围栏封育对黄河源区斑块化退化高寒草甸碳交换及其组分的影响
2024-01-17李成一李希来杨元武张世彬杨鹏年
李成一,李希来,杨元武,张世彬,杨鹏年
青海大学农牧学院,西宁 810016
在碳元素的生物地球化学循环过程中,净生态系统碳交换(Net ecosystem carbon exchange,NEE)是关键的过程之一,也是全球碳循环的重要组成部分[1-2]。该过程不是单一的碳源通量,而是植物光合(总初级生产力,Gross primary productivity,GPP)增益和生态系统呼吸(Ecosystem respiration,ER)损失的函数差值[3]。它们之间的微小失衡,会导致大气CO2的显著年际变化[4],并可能对气候变化产生反馈[5]。因此,研究陆地生态系统与大气CO2的运移规律及源和汇的评价至关重要。
黄河源区地处青藏高原腹地,是黄河流域生态环境保护和高质量发展重点区域,也是黄河上游主要水源涵养区和产流区[6]。其上广泛分布着高寒草甸,土壤有机碳储量大[7]。加之高寒草甸独特的生物地球化学过程与脆弱的生态环境使之对全球气候变化和人为干扰的响应更为敏感[8-9],一直是生态学研究陆地生态系统碳交换对环境变化响应规律的理想场所。但近年来黄河源区近1/3的草地发生了不同程度的退化,其中严重退化的草地面积占总退化草地面积的26.79%[10-11]。退化是由人类过度放牧引起,啮齿动物高原鼠兔等暴发,致使生境破碎化,形成许多残存草地裸露斑块[12-13],而草地的斑块化削弱了高寒草甸的碳汇功能,加快了生态系统碳排放速率[14],这对黄河源区高寒草甸生态系统的稳定性和生态环境的可持续发展产生了极大的影响。因此,必须采取有效措施来修复和治理退化高寒草甸。
围栏封育作为退化草地恢复与重建的常用措施之一,已被广泛应用于草地生态恢复工程中[15-16]。这是因为根据草地的自我恢复的能力,退化草地通常有可能在干扰停止较长时间后,恢复其生长和生态功能[17]。有研究表明,适度围封有利于退化草地植被和土壤恢复,长期围封会降低草地生产力,影响草地恢复效果[18]。但也有学者认为随着围封年限的增加,草地生产力依然增加[19]。关于放牧草地和围封草地间的碳通量的比较研究开始于1995年[20],从高山到热带草原围封持续时间在1-56 a的相关研究,表现出非常矛盾的结果,分别有54%、23%和23%的研究报告表明了围封后碳通量的增加、不变和减少[21]。这些结果表明围封对草地碳通量的影响仍然存在很大的不确定性,需要进行更多的研究[22]。由于学术界对围封措施的有效性存在争议,围封年限可能也是不同学者获得不同结论的重要原因[23]。因此,研究退化高寒草甸碳通量对不同围封年限的响应具有重要的科学意义。此外,需要注意的是,ER作为NEE的重要分量,本身也是一个非常复杂的生态学过程,因为它整合了来自不同呼吸源的通量[24]。ER依据释放来源的空间差别,可以区分为地上部分植物体呼吸(Vegetation canopy respiration,Rvc)和地下部分土壤呼吸(Soil respiration,Rs)两部分。而ER根据CO2释放来源生物属性,又常被拆分为植物体自养呼吸(Autotrophic respiration,Ra)和主要释放自微生物的异养呼吸(Heterotrophic respiration,Rh)。每个呼吸组分的生态学及生物学过程不同[25],对退化高寒草甸植被群落围封恢复年限的响应可能也存在差异,NEE组分的区分对于认识生态系统呼吸机制本身及其对退化草地围封年限的响应是很关键的,而退化高寒草地NEE组分对不同围封年限的响应研究相对薄弱。
基于此,本研究在青藏高原黄河源区退化高寒草甸进行不同年限(1 a、2 a、5 a、11 a)围封试验,监测了7-8月植物生长高峰期的NEE及组分。本研究试图回答两个主要问题:(1)退化高寒草甸NEE及组分对不同围封年限的响应是否相同?(2)土壤理化性质及生物量和NEE组分的关系怎样?
1 材料与方法
1.1 研究区域概述
研究区位于青藏高原黄河源区东部河南蒙古族自治县(34°52′51″ N,101°32′8″ E,平均海拔3590 m),属高原大陆性气候。该区全年四季特征不明显,每年5-10月温暖、多雨,11月至次年4月寒冷、干燥、多大风天气。年降雨量为597.1-615.5 mm,年均气温0.0℃。年日照时数2551.3-2577.2 h,日照率58.0%[26]。全县草地的92.68%为典型高寒草甸[27],研究区植被群落组成以高山嵩草(Kobresiapygmaea)为主,在青藏高原具有代表性,土壤类型为高山草甸土。研究地点位于黄河漫滩上,地形平缓,受到啮齿动物和过度放牧的影响,植被总盖度在80%以下,秃斑块占比20%以上,为中度退化草地。
1.2 试验设计和数据收集
1.2.1试验设计
分别选择2012年、2018年、2020年、2021年5月围建的地形平坦、植被群落特征基本一致和退化程度相似的斑块化退化高寒草甸作为试验样地,面积分别为300 m×250 m,围封后不进行放牧利用,截止2022年8月中旬围封年限分别为11 a、5 a、2 a和1 a。围栏外为高寒草甸正常放牧区(ck),属于牧民冬季牧场。不同围封年限高寒草甸地理位置、海拔高度和主要植物物种情况详见图1,表1。分别在各样地设置12个试验区组作为重复(区组间间隔超过30 m),每个区组内包含3个25 cm×25 cm样方。在每个区组随机选择1个样方用于原位观测(植被群落NEE和ER)和完成后期采样工作。此外,在该样方旁另设的2个25 cm×25 cm样方,以2种不同的处理方式随机分配,用以测定生态系统呼吸组分。区组内一个随机样方,采用根移走法,按0-5 cm、5-10 cm、10-15 cm、15-20 cm、20-25 cm、25-30 cm、30-35 cm、35-40 cm层次去除0-40 cm土壤中的根系(挖出每层土壤时用保鲜膜进行封存处理),沿坑边缘放置透明尼龙网(以确保地下水气流通,防止周围植被根系的进入),然后将无根土壤逐层回填,用于监测土壤异养呼吸速率(Rh)[28]。剩余的一个样方与以往测定土壤呼吸速率的方法保持一致,齐地面剪去地表植被,并在每次测定前一天剪去地面新萌发植被部分,保持样方地表裸露,测定土壤呼吸速率(Rs),处理后效果情况如图1所示。

表1 各区块特征Table 1 Characteristics of each block

图1 研究区及试验处理效果展示Fig.1 Study area and experimental treatment effect displayck,正常放牧;1 a,围封1 a;2 a,围封2 a;5 a,围封5 a;11 a,围封11 a
1.2.2植被群落NEE、ER、Rs及Rh测定
2022年7-8月植物生长高峰期,使用美国PP SYSTEM公司生产的多功能TARGAS-1便携式光合作用测量系统,利用CO2吸收激光电波束的红外光谱特性,以静态箱密闭式气路系统原理,明箱采用高透光率有机玻璃材料(汉莎科技集团有限公司制)同化箱(箱体大小为25 cm×25 cm×18 cm,内加设气体循环风扇并用12V蓄电池驱动),同化箱所设计的进气口和出气口通过塑料管与TARGAS-1连接,箱底座为不锈钢底槽,垂直砸入所选样点,测定时保持底座水平,以保证箱内气密性良好,监测生态系统的NEE。当NEE为负值时,表示碳固定,即为碳汇;NEE为正值时,表示碳释放,即为碳源。暗箱采用不透光黑布罩覆盖于明箱,测定ER及其组分(Rs、Rh)。测定是在每月13-17日早上9:00-12:00进行,每个样点测量300 s。为避免生态系统“CO2饥饿”现象,每次测定完后,将同化箱拿起通气10 s,确保箱内外气体浓度均匀一致。介于水汽对CO2吸收的影响,试验数据尽量选择晴朗天气测定。
1.2.3生态系统碳交换拆分及碳释放组成占比计算
总初级生产力(GPP)是ER与NEE的差值: GPP =ER-NEE
植被自养呼吸(Ra)是ER与Rh的差值:Ra=ER-Rh
植被冠层呼吸(Rvc)是ER与Rs的差值: Rvc =ER-Rs
根系呼吸(Rr)是Rs与Rh的差值:Rr=Rs-Rh
土壤呼吸占生态系统呼吸比例:Rs/ER
植被冠层呼吸占生态系统呼吸比例: Rvc/ER
植被自养呼吸占生态系统呼吸比例:Ra/ER
根系呼吸占生态系统呼吸比例:Rr/ER
土壤异养呼吸占生态系统呼吸比例:Rh/ER
1.2.4植被生物量测定及土壤理化性质分析
使用TDR 350在0-20 cm深度测量土壤含水量(Soil water content,SWC)和土壤温度(Soil temperature,Ts),与NEE测定同步进行。NEE、ER测定完成后,在8月中旬,将样方中的植被群落地上生物量全部齐地面剪下,并将这些样品放入纸袋中,待测定地上生物量(Above-ground biomass,AGB)。在剪完地上生物量的样方内用土钻(内径3 cm),采集表层土(0-20 cm)样品各9钻。其中3钻装入布袋用水漂洗,去除杂物,尽可能多地保持根量,以测定地下生物量(Below-ground biomass,BGB),所有植物样品在60℃的烘箱内烘干48 h至恒重并用分析天平称重。总生物量(Total biomass,TB)是AGB和BGB的和。剩余6钻土壤样品进行混合,使用2 mm筛网进行筛分,去除根系、石头等杂质,形成每个样方的复合样品。带回实验室,一部分新鲜土壤样本储存在4℃低温冰箱中,另一部分则自然风干后研磨过筛,供土壤元素分析用。采用K2Cr2O7-H2SO4法测定土壤有机碳(Soil organic carbon,SOC),全氮(Total nitrogen,TN)采用凯氏法消解法,全磷(Total phosphorus,TP)采用氢氧化钠熔融--钼锑抗比色法,用SEAL公司(德国)AA3连续流分析仪进行测定。
1.3 数据分析
采用单因素方差分析(One-way ANOVA)分析组间差异,检验不同围封年限土壤理化性质、碳通量及呼吸组分占比的差异,用Sigma plot 14.0完成绘制。利用Origin 2021对土壤温度和含水量与NEE及组分进行曲线拟合,土壤温度与NEE、GPP的关系采用二次曲线模型,土壤温度与ER以及其他呼吸组分的关系采用指数模型,土壤含水量与NEE及组分的关系均采用线性模型。利用R(v2.15.3)中的corrplot和hmisc软件包对有机碳、全氮、全磷、生物量、NEE及其组分进行相关性分析。
2 结果与分析
2.1 土壤理化性质及生物量变化
围栏封育会显著改变土壤温度(Ts)和含水量(SWC,P<0.01;图2)。随着围封年限的增加,Ts先降后增,而SWC先增后降。单因素方差分析结果表明,围封均显著降低了Ts,增加了SWC。围封5 a时的Ts(21.44℃)最低,而SWC(30.35%)最高。与ck相比,围封5 a后Ts显著降低了21.93%,SWC显著增加了100.66%。此外,围封会显著改变土壤有机碳含量(SOC)、全氮含量(TN)、全磷含量(TP)、地上生物量(AGB)和总生物量(TB,P<0.01;图2)。随着围封年限的增加,SOC、TN、TP和TB先增后减,在围封5 a时均达到最大值,AGB随着围封年限的增加而增加。其中,围封2 a、5 a和11 a的SOC显著高于ck,围封2 a和5 a的TN和TP显著高于ck、围封1 a和11 a,围封5 a和11 a的AGB和TB显著高于ck、围封1 a和2 a。

图2 土壤温度、含水量、土壤有机碳、全氮、全磷、地上生物量、地下生物量和总生物量Fig.2 Soil temperature, soil water content, soil organic carbon, total nitrogen, total phosphorus, above-ground biomass, below-ground biomass and total biomassck,正常放牧;1 a,围封1 a;2 a,围封2 a;5 a,围封5 a;11 a,围封11 a;条形表示平均值±SE,字母表示不同围封年限之间的显著差异
2.2 碳交换组分及占比变化
在7-8月生长高峰期,随着围封年限的增加,总初级生产力(GPP)增加,最大值出现在围封5 a时,围封11 a的样地GPP下降(图3)。围封5 a的GPP(1.33 g m-2h-1)显著高于ck(0.59 g m-2h-1)、1 a(0.91 g m-2h-1)、2 a(0.98 g m-2h-1)和11 a(0.68 g m-2h-1,P<0.01)。同时,围封1 a和2 a的GPP显著高于ck和围封11 a。随着围封年限的增加,生态系统呼吸(ER)最大值出现在围封5 a,不同围封年限间ER显著性差异结果和GPP表现一致。净生态系统碳交换(NEE)随着围封年限的增加,表现为先增后减,最小值出现在围封5 a。围封2 a(-0.34 g m-2h-1)和5 a(-0.42 g m-2h-1)的NEE显著低于其他(P<0.01)。

图3 总初级生产力、生态系统呼吸、净生态系统碳交换、植被冠层呼吸、土壤呼吸、植被自养呼吸、根系呼吸、异养呼吸Fig.3 Total primary productivity, ecosystem respiration, net ecosystem carbon exchange, vegetation canopy respiration, soil respiration, vegetation autotrophic respiration, root respiration and heterotrophic respirationck,正常放牧;1 a,围封1 a;2 a,围封2 a;5 a,围封5 a;11 a,围封11 a;条形表示平均值±SE,字母表示不同围封年限之间的显著差异
植被冠层呼吸(Rvc)、土壤呼吸(Rs)、植被自养呼吸(Ra)、根系呼吸(Rr)、异养呼吸(Rh)这些ER组分对围封的响应情况表现不同。其中,随着围封年限的增加,Rvc的变化规律和GPP一致,Rs、Ra、Rr的变化规律和ER一致。此外,除Rh外,围封5 a时的Rvc、Rs、Ra、Rr最大。围封5 a的Rvc和Ra显著大于其他,围封1 a和5 a的Rs显著大于ck、围封2 a和11 a,而围封1 a、2 a和5 a的Rr显著大于ck和围封11 a的。
不同围封年限生态系统呼吸组分占比分析结果表明,土壤呼吸Rs占生态系统呼吸ER比例均高于60%,Rvc在40%以下,但不同围封年限间Rs和Rvc所占ER的比例差异不显著(表2)。但是Ra、Rr和Rh占ER的比例在不同围封年限间差异显著(P<0.01)。其中,Ra占比随着围封年限的增加,先增后减,在围封5 a时最大(67.64%),围封11 a(42.67%)后显著下降,且为最小值。Rh的占比变化表现相反,围封11 a的Rh占比显著大于其他。此外,围封1 a、2 a和5 a的Rr显著大于ck和围封11 a的。
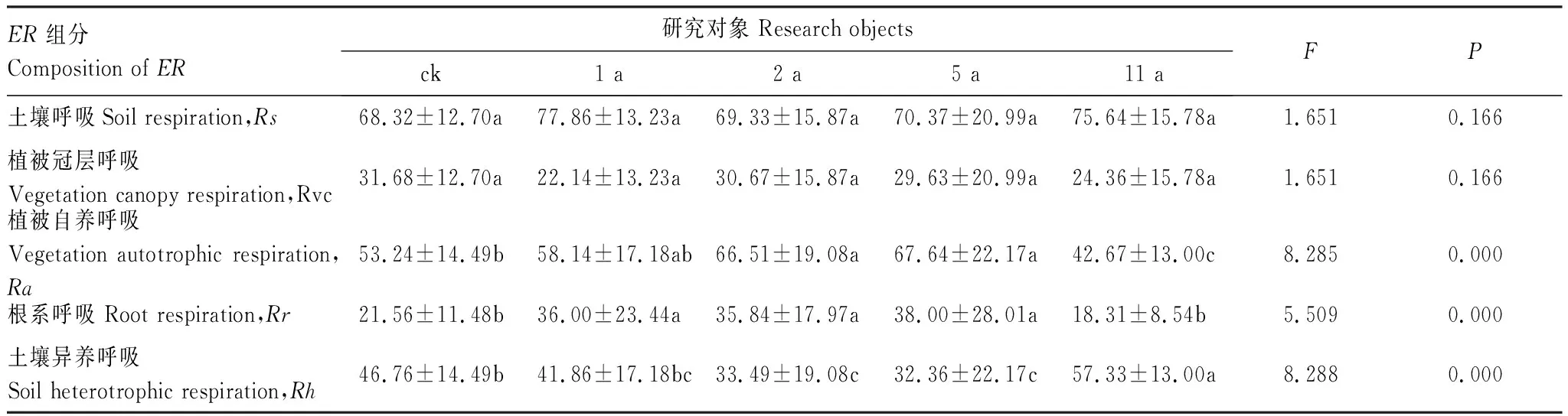
表2 不同围封年限生态系统呼吸组分占比Table 2 Proportion of components of ecosystem respiration in different enclosure years
2.3 相关性分析
土壤温度与NEE呈二次曲线的关系(P<0.01,图4),与ER以及除Rh以外的其他呼吸组分呈指数关系(P<0.01)。土壤含水量与NEE、GPP、ER、Rs、Ra、Rr呈线性关系(P<0.05),土壤含水量和NEE的相关系数最大,R2为0.4969。而土壤含水量与Rh和Rvc没有显著的关系。通过决定系数R2和P值可知,土壤含水量对NEE及组分的调控作用要大于土壤温度。此外,本研究发现TN、TP、AGB、BGB、TB和GPP显著正相关,它们与NEE显著负相关。TN、TP和ER、Ra,TN、TP、AGB和Rvc以及TN和Rr显著正相关(P<0.05,图5)。

图4 土壤温度(Ts)、含水量(SWC)和NEE组分的关系Fig.4 Relationship among soil temperature (Ts), water content (SWC) and NEE componentsGPP,总初级生产力;NEE,净生态系统碳交换;ER,生态系统呼吸;Rvc,植被冠层呼吸;Rs,土壤呼吸;Ra,植被自养呼吸;Rr,根系呼吸;Rh,土壤异养呼吸;ck,正常放牧;1 a,围封1 a;2 a,围封2 a;5 a,围封5 a;11 a,围封11 a;条形表示平均值±SE,字母表示不同围封年限之间的显著差异;Ts-Rh指数关系拟合失败,图中缺少P和R2

图5 有机碳(SOC)、全氮(TN)、全磷(TP)、地上生物量(AGB)、地下生物量(BGB)、总生物量(TB)和NEE及组分的关系Fig.5 Relationship among organic carbon (SOC), total nitrogen (TN), total phosphorus (TP), above-ground biomass (AGB), below-ground biomass (BGB), total biomass (TB), NEE and its componentsGPP,总初级生产力;NEE,净生态系统碳交换;ER,生态系统呼吸;Rvc,植被冠层呼吸;Rs,土壤呼吸;Ra,植被自养呼吸;Rr,根系呼吸;Rh,异养呼吸。* P<0.05, ** P<0.01, *** P< 0.001。ck,正常放牧;1 a,围封1 a;2 a,围封2 a;5 a,围封5 a;11 a,围封11 a
3 讨论
大量研究表明围栏封育是恢复退化草地碳汇功能的有效措施[29]。在本研究中,围封会增加斑块化退化高寒草甸植被群落GPP,围封1 a、2 a和5 a草地的GPP显著增加。这是因为围封后植被得到了休养生息,开始恢复[30],地上绿色植被是植物固定光能的唯一途径[31],地上生物量是生态系统的功能指标和植物光合作用获取能量能力的集中表现,与碳通量的大小显著相关[32]。植被恢复后,光合作用也会增强[33]。本研究发现地上生物量、地下生物量、总生物量和GPP显著正相关。GPP是生态系统中所有光合产物的总和,在光合作用过程中,植物将光能转化为化学能,用于合成有机物质,GPP是生物量增长的主要来源。此外,全氮、全磷和GPP显著正相关。这是因为土壤中的氮、磷等元素是植物生长所必需的营养元素,当土壤养分含量较高时,植物可以更好地吸收和利用这些养分,从而增强光合效率,促进植物生长和生产力的提高。反之,当土壤养分含量较低时,植物的生长和光合作用效率会受到限制,从而影响GPP。同时,光合作用增强时,有一部分光合产物通过根系分泌,输送到地下,提高土壤养分状况,增加土壤中的有机碳、全氮和全磷含量。长期围封至11 a时,只会显著降低土壤全氮和全磷含量,同时降低草地植被群落的GPP。这是因为随着围封时间的延长,植被群落内的生态过程逐渐达到饱和状态。植物的生物量增长趋势减缓甚至停滞,新的生长空间和资源变得有限。长期围封导致植物群落的结构发生变化,有研究表明禁牧对植物多样性产生负面影响[34],植被群落结构较自由放牧简单,长期围栏封育由于缺少家畜对牧草过腹还田作用使群落向均一化方向发展[35],还有毒杂草增加[36],对全氮和全磷需求较大的植物可能变为主导或者是土壤中的养分被植物长期吸收和利用,对草地生态系统产生的负面影响,从而使得土壤中营养元素的含量降低。同时,长期围封可能导致植物群落的年龄结构变老,老化的植物群落和增加的大量凋落物可能具有较低的生长速度和光合效率,从而降低草地的固碳能力。
不同围封年限间ER显著性差异结果和GPP情况一致,围封5 a时高寒草甸ER显著大于其他。在这里有两种组分来源变化可以解释围封5 a样地的ER最大。一方面,依据碳排放来源的空间差别,围封5 a时的地上部分植被冠层呼吸Rvc显著大于其他,围封1 a和5 a地下部分土壤呼吸Rs显著大于ck、围封2 a和11 a。另一方面,依据碳排放来源生物属性,围封5 a时的植物体自养呼吸Ra显著大于其他,围封1 a、5 a和11 a的异养呼吸Rh显著大于ck和围封2 a。因此,围封5 a时的ER最大。此外,Rvc和Rs占ER的比例在不同围封年限间差异均不显著。而Ra占ER的比例随着围封年限的增加,先增后减,Rh占ER的比例随着围封年限的增加,先减后增。这和围封后生物量的变化有关,在生长季尺度上,更高的生产力与呼吸通量的增加相吻合[24]。
净生态系统碳交换NEE随着围封年限的增加,先减后增,NEE在围封5 a时最小,围封2 a和5 a时的NEE显著小于其他。围封后植被开始恢复,生物量增加,NEE下降。NEE是植物光合GPP增益和生态系统呼吸ER损失的函数差值,它是由GPP和ER共同决定的。此外,本研究发现围封均会显著降低土壤温度,增加含水量,这是因为围封使得植被得以恢复,更高的植被覆盖和凋落物可以减少土壤的直射日照,从而降低土壤温度影响土壤微生物分解作用。同时,植被的恢复也有助于减少水分蒸发,增加土壤含水量有利于种子繁殖植物的快速生长发育。本研究也进一步证明了土壤含水量在生态系统碳收支的调控中起重要的作用,这是因为土壤含水量会显著影响土壤的物理性质和植被的生长以及土壤中的微生物活动[37],因此,提高退化高寒草甸植被可利用的土壤含水量也可以促进高寒草甸生态系统碳交换,这与胡毅等在新疆天山北坡草甸草原围封试验结果相似[33]。本研究的结果进一步强调了正确合理的草地利用方式不仅可以提高牧草产量,还有利于草地碳汇功能的发挥[38]。需要注意的是,年际变化对草地碳通量的预算估计很重要[39],未来的研究也将建立定位观测站点,进一步监测年际尺度上不同围封年限对黄河源区斑块化退化高寒草甸碳交换及其组分的影响,以提高预测和解释草地碳循环对未来全球变化的响应的能力。
4 结论
青藏高原黄河源区斑块化退化高寒草甸净生态系统碳交换及组分对不同围封年限的响应不同。基于本研究7-8月植物生长高峰期野外监测结果,发现围封5 a时的总初级生产力(1.33 g m-2h-1)和生态系统呼吸(0.91 g m-2h-1)以及固碳速率(0.42 g m-2h-1)最大,且与其他围封年限间差异显著。植被冠层呼吸、土壤呼吸、植被自养呼吸、根系呼吸、异养呼吸组分对围封的响应情况表现不同。随着围封年限的增加,植被自养呼吸、根系呼吸、异养呼吸所在生态系统呼吸的比例也会发生显著变化。土壤温度、含水量、全氮、全磷、生物量和净生态系统碳交换及组分存在显著相关关系。长期围栏并不会带来更多生态和经济效益,应采用短期围封来修复退化的高寒草甸。
