若尔盖不同生境植物多样性与植物竞争强度和生态位重叠度的关系
2024-01-17吴凯婷马燕丹郑秋竹岳海涛王晓蓉梁克敏
吴凯婷, 张 勇,*,马燕丹,郑秋竹,岳海涛,王晓蓉,梁克敏,曾 昊
1 西南林业大学 国家高原湿地研究中心,昆明 650224 2 西南林业大学 云南省高原湿地保护修复与生态服务重点实验室,昆明 650224 3 华中农业大学农业微生物学国家重点实验室,武汉 430070 4 华中农业大学资源与环境学院,武汉 430070
物种多样性是生物多样性在物种水平上的表现形式[1]。一定空间范围内物种的丰富度和均匀度体现了群落中物种的数量及其分布特征,是反映群落物种多样性的重要指标[2],也是群落功能复杂性与稳定性的重要度量指标[3]。
物种多样性的形成和维持机制是理论生态学的核心研究议题[4-6]。竞争现象在生物群落中普遍存在,它为研究物种生态位分化以及物种多样性维持提供了钥匙[7-10]。基于物种竞争、物种生态位重叠和物种多样性的关系,目前主要通过生态位分化理论解释物种多样性维持机制。在生态位分化理论中,竞争排斥法则表明生态位相同的物种不可能长期稳定共存[11]。基于控制实验和模型模拟的研究表明,物种稳定共存于同一个生境的前提条件是它们出现了生态位分化,物种生态位分化越明显,它们的生态位重叠度越低,对资源的竞争强度得到缓解[12-13]。如果两个物种间的生态位重叠度和竞争强度增加,往往意味着竞争排斥[14]。但也有控制实验表明,改善微环境可以塑造出更多生态位使物种共存,植物发生竞争排斥现象的概率很低[15]。
但在实际情况中(真实植物群落往往包含多个物种),植物物种竞争强度与生态位重叠度的关系较复杂,常表现出无关、正相关等形式。例如,对大兴安岭次生林不同龄级落叶松(Larixgmelinii)的研究发现,落叶松个体间的生态位重叠度与彼此间的竞争作用无显著相关性[16]。对香合欢群落(Ass.Albiziaodoratissima)的研究则表明,群落内物种生态位重叠度与物种竞争强度呈正相关[17]。生境异质性是导致这种差异的重要原因。此外,对植物生态位重叠度和竞争强度的计算方式不一致也是导致它们之间关系不统一的原因[18]。目前大多数研究通过Pianka生态位重叠指数量化群落水平的植物生态位重叠度[19],用Lotka-Volterra竞争模型计算两个物种之间的竞争强度[20-21],但难以在群落水平定量描述植物的竞争强度。
若尔盖地区是我国面积最大的典型泥炭沼泽分布区[22],其对涵养水源、调节气候、维持生物多样性等具有十分重要的作用[23-24]。近年来,在气候变化和人为干扰的共同影响下,若尔盖地区的湿地生态系统退化严重,沼泽植物群落的生境逐渐由水生向中生转变,在沼泽周边形成了明显的水生、湿生、湿生-中生以及中生生境[25-27]。这种生境分布特征为研究物种多样性与物种竞争、物种生态位重叠之间的关系提供了理想的环境梯度。目前,若尔盖地区的植物多样性研究主要集中在植物多样性分布与环境因子的关系[28-30]、排水梯度上植物种间相关性特征[31]以及退化梯度上草地植物生态位重叠度变化特征[32]等方面,对植物多样性与植物竞争、生态位重叠关系的研究不足。因此,本研究在若尔盖县选取典型的生境梯度,开展植物群落调查,基于群落调查数据构建植物竞争强度指数(Competition intensity,CI),进而探讨植物多样性与CI和生态位重叠度的关系。依据竞争排斥原则,本研究拟验证以下假设:在群落水平上,若尔盖地区植物CI和物种生态位重叠度呈正相关关系,该地区的植物多样性指标与植物CI和物种生态位重叠度呈负相关关系。
1 材料与方法
1.1 研究区概况及野外调查
若尔盖县位于四川省阿坝藏族羌族自治州,平均海拔为3400-3700 m,属高原亚寒带湿润气候,常年无夏,日照时间长,相对湿度高,年均温为0.6-1.0℃,年降水量为650-750 mm[26]。其主要植被类型是沼泽化草甸和高寒草甸,主要优势种有木里薹草(Carexmuliensis)、藏嵩草(Kobresiatibetica),线叶嵩草(K.capillifolia)等[27]。近年来由于气候变化以及人为开渠排水等因素,研究区沼泽水位下降,植物群落类型发生明显变化。
植物群落往往具有特定的种类组成及分布范围,并指示不同的生境条件[33]。从水生到中生转变的过程中,若尔盖地区植被类型由沼泽植被逐渐过度成高寒草甸[29]。根据植物群落的优势种及伴生种构成情况,将研究区的植物群落生境依次划分为水生、湿生、湿生-中生和中生4个类型(表1)。于2021年8月植物生长季在若尔盖县的典型沼泽分布区(图1),根据生境梯度,分别在卡哈尔乔、昂当乔和那勒乔设置9个、6个和4个10 m×10 m的样地(共19个样地,相邻样地之间平均间距约220 m,最大间距约860 m,最小间距约40 m,各样地在生境梯度上的分布情况见表1),在每个样地中随机设置3个1 m×1 m的调查样方,共计57个样方。在样方内进行植物群落调查,记录样方内植物群落的垂直投影盖度、平均高度、植物名称、分种盖度和分种高度。

图1 研究区样地及样方分布示意图Fig.1 Location of study area and the distribution of sampling sitesAD0-AD4:昂当乔样地 Sampling sites in Angdangqiao;NL1-NL 4:那勒乔样地 Sampling sites in Naleqiao;KH1-KH9:喀哈尔乔样地 Sampling sites in Kahaerqiao
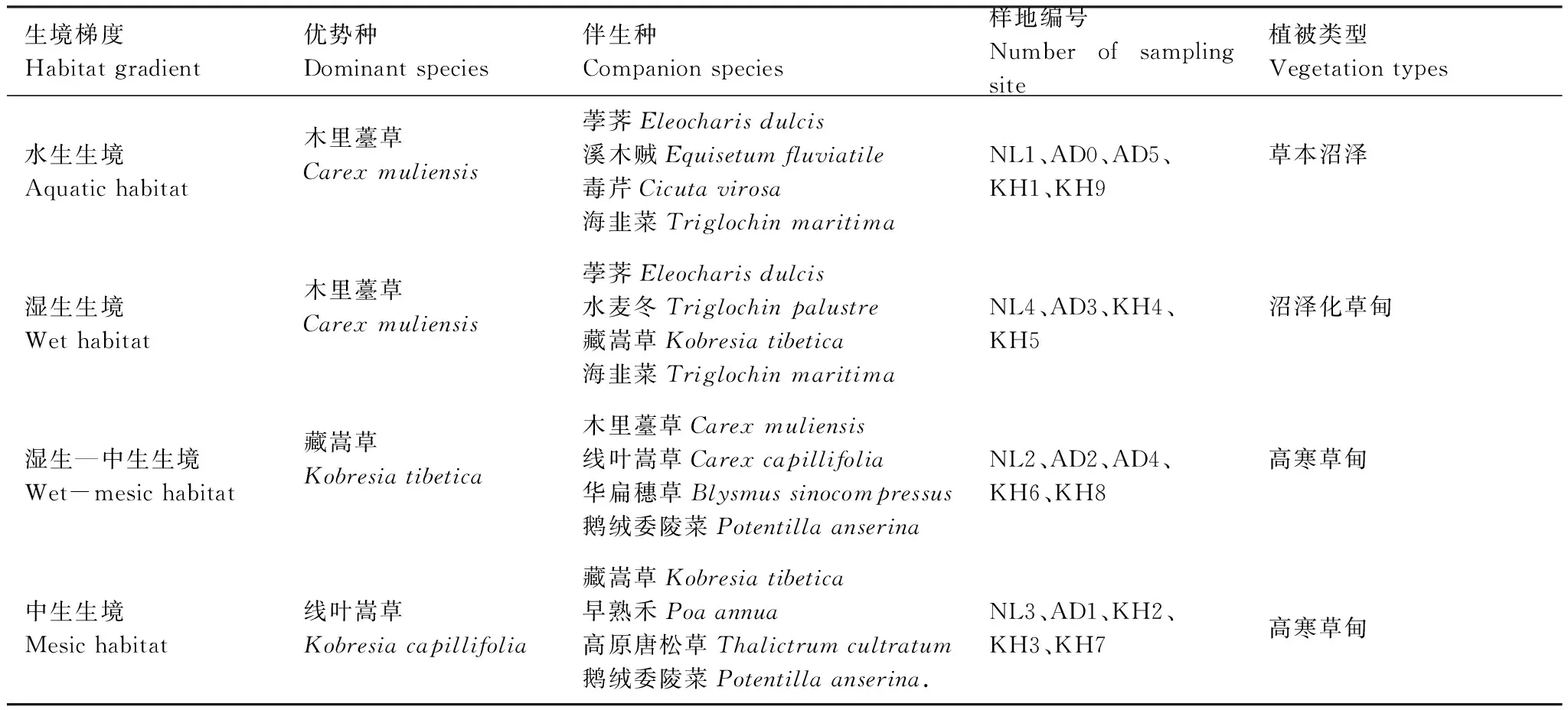
表1 研究区生境梯度上植物群落构成情况Table 1 Plant community composition across the habitat gradient in the study area
1.2 指数计算
(1)物种多样性指数计算
本研究采用了常用的物种多样性指数,分别为物种丰富度指数(R0)、Simpson指数(D)、Shannon-Weiner指数(H′)和Pielou均匀度指数(J),它们的计算公式[34-36]如下:
R0=S
Pi=Ni/N
J=H′/lnS
式中,S为物种数,Pi为种i的个体在全部个体中的比例,Ni为种i的个体数,N为群落中全部物种的个体数。
(2)竞争强度指数(CI)计算
本研究通过植物群落物种实际生存空间和群落潜在生存空间的占比表示植物竞争强度(图2),CI计算公式为:

图2 植物竞争强度计算示意图Fig.2 Schematic diagram for the calculation of plant competition intensity
式中,CI为植物竞争强度,Lsi为物种i的实际生存空间,用物种i的盖度Ci(单位:m2)与实际高度Hi(单位:m)计算得到,即Lsi=Ci×Hi;Lsp为潜在生存空间,用样地面积(即1 m2)与梯度内植物潜在高度Hp(用植物高度中位数表示,单位为m)相乘得到,即Lsp=1×Hp。
(3)物种生态位重叠度(NOS)计算
本研究基于物种重要值(Important value,IV)计算NOS,其中IV的计算方式为:
IV=(RH+RC)/2

式中,RH为植物相对高度,RC为植物相对盖度,Hi为物种i的高度(m),Ci为物种i的盖度(%)。
基于IV,采用Pianka生态位重叠度指数计算计算每个物种的生态位重叠度,其计算公式为[37-38]:
式中,Qik为物种i与物种k的重叠度指数,pij和pkj为种i和种k对第j个资源的利用占它对全部资源利用的频率,r代表样方数。植物群落的物种生态位重叠度(即NOS)由群落中全部物种的生态位重叠度求均值得到。生态位重叠度计算在R4.1.3中完成。
1.3 统计分析
采用单因素方差分析(One-way ANOVA)对比物种多样性指数(即物种丰富度指数、Simpson指数、Shannon-Weiner指数和Pielou均匀度指数)、CI和NOS在生境梯度间的差异性。采用回归分析探讨物种多样性指数与CI和NOS之间的关系。上述统计分析在R4.1.3中完成。
2 结果与分析
2.1 生境梯度上植物多样性指数的变化特征
从水生到中生生境,植物群落的四个物种多样性指数呈现增加的趋势(图3)。生境梯度上植物群落物种丰富度的排序为湿生-中生((19.9±1.2)种)>中生((16.7±0.7)种)>湿生((10.3±1.2)种)>水生((5.7±0.7)种)。湿生、湿生-中生、中生生境中植物群落的Simpson指数分别为0.68±0.02、0.75±0.03和0.78±0.02,它们显著高于水生生境植物群落的0.47±0.05。湿生-中生和中生生境中,植物群落的Shannon-Weiner指数分别为2.9±0.2和2.8±0.1,它们显著高于水生(1.4±0.2)和湿生(2.2±0.1)植物群落。湿生、湿生-中生和中生生境植物群落的Pielou均匀度指数分别为0.97±0.04、0.96±0.04和1.10±0.03,它们显著高于水生植物群落的0.82±0.05。

图3 植物群落物种多样性指数在生境梯度间的差异性Fig.3 Changes in plant diversity indices across the habitat gradient图中不同小写字母表示显著差异
2.2 生境梯度上植物CI和NOS的变化特征
湿生-中生生境中,植物群落的CI为1.77±0.14,其显著高于湿生植物群落的1.15±0.16(P<0.05);水生和中生植物群落的CI与湿生-中生和湿生植物群落无显著差异(P>0.05)。湿生植物群落的NOS为0.6±0.04,其显著高于水生植物群落的0.48±0.03(P<0.05);湿生-中生和中生植物群落的NOS与湿生和水生植物群落无显著差异(P>0.05)。整体来看,植物群落的CI与NOS无显著相关性(P>0.05)。在湿生植物群落中,CI和NOS呈倒抛物线关系;当CI低于1.31时,NOS随CI增加而下降;当CI高于1.31时,NOS随CI增加而上升(图4)。
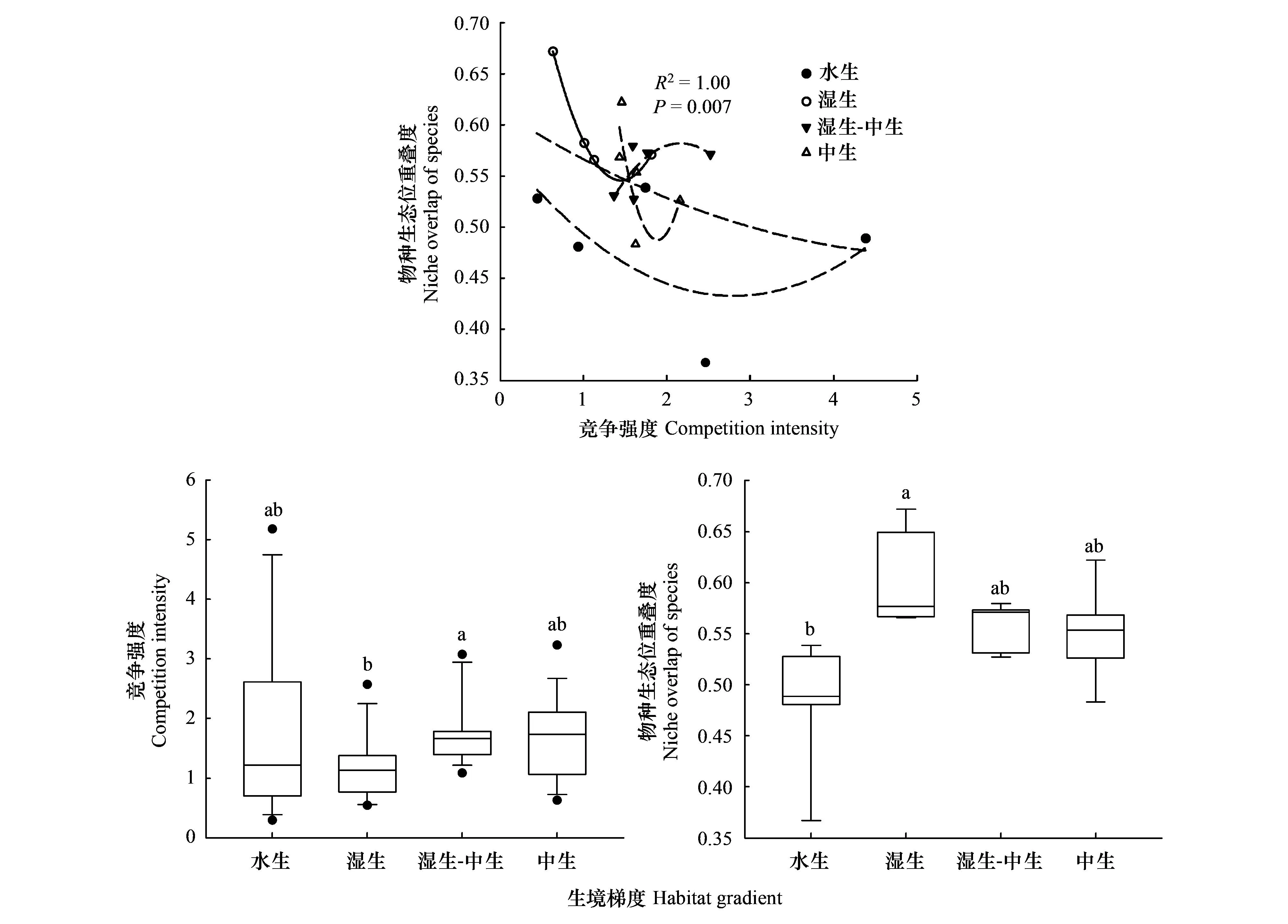
图4 植物竞争强度和生态位重叠度在生境梯度间的差异及相关性Fig.4 Changes in competition intensity, niche overlap and their relationships across the habitat gradient图中不同小写字母表示显著差异
2.3 生境梯度上植物多样性与CI和NOS的关系
整体来看,植物群落的物种丰富度与CI呈抛物线关系;当CI小于2.41时,物种丰富度随CI增加而上升;当CI大于2.41时,物种丰富度随CI增加而下降(P<0.05)。植物群落的Shannon-Weiner指数、Simpson指数和Pielou均匀度指数与CI无显著相关性(P>0.05) (图5)。植物群落的Simpson指数、Shannon-Weiner指数和Pielou均匀度指数均与NOS呈显著正相关关系(P<0.05),但植物物种丰富度指数与NOS之间的正相关关系未达到统计显著性(P>0.05)(图6)。

图5 生境梯度上植物群落物种多样性与竞争强度的关系Fig.5 Relationships between plant diversity and plant competition intensity across the habitat gradient

图6 生境梯度上植物群落物种多样性与物种生态位重叠度的关系Fig.6 Relationship between plant diversity and niche overlap of species across the habitat gradient
从各生境看,所有生境的物种丰富度与CI无显著相关性(P>0.05);湿生-中生植物群落的Simpson指数、Shannon-Weiner指数和Pielou均匀度指数均与CI呈显著负相关关系(P<0.05),但水生、湿生和中生植物群落的上述植物多样性指数与CI无显著相关性(P>0.05)(图5)。所有生境中,四个植物多样性指数均与NOS无显著相关性(P>0.05)(图6)。
3 讨论
系统复杂性常导致生态系统过程难以准确模拟[39]。植物竞争(包括种间竞争和种内竞争)是一个复杂的生态系统过程,因此,当前对植物竞争关系的研究多以控制实验和模型模拟为主。衡量真实群落中的植物竞争强度对解释群落物种共存有重要意义。在生态学研究中,除了细致模拟系统各要素的变化动态及要素间的相互关系外[39],还可以采用“黑箱”或“灰箱”的方式来处理系统复杂性[40-41]。本研究基于潜在生存空间和实际生存空间的概念,将复杂的种内、种间竞争过程当作黑箱处理,只关注竞争的结果:植物群落占据了多少实际生存空间。进而通过实际生存空间和潜在生存空间的占比(即CI)衡量群落水平上的植物竞争强度。这种植物竞争强度计算思路的基本假设与Lotka-Volterra竞争模型一致。L-V竞争模型假设环境中资源是有限的,物种A对资源的使用会限制物种B对资源的使用[42]。本研究中CI计算的假设是植物群落在特定生境中可达到的生存状态(用“潜在生存空间”表示)是一定的,已经形成的生存状态(用“实际生存空间”表示)会限制群落未来的生存状态。在生存压力下,CI值高就意味着植物竞争强度大。
基于上述新算法,本研究得到了群落水平上若尔盖地区典型生境中植物多样性、植物竞争强度和植物生态位重叠度三者之间的关系。
在生态位分化理论框架下,物种生态位重叠度通常与物种对资源的竞争强度呈正相关[43-44]。本研究中,仅在湿生生境中发现植物生态位重叠度与植物竞争之间存在显著的倒抛物线关系,在水生、湿生-中生和中生生境中它们并无显著回归关系。此外,相较于其他生境,湿生生境中的植物竞争强度最低、物种生态位重叠度最高,水生生境中植物竞争强度较高、物种生态位重叠度最低。这种差异性表明,植物生态位重叠度与竞争强度的关系很大程度上受制于局域小生境的影响[28]。在若尔盖地区的沼泽退化的过程中,植物生境逐渐由水生转变为中生,地表积水逐渐减少,土壤环境中的氧气含量增加,土壤营养元素的转化过程(如氮素的转化过程由反硝化过程转变为硝化过程)改变,使得土壤速效养分含量随之增加[45]。土壤速效养分含量的增加改变了植物可使用资源的数量和维度,进而导致植物竞争和生态位重叠的关系发生变化[18]。
植物多样性不仅随全球尺度的环境梯度(如水热格局)发生显著变化[36-37],也随局地小生境发生变化,而竞争和生态位分化对植物多样性形成起到重要作用[46-50]。本研究发现,植物生境由水生到中生的转变过程中,若尔盖地区的植物多样性呈增加趋势,该现象与已有研究结果一致[28-29]。在生境梯度上,植物物种丰富度与植物竞争强度呈抛物线关系,该关系与Lotka-Volterra模型推导的结论不一致,这说明在真实植物群落中一定强度的竞争有利于维持最大的物种数量,其与中度干扰理论揭示的单峰过程类似[51]。
此外,本研究发现生境梯度上植物多样性指数随物种生态位重叠度增加而显著上升。该现象也与经典的生态位分化理论假设不符。这是由于从水生到中生转变的过程中,群落中植物物种数量增加,而在高寒地区植物对土壤速效氮、磷等养分资源具有相似的使用需求[52],因此不可避免地会出现物种生态位重叠度增加的现象。
值得注意的是,生态交错区往往是生物多样性较高的区域[53]。因本研究中湿生-中生生境中的植物多样性最高,且处于湿生向中生过渡的区域,故类比“生态交错区”的概念,将该生境称为“生境交错区”。在本研究中,水生、湿生和中生生境中植物多样性指数与植物竞争强度无显著回归关系,但在湿生-中生生境中,植物群落的Simpson指数、Shannon-Weiner指数和Pielou均匀度指数都与植物竞争强度呈显著线性负相关。从植物多样性与物种生态位重叠度的关系来看,湿生-中生生境中的关系趋势也与其他三个生境相反。这说明若尔盖“生境交错区”的植物多样性与植物竞争和生态位重叠的关系可能与其他单一生境中的不一样。后续研究需继续关注该现象,揭示其背后机理可能有助于理解沼泽退化背景下若尔盖地区植物多样性的维持机制。
4 结论
本研究表明,从水生植物群落到中生植物群落的演变过程中,若尔盖地区的植物多样性呈增加趋势。除湿生生境外,植物竞争强度和生态位重叠度之间无显著回归关系。植物多样性与植物竞争强度和生态位重叠度的关系在生境间的异质性高。上述结果表明本研究的假设不成立,即:在群落水平上,若尔盖地区的植物多样性、植物竞争强度、生态位重叠度三者之间并非简单的线性关系,其会随生境发生变化。特别地,湿生-中生生境中植物多样性与植物竞争强度和生态位重叠度的关系与其他生境不同,解析这种差异的原因有助于理解若尔盖地区植物多样性的维持机制。
