适生区主要在我国的高原气候区及中亚热带地区,高适生区面积占152.11×104km2,占总潜在适生区27.66%;中适生区主要集中在南亚热带、中亚热带及北亚热带地区,中适生区面积占191.89×104km2,占总潜在适生区34.89%;低适生区主要集中在高原气候区、南温带及中温带,低适生区面积占205.99×104km2,占总潜在适生区37.45%。

图6 当代气候环境下紫堇属适宜区分布Fig.6 The distribution of suitable area of Corydalis in contemporary climate environment
2.4 过去及未来紫堇属潜在适生区模拟
从图7和表3可以看出,LGM时期,紫堇属植物的总适生区面积涵盖了我国的东南部和西南部的绝大部分地区。总适生区涵盖了我国的从北亚热带地区到南温带地区,面积占395.17×104km2;其中高适生区主要分布在我国亚热带地区,面积占146.43×104km2,占总适生区面积的37.06%。
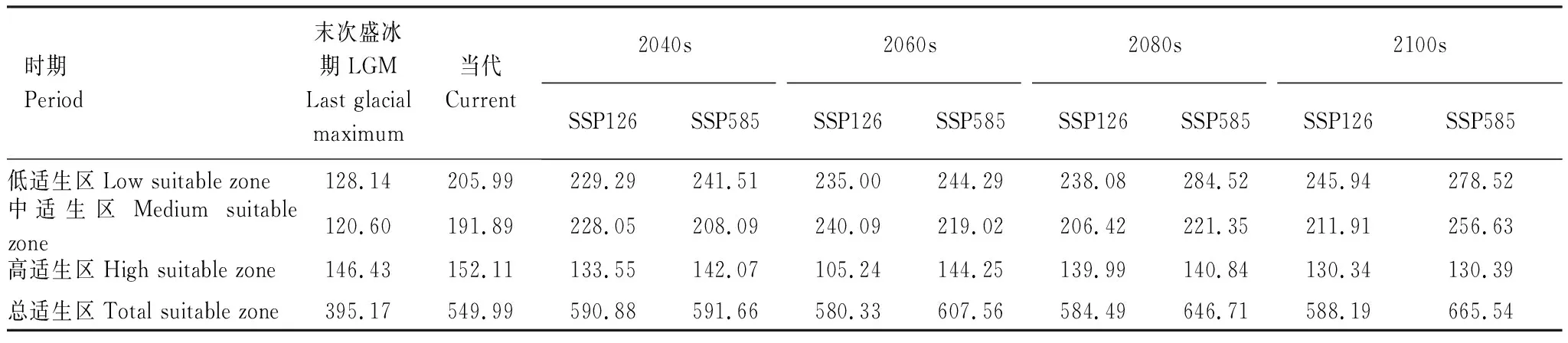
表3 不同时期紫堇属植物潜在适生区面积/(×104 km2)Table 3 Suitable area of Corydalis in different periods

图7 不同时期预测紫堇属植物潜在适生区面积Fig.7 Predict the suitable area of Corydalis plants in different periodsLGM:末次盛冰期Last glacial maximum;SSP:共享社会经济路径Shared Socioeconomic Pathways;SSP126:低浓度温室气体排放场景Low concentration greenhouse gas emission scenario;SSP585:高浓度温室气体排放场景High concentration greenhouse gas emission scenario
在未来时期两种温室气体排放场景(SSP126和SSP585)下,中国大部分地区仍然适宜紫堇属植物的分布,主要分布范围依然是我国的南亚热带、北亚热带、南温带和中温带地区。但是,自LGM到未来时期,在高原气候区内所占面积呈现增加趋势。高适生区自LGM到未来时期的分布变化较大,从LGM时期的主要分布在中亚热带以及北亚热带地区逐渐转为向高原气候区扩张,且破碎化逐渐严重。不同时期的气候变化对位于台湾省的南亚热带及北亚热带地区的紫堇属植物的潜在适生区影响较小,高适生区占比均较大。
2.5 紫堇属潜在适生区空间格局变化
由图8和表4可知,LGM至Current,紫堇属植物的潜在适生区呈现向高原气候区扩张的趋势,总适生区扩张面积为61.41×104km2,总扩张比例为15.54%,为本文所有时期中扩张比例最大的。扩张区域集中在我国的高原气候区,扩张面积约为70.55×104km2,占原潜在适生区的17.85%。
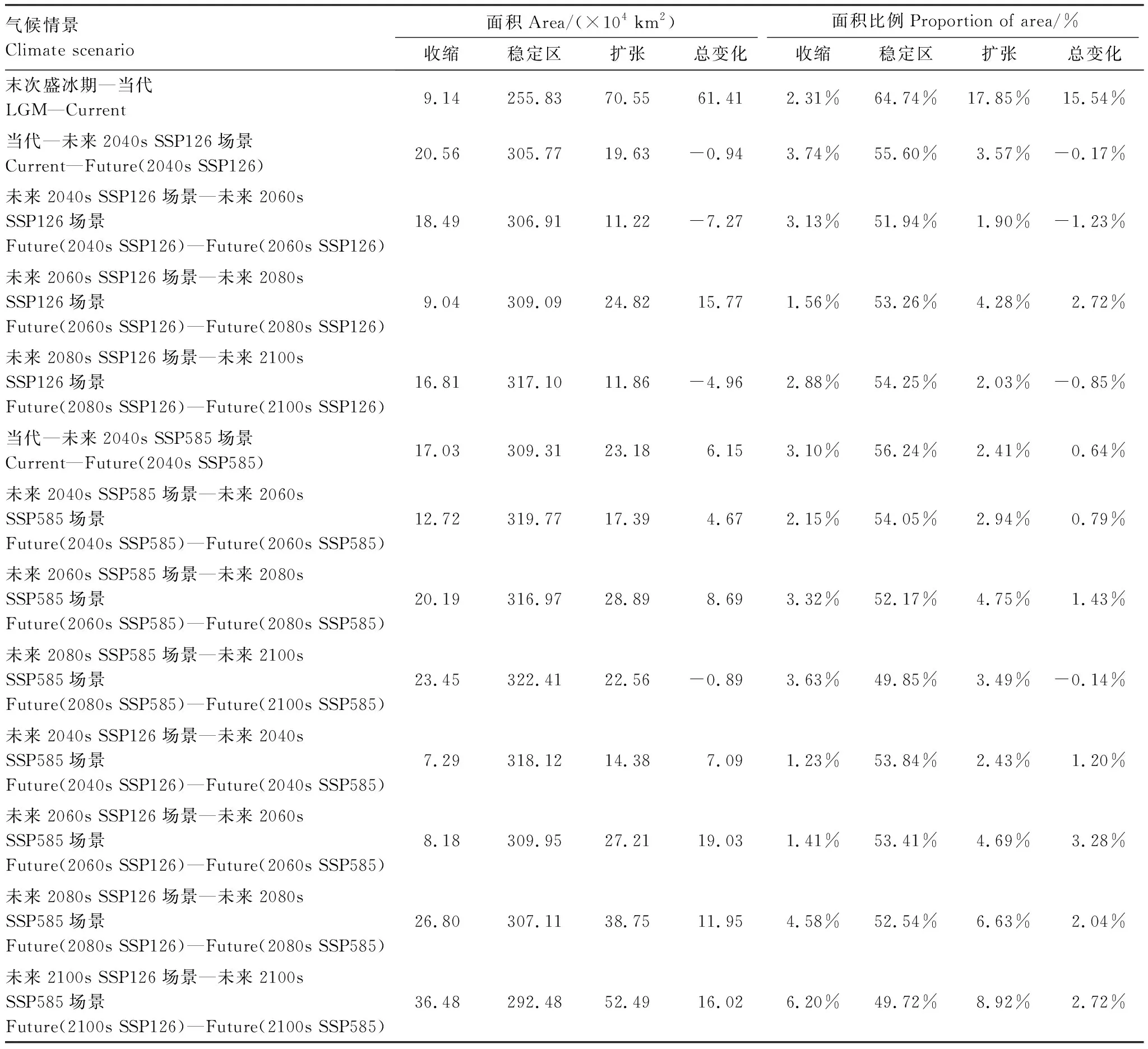
表4 不同时期紫堇属植物潜在适生区变化面积/(×104 km2)Table 4 The change area of suitable area of Corydalis in different periods

图8 不同时期紫堇属植物潜在适生区面积变化Fig.8 Changes of suitable area of Corydalis in different periodsCurrent:当代current;LGM:末次盛冰期Last glacial maximum;SSP:共享社会经济路径Shared Socioeconomic Pathways;SSP126:低浓度温室气体排放场景Low concentration greenhouse gas emission scenario;SSP585:高浓度温室气体排放场景High concentration greenhouse gas emission scenario
在未来SSP126场景下,Current至2040s,紫堇属植物的潜在适生区整体呈现收缩的趋势,总适生区收缩面积为0.94×104km2,总收缩比例为0.17%,收缩区域主要集中在我国的北亚热带和中亚热带地区,收缩面积为20.56×104km2,收缩面积占原潜在适生区的3.74%; 2040s至2060s,紫堇属植物的潜在适生区整体呈现收缩趋势,总适生区收缩面积为7.27×104km2,总收缩比例为1.23%,收缩区域主要集中在我国的在南温带及北亚热带地区,收缩面积为18.49×104km2,收缩面积占原潜在适生区的3.13%;2060s至2080s,紫堇属植物的潜在适生区整体反而呈现扩张的趋势,总适生区扩张面积为15.77×104km2,总扩张比例为2.72%。扩张区域主要集中在我国南亚热带、北亚热带以及南温带东部,扩张面积约为24.82×104km2,扩张面积占原潜在适生区的4.28%;2080s至2100s,紫堇属植物的潜在适生区在我国整体呈现收缩趋势,总适生区收缩面积约为4.96×104km2,总收缩比例为0.85%。收缩区域主要集中在我国的中亚热带和北亚热带的东部,收缩面积约为16.81×104km2,占原潜在适生区的2.88%。
在未来SSP585场景下,Current至2040s,紫堇属植物的潜在适生区在我国整体呈现向西北扩张的趋势,总适生区扩张面积为6.15×104km2,总扩张比例为6.15%,扩张区域主要集中在我国的高原气候区及中温带的东部地区,扩张面积约为17.03×104km2,扩张比例为3.10%;2040s至2060s,紫堇属植物的潜在适生区在我国整体反而呈现扩张趋势,总适生区扩张面积为4.67×104km2,总扩张比例为0.79%。扩张区域主要集中在我国的中温带的东北部地区,扩张面积为12.72×104km2,占原潜在适生区的2.15%;2060s至2080s,紫堇属植物潜在适生区在我国整体呈现继续向北扩张趋势,总适生区扩张面积为8.69×104km2,总扩张比例为1.43%。扩张区域主要向我国的高原气候区西北方向扩张,扩张面积为20.19×104km2,占原潜在适生区的3.32%;2080s至2100s,紫堇属植物的潜在适生区在我国整体呈现了收缩趋势,总适生区收缩面积为0.89×104km2,总收缩比例为0.14%。收缩区域主要集中在我国的南亚热带、北亚热带和中温带的一部分地区,收缩面积为23.45×104km2,占原潜在适生区的3.63%。
本研究还对比了在未来两种不同温室气体排放场景下,紫堇属植物的潜在适生区的变化,结果表明,在模拟高浓度温室气体排放场景下,高浓度温室气体的排放均使紫堇属植物的潜在适生区发生扩张,且扩张区域整体向北。具体来看:在2040s SSP126至2040s SSP585情况下,紫堇属植物的潜在适生区在我国的北亚热带和南亚热带呈现扩张的趋势,总适生区扩张面积为7.09×104km2,总扩张比例为1.20%;在2060s SSP126至2060s SSP585情况下,紫堇属植物的潜在适生区在我国的中亚热带、中温带、高原气候区均有扩张,总适生区扩张面积约为19.03×104km2,总扩张比例为3.28%;在2080s SSP126至2080s SSP585情况下,紫堇属植物的潜在适生区在我国的高原气候区出现了较大面积的扩张,总适生区扩张面积为11.95×104km2,总扩张比例为2.04%;在2100s SSP126至2100s SSP585情况下,紫堇属植物的潜在适生区在我国的高原气候区与中温带地区均有大面积的扩张,总适生区扩张面积为16.02×104km2,总扩张比例为2.72%。
2.6 潜在适生区质心的转移
本研究将紫堇属的潜在适生区以几何中心点来定义,以此方式模拟不同气候情景下质心迁移的变化。从表5和图9可以看出,Current的质心的位置相较于LGM的质心向西北方向发生迁移,迁移距离为198.06 km。在未来SSP126场景下,在2040s时的质心相较于Current时的质心继续向西北方向迁移,迁移距离为83.53 km;在2060s年时的质心则较2040s时的质心继续向西北方向迁移,迁移距离为27.53 km;2080s时的质心则较2060s时的质心向东南方迁移,迁移距离为48.23km;在2100s时的质心相较于2080s时的质心又向西北方迁移,迁移距离为45.70 km。

表5 不同时期紫堇属分布质心经纬度Table 5 The latitude and longitude of the distribution center of Corydalis in different periods
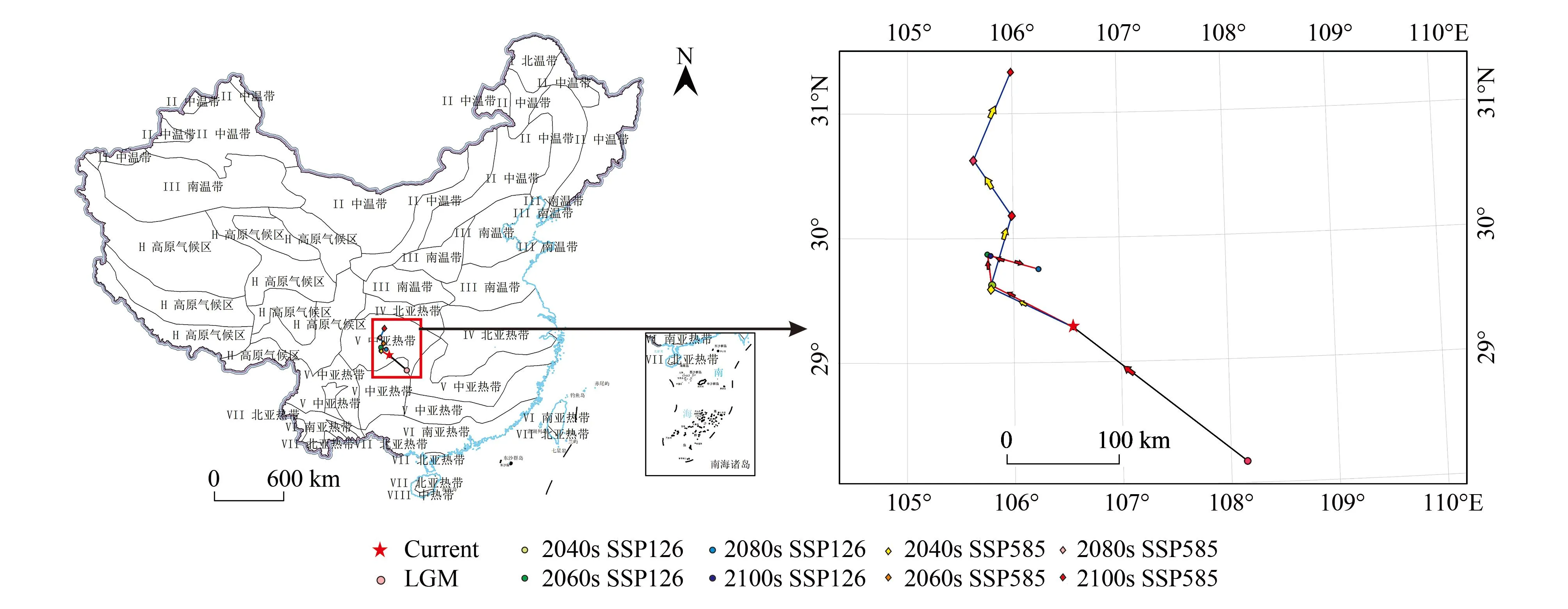
图9 不同时期紫堇属潜在适生区质心变化Fig.9 Centroid changes of Corydalis in different periodsCurrent:当代current;LGM:末次盛冰期Last glacial maximum;SSP:共享社会经济路径Shared Socioeconomic Pathways;SSP126:低浓度温室气体排放场景Low concentration greenhouse gas emission scenario;SSP585:高浓度温室气体排放场景High concentration greenhouse gas emission scenario
在未来SSP585场景下,2040s时的质心较Current时的质心向西北方向迁移,迁移距离为101.52 km;2060s时的质心相较于2040s时的质心向东北方继续迁移,迁移距离为66.93 km;2080s时的质心相较于2060s时的质心继续向西北方迁移,迁移距离为60.61 km;2100s时的质心相较于2080s时的质心则向东北方迁移,迁移距离为86.32km。
从LGM至Current时的质心总体从我国的东南部逐渐向我国的西北方向迁移。在未来SSP126场景下,质心先不断向西北方向移动,在2080s时突然向东南方向移动,到2100s时又向西北突然折返;在SSP585场景下,质心总体一直不断向西北方向迁移,不论LGM至未来时期如何变化,质心依然位于我国的中亚热带气候区。
2.7 紫堇属植物保护现状
由图10可知,中国面积最大的自然保护区主要集中在西北地区。而紫堇属植物在当代及未来不同时期的主要潜在适生区仍然分布于中国的东南部及西南部地区,且高适生区主要集中我国西南部地区,而此处的自然保护区却呈现面积较小,且破碎严重的特点。因此在我国的西南部地区对紫堇属植物的保护尤为关键。由图11可知,在未来模拟温室气体不断增多的情景里,无论在模拟高浓度温室气体排放场景还是在低浓度温室气体排放场景下,高适生区随着温室气体浓度的增加位于保护区内的面积变化较小。但随着温室气体浓度的增加,位于我国西北部保护区内的潜在适生区的面积呈现出不断增加的趋势。

图10 不同时期紫堇属植物保护区内潜在适生区分布Fig.10 Distribution of suitable areas in protected areas of Corydalis plants in different periods

图11 不同时期紫堇属植物保护区内高潜在适生区与总潜在适生区所占面积Fig.11 The area of high suitable area and total suitable area in different periods of Corydalis plant protection area
3 讨论
3.1 影响紫堇属植物分布的主要环境因子
Maxent模型具有用较少样本量便能对物种的分布进行预测的特点[46],广泛应用于生态学中的适宜性评价等领域[26]。本研究根据优化后的MaxEnt模型所得出的结果中的训练增益值、贡献率以及单因子响应曲线,揭示影响紫堇属植物分布的主要环境因子为气温年较差(bio7)、气温季节性变动系数(bio4)、年降水量(bio12)、平均气温日较差(bio2)。由此可见,温度是影响紫堇属植物分布重要的环境因子。温度条件通常对植物的分布起着非常重要的作用[13]。杨冬璠等人在对尼泊尔黄堇的潜在适生区及主要驱动因素进行研究中发现最暖月的最高温和等温性对尼泊尔黄堇的潜在适生区具有显著的影响[47],表明温度的相关变量对紫堇属植物分布的影响显著,与本研究所得的结果相吻合。
高山低温环境对高山植物的影响尤为显著[48],紫堇属植物是主要分布于横断山脉与东喜马拉雅地区的植物类群[18],其胚胎需要在低温下培育8周以上才能开始发育[49],对低温环境展现出了广泛的适应性,这些生理特征大大提高了紫堇属植物在高寒环境下的适应能力。另外,横断山脉与东喜马拉雅地区因受东亚和南亚季风的影响而湿度较大[48],降水量也会影响紫堇属植物叶片的N、P和地上生物量[50],因此,降水量也会与温度一起参与对紫堇属植物分布的影响。
3.2 紫堇属植物潜在适生区的分布、变化以及质心转移分析
紫堇属植物的适生区在Current时期的分布几乎涵盖中国的大部分地区,在气候分区上主要集中在北亚热带、南亚热带、中温带、南温带和高原气候区,与实际所搜集到的紫堇属植物分布点坐标基本一致,表明模型模拟效果良好。
LGM时期,紫堇属植物的潜在适生区仍主要集中在我国亚热带气候区,从LGM至Current,紫堇属植物的潜在适生区呈现向高原气候区扩张的趋势,与王晓帆等对东喜马拉雅地区的高山栎组植物[12]的研究相反,可能是由于温室气体浓度的上升使得气候不断变暖,高原气候区的环境更加的温暖湿润,使原本主要位于亚热带地区的紫堇属植物不断向西北方向的高原气候区扩张。本文也发现气候变化对紫堇属植物在台湾省的潜在适生区影响较小,与Runan Zhao等人对天台鹅耳枥(Carpinustientaiensis)的潜在适生区模拟的结论相悖[51],可能是由于紫堇属植物具有较广的海拔分布范围(0-6000m)[15],而台湾省中央山脉从东北向西南贯穿整个省,丘陵和山脉成为台湾省的主要的地形地貌。高山等复杂的地形地貌为紫堇属植物创造了多样化的生境,生境异质性使紫堇属植物具有较高的物种多样性,也为紫堇属植物在气候变化过程中提供了避难所[17],从而使其影响较小。
整体来看,从LGM至未来时期的紫堇属潜在适生区呈现出破碎化逐渐严重的趋势,与王晓帆对高山栎组的研究结果相似[12],这可能是由于CO2排放增加,过度放牧、开荒等人类活动因素加剧了生境的破碎化[52]。在未来SSP126低浓度温室气体排放场景下,紫堇属植物潜在适生区除了在2080s呈现了扩张趋势,均呈现收缩趋势,与张殷波对山西翅果油树的研究结果[43]类似,且收缩气候区主要集中在我国的北亚热带与南亚热带地区,由于温室气体浓度的增加,使位于我国东部的北亚热带与南亚热带气候区变得更加的炎热,使原本的紫堇属植物的潜在适生区内的水热条件发生变化,且极端气候频发,加剧了紫堇属植物潜在适生区的收缩。
在未来SSP585高浓度温室气体排放场景下,从Current至未来2100s,潜在适生区均呈现扩张趋势,主要向我国的北方扩张,扩张区域主要集中在我国的高原气候区以及中温带地区,与陈美霖对黄土高原地区的4种常见树种适生区随气候变化[53]的研究结果相似,在高浓度排放场景下,在我国的高原气候区及中温带地区原本不适宜紫堇属植物生长的地区,由于气候变暖改变了当地的生境,使其转变成适宜紫堇属植物生长的生境,从而促使紫堇属植物不断向我国的中温带以及高原气候区扩张。
质心迁移结果表明不同时期的质心迁移均在中亚热带地区,在SSP126低浓度排放场景场景下质心呈现西北-西北-东南-西北的迁移,而在SSP585高浓度排放场景下,质心则向西北-东北-西北-东北方向迁移,整体仍主要向西北方向迁移。相关研究表明,在气候变暖背景下,植物分布具有向高纬度高海拔地区扩张的趋势[6],与本文研究的结果相吻合。
在SSP126低浓度温室气体排放场景下,适生区的质心迁移出现了折返的现象,质心迁移发生折返的地区位于四川省与重庆市交界的川东平行岭谷地区,该地区是一系列东北西南走向的平形状山岭和谷地,是典型的褶皱山区[54]。复杂的地形地貌为在低浓度温室气体排放场景(SSP126)下的紫堇属植物提供了避难所[55],同时也阻碍了其向更高海拔和更高纬度地区迁移。在SSP585场景下,气候变暖使高纬度高海拔地区更加温暖湿润[21],为其向更高海拔更高纬度地区的迁移提供了可能,因此在SSP585高浓度温室气体排放场景下,紫堇属植物的潜在适生区的质心不断向西北方的高海拔高纬度地区迁移。从整体来看,紫堇属植物的适生区的破碎化逐渐严重,不论是低浓度排放场景下的潜在适生区不断收缩,还是在高浓度排放场景下的潜在适生区的扩张,气候变化都对紫堇属植物的生境造成了巨大的影响,气候变化严重威胁着紫堇属植物的生存。
根据现有的自然保护区分析来看,我国西南地区在气候变化下各个时期中均为紫堇属植物分布的集中区,因此,我国西南地区为紫堇属物种多样性保护的关键区域。但我国现有的自然保护区面积最大区域主要集中在西北地区,在西南地区的保护区则呈现出破碎且保护区面积较小的特点,因此应加大我国西南部的自然保护区的面积且增加自然保护区之间的连通性,以更好地保护紫堇属植物的物种多样性。在未来气候变化下,紫堇属植物呈现向西北方向迁移的趋势,现有的保护区在我国西北部具有较大的分布面积,在未来气候下更多紫堇属植物将受到保护。本研究建议应继续增加我国在西北部高原气候区自然保护区的面积以适应未来紫堇属植物潜在适生区的变化。
4 结论
本研究基于Kuenm包优化的MaxEnt模型对紫堇属植物在过去LGM时期,Current以及两种温室气体排放模式(SSP126和SSP585)下未来4个时期的潜在适生区以进行了模拟。
(1)紫堇属植物在Current气候条件下,其潜在适生区从北亚热带、南亚热带、中温带、南温带和高原气候区均有分布。
(2)气温年较差(bio7)、气温季节性变动系数(bio4)、年降水量(bio12)、平均气温日较差(bio2)是影响紫堇属植物分布的主导环境因子,可看出温度是影响紫堇属植物分布重要的环境因子。
(3)LGM时期的紫堇属潜在适生区主要分布在我国的北亚热带到南温带地区,而Current时期紫堇属植物则大面积向高原气候区扩张,未来在SSP126低浓度温室气体排放场景下,紫堇属植物潜在适生区主要呈现收缩的趋势,收缩区域主要集中在我国南亚热带与北亚热带地区,未来SSP585高浓度温室气体排放场景下,紫堇属潜在适生区呈现扩张的趋势,扩张区域主要集中在我国高原气候区与东北部的中温带地区。从质心迁移来看,紫堇属潜在适生区的质心在SSP126低浓度温室气体排放场景下,呈现西北-西北-东南-西南的迁移,在SSP585高浓度排放场景下,质心则向西北-东北-西北-东北方向迁移,整体仍主要向西北方向迁移,且紫堇属植物潜在适生区的质心始终在中亚热带地区。整体来看,紫堇属植物的潜在适生区从过去到未来呈现出不断向高纬度高海拔的高原气候区,中温带地区移动的趋势,且其高适生区出现了破碎化逐渐严重的现象,气候变化对紫堇属植物造成了严重的威胁。
(4)从紫堇属植物的保护现状来看,紫堇属植物在气候变化下的不同时期,我国西南东喜马拉雅地区与横断山脉地区依然为紫堇属植物的主要分布地区,为保护的关键区,然而实际分布于我国西南地区的自然保护区呈现面积小和破碎化严重的特点,极不利于紫堇属物种的保护。在未来时期,随着气候变化,紫堇属植物将会向保护区分布面积较大的西北地区迁移,从而可获得更多的保护,但是对于我国西南部的保护区建设依然极为重要且关键。
