川西北高寒地区多年生禾本科人工草地土壤线虫群落动态
2024-01-17侯春雨马金豪王玉英吴鹏飞
侯春雨,魏 雪,周 磊,马金豪,任 晓,王玉英,吴鹏飞,*
1 西南民族大学青藏高原研究院, 成都 610225 2 云南大学云南省植物繁殖适应与进化生态学重点实验室, 昆明 650500 3 云南大学生态与环境学院, 云南省高校土壤生态与健康重点实验室, 昆明 650500
青藏高原是中国最大、世界平均海拔最高的高原,是长江、黄河重要的水源涵养地,素有“地球第三极”和“亚洲水塔”之称。高寒草地是青藏高原上最主要的生态系统,对维护我国生态安全发挥着极其重要的作用[1]。在全球气候变化和人类活动的综合影响下,青藏高原草地退化现象十分明显。据统计,青藏高原可利用草地面积为1.28×109hm2,退化草地面积约4.25×108hm2,占该地区草地面积的32.69%,严重退化草地面积约7.03×107hm2,占退化草地的16.54%[2]。高寒草地的退化降低草地生产力以及加剧草畜供需矛盾,从而制约草地生态系统和高原畜牧业的可持续发展[3]。
建植人工草地可以较快地恢复退化草地,调节植物群落结构、改善土壤理化性质和改变土壤碳汇功能,加快植被的正向演替,同时能够增加饲草产量及缓和草畜矛盾[4]。截止2013年,我国人工草地面积约2.09×107hm2,较1990年增加了2倍以上[5];另外,从2010到2020年,青藏高原人工草地面积由1.0×106hm2增加到3.0×106hm2,牧草良种繁育面积在7.0×104hm2的基础上增加了2.0×104hm2[6]。
垂穗披碱草(Elymusnutans)根系发达,具旱生结构的叶片,抗旱能力强,产量高,营养价值和适口性好[7]。老芒麦(Elymussibiricus)具有叶量丰富、高蛋白、草质柔软等特点,同时适应高寒、干旱、盐碱等环境,生态适应性较强[8]。两者被广泛应用于我国西北高寒地区人工草地建植以及退化草地补播改良。研究发现,垂穗披碱草人工草地物种多样性[4]、粗蛋白和粗脂肪含量[9]随种植年限的增加呈现下降趋势;生物量[3]和土壤碳储量[4]随种植年限的增加而增加。老芒麦人工草地的土壤有机质、全磷、速效磷以及土壤细菌群落丰富度[10]随种植年限增加呈下降趋势,而植物地下生物量[11]呈增加趋势。许多研究表明植物群落和土壤性质的变化是影响土壤动物群落组成和多样性的主要因素[12-14]。
土壤线虫作为土壤动物的优势类群,具有较高的生物多样性[15],参与有机物分解、养分循环、能量流动等,可改变土壤理化性质,调节土壤微生物群落等,是评价土壤健康状态的典型指示生物[16]。同时,植物群落通过影响土壤生态系统物质和能量的输入,可以驱动土壤线虫群落组成结构和功能的变化[17]。建植多年生禾本科人工草地生产力在高产3-5年后开始快速衰退[18]。本课题组前期对高寒地区不同人工草地间土壤动物群落多样性差异情况进行了研究[19-21],但目前并不清楚在多年生禾本科单播人工草地建植早期,土壤线虫群落随建植年限增加的变化动态及其主要影响因子。
本研究以青藏高原地区人工建植的垂穗披碱草和老芒麦单播人工草地为研究对象,以天然草地为对照,连续3年定位观测垂穗披碱草和老芒麦两种多年生单播人工草地土壤线虫群落及环境因子,旨在掌握天然草地转变为多年生人工草地后土壤线虫群落随建植年限的变化趋势及关键影响因子。研究结果可为评估人工草地健康状态,促进高寒草地的生态安全和持续利用提供科学依据。
1 材料与方法
1.1 试验地概况
试验地位于四川省阿坝藏羌自治州红原县(102°35′-102°58′E,32°5′-32°83′N),地处青藏高原东部边缘,平均海拔3600 m以上,地势为东南向西北倾斜,主要地貌类型为浅丘山地和山间低地。属于大陆性高原寒温带半湿润季风气候,气候寒冷,昼夜温差大,气温年较差小,冬季干燥夏季湿润,干雨季和冷暖季节分明,无绝对无霜期。日照时间长,太阳辐射强,年平均日照2417.9 h,太阳辐射年总量6.19×106kJ/m2。年均气温1.1℃,最低温度-10.3℃,最高温度24.6℃,极端高温25.6℃,极端低温-33.3℃。年降水量650-800 mm,80%集中在5-9月,年均蒸发量达1262.5 mm,水热同期[22]。该区域土壤类型主要为亚高山草甸土,占全县土地总面积的55.4%。植被覆盖度为70%-90%,植被类型主要为高寒草甸和灌丛。其中高寒草甸的植物种类主要包括禾本科的洽草(Koeleriacristata)、垂穗披碱草和草地早熟禾(Poapratensis)等,豆科的多枝黄芪(Astragaluspolycladus)和高山豆(Tibetiahimalaica)等,杂类草主要为鹅绒委陵菜(Potentillaanserina)、乳白香青(Anaphalislactea)和钝苞雪莲(Saussureanigrescens)等。
1.2 试验设计
2018年4月在西南民族大学青藏高原生态保护与畜牧业高科技研究示范基地设置人工牧草建植试验小区,选取青藏高原地区广泛种植的多年生牧草垂穗披碱草和老芒麦,连续观测3年。每种牧草设置5个5 m×5 m的种植小区作为5个重复,为了降低每个小区与相邻小区之间的边缘效应,每个小区水平间隔0.5 m,同时在试验区域内选取5个5 m×5 m的天然草地作为对照,共15个小区。各草地按照顺序排列,并使同种草地的小区不相邻。
在布置试验之前,天然草地无人工管理,存在放牧现象。人工草地由地势平坦且地上植被分布相对均匀的天然草地翻耕开垦,周围用铁丝围栏隔开。种植前进行机耕翻耙处理,后经人工整地,耙平、开沟条播,每年春季苗期施加225 kg/hm2复合肥(氮、磷、钾养分≥45%)做基肥,旱作无灌溉,生长期人工除杂,生长季完全禁牧,九月下旬刈割作为青贮饲草,留茬4 cm。
1.3 样品采集与处理
2018-2020年的9月进行植物群落高度(vegetation high, VH)、群落盖度(coverage, CO)调查及取样。利用直径为5 cm的土钻按对角线五点法采集每个小区0-10 cm的混合土样,装入做好标记的自封袋中,共采集土样45份(3种牧草×3个建植年限×5个重复)。将每个土壤样品一分为二,分别用于分离土壤线虫和测定土壤理化性质。此外,在每个小区内选取50 cm×50 cm的样方,用剖面方法采集地下根系。因高寒草地根系生物量主要集中分布于上层0-10 cm土壤中[23],所以根系采集深度为0-10 cm。将根系清洗干净后,置于65℃烘箱中烘干至恒重,研磨成粉状,用于测定其营养成分。
在实验室内,称取50 g混合土样,用湿漏斗法(Baermann 法)分离土壤线虫,连续分离48 h,用4%的甲醛溶液固定保存。依据《中国土壤动物检索图鉴》[24]和《长白山森林土壤线虫》[25]等参考书籍,在体式镜(Olympus SZX16)和显微镜(Olympus BX53)下将分离到的土壤线虫统计个体数量,并随机抽取100条线虫进行属的鉴定,不足100条则全部鉴定;土壤线虫的个体数量用条/100g干土表示。依据土壤线虫的食性以及形态学特征,将其分为食细菌线虫(Bacterivores,Ba)、食真菌线虫(Fungivores,Fu)、植物寄生性线虫(Plant parasites,Pl)和捕食/杂食性线虫(Predators-omnivores,Pr)4个营养类群[26]。
1.4 指标测定及方法
土壤理化性质测定:依据《土壤农业化学分析方法》[27]测定土壤含水量、土壤pH值、有机质、全氮、碱解氮、全磷、速效磷、全钾、有效钾。
根系营养成分测定:依据《饲料分析及饲料质量检测技术(第3版)》[28]测定牧草根系的全氮、全磷、全钾、粗蛋白、粗脂肪、粗纤维、粗灰分。
1.5 数据分析处理
优势度划分:个体数量大于总捕获量10%的为优势类群,1%-10%为常见类群,低于1%为稀有类群。

土壤线虫对外界扰动的响应采用自由生活线虫成熟度指数(maturity index, MI)和植物寄生线虫成熟度指数(plant parasite index, PPI)来评价[29]。主要计算公式如下:
MI=∑v(i)×f(i)
PPI=∑v(i)×f′(i)
式中,v(i)为第i类群的c-p值;f(i)为自由生活线虫个体数占总个体数的比重;f′(i)为植物寄生性线虫个体数占总个体数的比重。
外界扰动的程度和食物网的变化情况采用线虫功能团指数即富集指数(enrichment index, EI)、结构指数(structure index, SI)、基础指数(basal index, BI)和通路指数(channel index, CI)来表示[30]。指数计算公式如下:
EI=100×e/(b+e)
SI=100×s/(b+s)
BI=100×b/(b+e+s)
CI=100×ef/(eb+ef)
式中,b代表食物网中的基础成分,主要指食细菌线虫和食真菌线虫中c-p值为2的类群;e代表食物网中的富集成分,主要指食细菌线虫中c-p值为1和食真菌线虫中c-p值为2的类群;s代表食物网中的结构成分,包括食细菌线虫、食真菌线虫和杂食线虫中c-p值为3-5的类群以及捕食线虫中c-p值为2-5的类群。ef为食真菌线虫e组分值;eb为食细菌线虫e组分值。
利用主成分分析法(Principal component analysis,PCA)对不同种植年限垂穗披碱草、老芒麦种植地以及天然草地的土壤线虫群落进行排序。采用重复测量方差分析(repeated measures ANOVA)分别对草地类型和种植年限土壤线虫群落密度、多样性及生态指数的差异显著性进行检验。另用单因素方差分析(One-way ANOVA)分析相同牧草不同种植年限间的差异;对于不服从正态分布的数据进行Log(x+1)转换,如果仍不服从正态分布,则用非参数进行检验。为降低共线性环境因子的影响,对环境因子的膨胀因子VIF值进行检验,删除大于2的环境因子。采用冗余分析(redundancy analysis, RDA)解析不同牧草样地间土壤线虫群落与环境因子的关系,并用多元回归分析(multiple regression analysis)检验环境因子与土壤线虫群落指标的相关关系。
采用Microsoft Excel 2010、IBM SPSS 20.0、Origin 2018以及Canoco for Windows 5软件进行数据处理与分析。
2 结果
2.1 土壤线虫群落组成特征
共分离到土壤线虫38470条,隶属于2纲8目65科172属,平均密度2482条/100g干土。本实验土壤线虫群落组成中无优势属。营养类群的组成上,食细菌线虫平均密度最高,为1174条/100g干土,占线虫总数的47.31%;捕食/杂食线虫平均密度最低,为276条/100g干土,占线虫总数的11.12%。密度来看,食细菌线虫是研究样地土壤线虫的主要营养类群。
不同牧草样地土壤线虫群落组成存在一定的差异。在属水平上,垂穗披碱草样地的优势属为头叶属(Cephalobus)(占10.27%);老芒麦样地和天然草地无优势属。从各营养类群的组成情况看,食细菌线虫和捕食/杂食性线虫在天然草地、垂穗披碱草和老芒麦样地中分别占线虫总数的45.42%和12.64%,46.14%和12.02%,50.60%和8.52%。由此可知,食细菌线虫是多年生禾本科人工草地的主要营养类群。
2.2 土壤线虫群落组成结构差异
不同草地间土壤线虫群落主成分分析结果表明,2018-2020年间,垂穗披碱草和老芒麦样地的土壤线虫群落组成较相似,而两者与天然草地间的差异随建植年限增加而降低(图1)。在PC1和PC2轴上,影响群落组成结构的主要属随建植年限的增加而变化(图1)。

图1 相同建植年限不同草地间土壤线虫群落排序Fig.1 Principal component analysis on the soil nematode communities among different pastures in the same cultivation years NG: 天然草地 Nature Grassland; EN: 垂穗披碱草 Elymus nutans; ES: 老芒麦 Elymus sibiricus
相同草地不同建植年限间土壤线虫群落主成分分析结果表明,天然草地和垂穗披碱草样地的土壤线虫群落组成存在明显的年际间差异,而老芒麦样地2019年与2018和2020年间的差异较明显,后两者间差异不明显(图2)。不同草地中,在PC1和PC2轴上影响线虫群落组成结构的主要线虫属不同(图2)。

图2 相同草地不同建植年限间土壤线虫群落排序Fig.2 Principal component analysis on the soil nematode communities among different cultivation years within the same grasslands
2.3 土壤线虫群落密度及多样性动态
土壤线虫群落密度及多样性在不同草地之间无显著差异(图3)。2020年的线虫群落密度显著高于2018和2019年(P<0.001),2018年Pielou指数显著高于2020年(P<0.05),但类群数及Shannon多样性指数年际间差异不显著(图3)。进一步的单因素方差分析表明,2019年垂穗披碱草样地线虫密度显著低于天然草地,2020年垂穗披碱草Pielou指数显著低于老芒麦与天然草地(P<0.05)。天然草地、垂穗披碱草与老芒麦三种样地间的线虫类群数、Shannon多样性指数和Pielou指数这三个指数在同一建植年限的变化趋势相同。

图3 土壤线虫群落密度及多样性Fig.3 The abundances and diversity of the soil nematode communitiesGL:相同建植年限不同草地间差异显著性检验 Test of grassland effects on the abundance and diversity within the same cultivation year;CY:不同建植年限间差异显著性检验 Test of cultivation year effects on the abundance and diversity;GL×CY:草地和年间交互作用显著性检验 Test of interactional effects of grasslands and cultivation years on the abundance and diversity
2.4 土壤线虫群落营养类群结构动态
土壤线虫各个营养类群的属数在相同年限的三种草地间无显著差异(图4)。统计分析结果表明,在相同草地中线虫4个营养类群间的属数均存在显著差异(P<0.05),其中2018和2020年食细菌线虫属数总体上显著高于其它营养类群,但2019年三种草地中植物寄生性线虫的属数最高(P<0.05)。
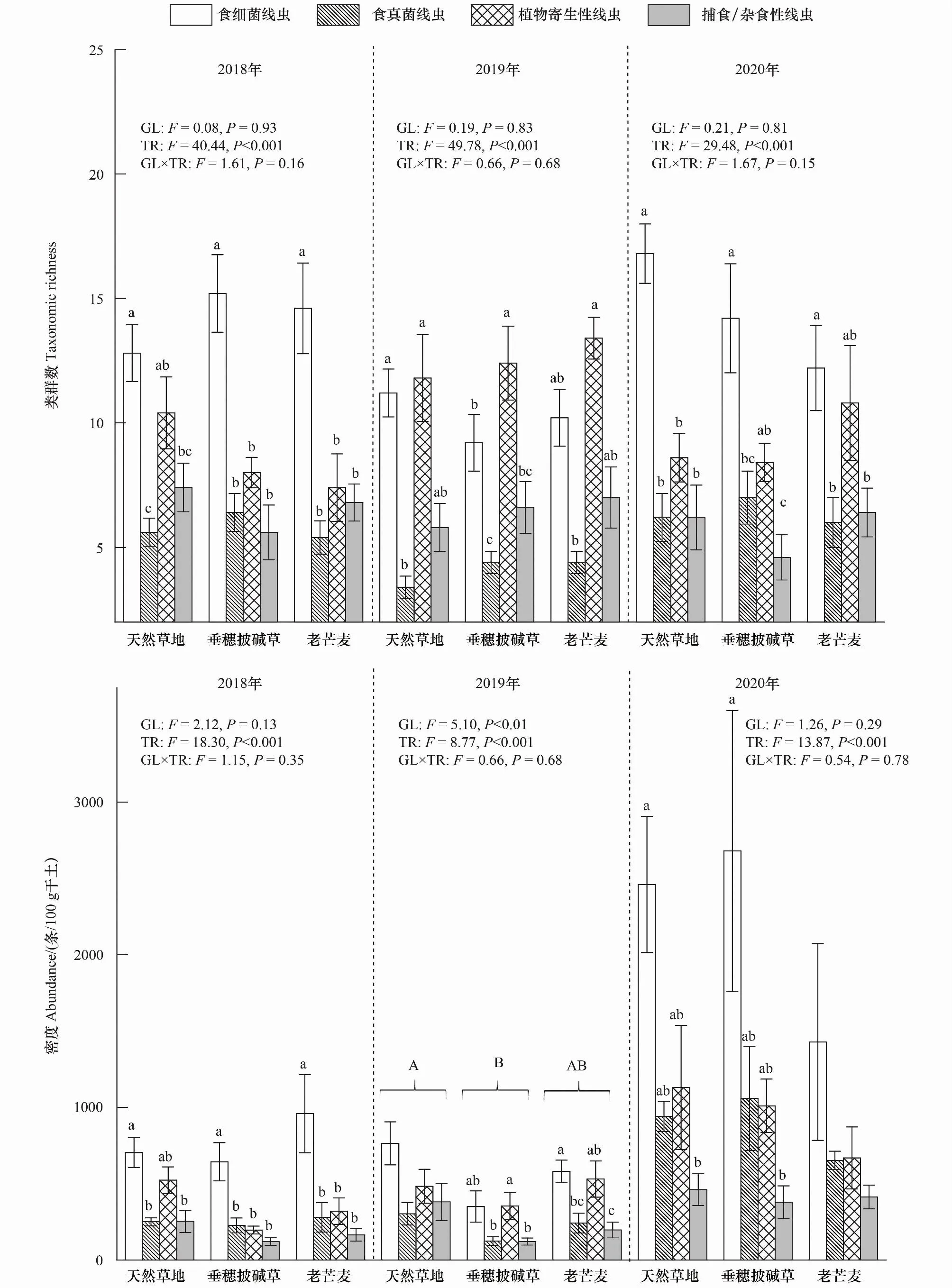

图4 土壤线虫各营养类群的类群数、密度与百分比Fig.4 Taxonomic richness, abundance and percentage of the four trophic groups of the soil nematodesTR:不同营养类群间差异显著性检验结果 Test of differences between different trophic groups;GL×TR:草地和营养类群交互作用显著性检验结果 Test of interactional effects between grasslands and trophic groups
仅2019年土壤线虫营养类群密度在不同草地间存在显著差异(P<0.05)。在相同草地中线虫各营养类群间的密度均存在显著差异(P<0.05)。其中,2018年,两种人工草地的食细菌线虫密度显著高于其他营养类群线虫(P<0.05),而天然草地中食细菌线虫和植物寄生性线虫密度无显著差异。2019年,垂穗披碱草人工草地植物寄生性线虫密度显著高于食真菌线虫和捕食/杂食性线虫(P<0.05);老芒麦人工草地食细菌线虫和植物寄生性线虫密度无显著差异,并显著高于食真菌和捕食/杂食性线虫密度(P<0.05);天然草地各营养类群间差异不显著(P>0.05)。2020年,天然草地和垂穗披碱草食细菌线虫密度显著高于捕食/杂食性线虫密度(P<0.05)(图4)。
相同建植年限土壤线虫营养类群百分比在三种草地间均无显著差异(图4),但相同草地的4个营养类群线虫个体百分比有显著差异,且变化趋势在不同草地间存在差异,其中2018年和2020年三种草地间食细菌线虫比例最高,2019年垂穗披碱草样地植物寄生性线虫比例最高(P<0.05)(图4)。
2.5 土壤线虫群落生态指数动态
土壤线虫群落6个生态指数在3种草地间均无显著差异。随建植年限增加,天然草地的富集指数呈持续增加趋势,垂穗披碱草人工草地则呈先下降后增加趋势(P<0.05)。两种人工草地的基础指数随建植年限增加均呈先增加后下降的趋势(P<0.05),而人工草地在不同年际间则无显著变化。此外,老芒麦人工草地的通路指数随建植年限增加而增加(P<0.05)(图5)。其他三个指数均无显著变化。

图5 土壤线虫的生态指数Fig.5 Ecological indices of soil nematode community
2.6 环境因子及其与土壤线虫的关系
观测的19个环境因子中(表1),植物高度、盖度分别以垂穗披碱草和天然草地最高(P<0.01)。天然草地的地下生物量显著高于垂穗披碱草和老芒麦样地(P<0.01),而植物根系全钾含量则呈相反趋势(P<0.05)。土壤pH、含水量、有机质、全氮以及速效钾均以天然草地最高(P<0.05)。

表1 不同草地的植物、根系及土壤化学性质特征(平均值±标准误)Table 1 The properties of plant, root and soil of different grasslands (Mean±SE)
通过共线性检验,从观测的19个环境因子中筛选出土壤全磷、全钾、有效磷、速效钾,植物高度、盖度,根系全氮、全钾、粗脂肪以及粗纤维共10个非共线性环境因子。该10个环境因子与土壤线虫群落的冗余分析结果(图6)可知,第一排序轴和第二排序轴对土壤线虫群落与环境因子关系的解释率分别为25.1%与6.4%。影响土壤线虫群落组成结构的主要环境因子是根系全钾(r=3.5,P<0.01)、根系粗脂肪(r=4.0,P<0.01)以及根系粗纤维(r=5.7,P<0.01)。由此可知,植物根系营养物质含量对线虫群落组成结构具有重要影响。

图6 土壤线虫群落与环境因子的冗余分析Fig.6 Redundancy analysis on the relationships between soil nematode community and environment factors
土壤线虫群落各指数与环境因子的多元回归分析结果表明(表2),群落密度、食细菌线虫和植物寄生线虫密度与根系全钾和粗脂肪含量呈显著正相关关系(P<0.05或0.01),与粗纤维含量呈显著负相关关系(P<0.01);植物寄生线虫密度还与植物高度呈显著负相关(P<0.05);食真菌线虫密度与根系全钾呈显著正相关(P<0.05),与土壤速效钾、根系粗纤维呈显著负相关(P<0.01);捕食/杂食线虫密度与土壤全磷、植物盖度呈显著正相关(P<0.05),与根系粗纤维呈显著负相关(P<0.05)。富集指数与植物高度、盖度呈显著正相关(P<0.01);结构指数(SI)与根系粗纤维呈显著负相关(P<0.05);基础指数(BI)与植物盖度、根系粗纤维分别呈显著负相关与正相关(P<0.05);通路指数(CI)与土壤有效磷呈显著负相关(P<0.05);植物寄生性线虫成熟度指数(PPI)与植物盖度呈显著正相关(P<0.05)。以上结果表明,土壤线虫群落密度主要受根系全钾、粗脂肪和粗纤维的影响;土壤线虫生态指数主要受植物盖度及根系粗纤维含量的影响。
3 讨论
3.1 建植多年生人工草地对土壤线虫群落组成结构及多样性的影响
天然草地转变为垂穗披碱草和老芒麦人工草地后,土壤线虫群落组成结构发生一定程度变化(图1),但2种人工草地及天然草地三者间的线虫群落密度和多样性指数总体上无显著差异(图3)。魏雪等[19]也发现高寒地区建植垂穗披碱草、老芒麦和早熟禾等人工草地间土壤线虫群落密度和多样性及其与天然草地间均无显著差异。其原因可能有以下三方面。首先,天然草地转变为垂穗披碱草和老芒麦人工草地后,植物群落组成和多样性发生显著变化,而土壤线虫组成受植物物种的影响[31]。其次,植物根系是连接地上植被和地下土壤食物网的关键纽带,在维持生态系统功能方面发挥着关键作用[32]。高寒地区多年生植物在秋季将养分转移到根系,以保证第二年返青期养分供给[33]。土壤线虫的营养物质直接或间接来源于植物根系[34],并在一定程度上依赖于根系生长[35]。虽然植物根系养分含量是影响线虫群落组成、密度及多样性的关键因子(图6,表2),但根系养分含量在3种草地间无显著差异(表1)。最后,部分土壤理化性质虽然在3种草地间存在显著差异(表1),但土壤理化性质与土壤线虫群落密度及多样性无显著相关关系(表2)。尽管土壤速效钾和全磷与食真菌线虫和捕食/杂食性线虫密度有显著相关,但这两个营养类群在线虫群落中所占比例相对较低,对线虫群落密度和多样性的影响较小[36]。因此,本研究的3种草地间土壤线虫群落密度及多样性差异不显著。此外,由环境因子与土壤线虫群落的关系可知,在高寒地区建植垂穗披碱草和老芒麦人工草地的早期阶段,植物群落根系营养物质含量对土壤线虫群落的影响强于土壤理化性质。
3.2 人工草地建植年限对土壤线虫群落组成结构及多样性的影响
土壤线虫群落密度和多样性的年际间变化动态受食物资源、气候、种间(种内)竞争等的影响。本研究发现,2020年土壤线虫密度和各营养类群密度显著高于2018年和2019年(图3、图4)。与吴雨薇等[37]在青藏高原三江源区不同恢复期人工草地土壤线虫多度随建植年限增加而增加的研究结果相似。其原因可能有以下三方面。首先,垂穗披碱草和老芒麦为多年生禾草,在建植早期植物生物量、群落盖度随建植年限的增加而增加[3, 11],提高了食物资源的数量和质量。改良土壤生物栖息环境,从而使土壤线虫群落特征发生显著变化。其次,根际微生物数量和多样性直接或间接影响食微线虫[38]。已有研究发现老芒麦人工草地土壤真菌数量随种植年限的增加呈上升趋势[39],垂穗披碱草混播人工草地也有类似发现[40]。再次,土壤线虫生活在土壤孔隙水膜中,适宜的土壤水分含量可促进线虫数量的增加,但过高或过低会限制线虫的运动、发育及繁殖[41]。2018年研究区7月降水量为212.7 mm,导致研究样地受到水淹干扰,土壤孔隙因积水缺氧而抑制线虫生长[42];2019年研究区7月降水量为94.3 mm,较低的土壤水分则限制线虫的移动与生长[43];2020年7月降水量为215.4 mm,虽然与2018年接近,但是由于实验样地的排水系统经过处理,没有再次被淹,线虫没有因强降水而受到干扰。
随建植年限的增加,土壤线虫的类群数和Shannon多样性指数无显著变化,但2020年Pielou指数显著低于2018年(图3)。这可能是由于食细菌线虫,尤其是小杆属、头叶属和棱咽属在2020年个体密度较高。其他研究也发现,部分土壤线虫对环境变化具有较强适应性,导致其优势度较高,从而降低线虫群落的均匀性和多样性[44]。
3.3 人工草地建植年限对土壤线虫群落营养类群结构影响
本研究发现,在第1年时,人工草地植物寄生性线虫的类群数、密度及个体百分比均显著低于食细菌线虫,而天然草地中两者无显著差异(图4)。这可能是因为植物寄生性线虫高度聚集在根的生长区与次生根区域,相较于其他营养类群线虫对根系具有较强依赖性[45];建植人工草地时对表层0-20 cm土壤进行翻耕等措施去除了原有植被及根系,改变了植物寄生线虫原有的生存环境。其他研究发现植物寄生性线虫密度和类群丰富度还受植物初级生产量、细菌和真菌或这些因素的综合影响[46],而高寒地区种植多年生禾本科牧草地上地下生物量在第1年时无法达到最大产量[11]。因此,多年生人工草地第1年时植物寄生性线虫密度及百分比显著低于天然草地。
第2年时,多年生人工草地的植物寄生性线虫类群数、密度和个体百分比均增加,并成为群落优势营养类群。其原因可能是垂穗披碱草和老芒麦根系生物量随建植年限的增加而上升,且根系代谢活动较强,促进根系的生长与分枝,增加根系生物量[47],为植物寄生性线虫提供了较多的食物资源与寄生空间。
第3年时,多年生人工草地土壤线虫各营养类群的密度与天然草地间无显著差异(图4),表明土壤线虫群落已恢复到与天然草地相似状况。这可能是因为随建植年限的增加,牧草残留物与根系分泌物在归还土壤的过程中增加了食物资源的多样性和异质性[48],促进食物资源累积以及微生物繁殖生长,为土壤线虫提供充足的食物资源和生态位空间。
3.4 多年生禾本科人工草地生态功能变化趋势
土壤线虫的生态指数能有效反映地下食物网结构的变化,揭示土壤生态学过程以及了解土壤有机质分解途径[30]。本研究发现,垂穗披碱草和老芒麦人工草地EI和BI指数高于天然草地(图5),表明建植多年生禾本科人工草地有利于土壤食物网资源的富集及增强食物网抵抗力。EI与植物高度呈显著正相关(表2),可能是因为垂穗披碱草和老芒麦是上繁疏丛型禾本科牧草,具有较高的光能利用率,提高植物生物量[49],增加土壤食物网资源的输入。而较高的食物资源输入为线虫在土壤团聚体周围和内部的水膜中觅食与繁殖提供了有力条件[50]。
研究表明对线虫栖息地的任何干扰都会影响其食物资源与环境稳定性[51],进而影响线虫群落组成与数量。垂穗披碱草和老芒麦人工草地的SI、MI和PPI指数低于天然草地(图5),表明建植多年生禾本科人工草地后土壤食物网稳定性、高营养级线虫抗干扰能力和繁殖能力均呈下降趋势,即土壤线虫群落中k-策略者比例较低。其原因可能是建植人工草地对土壤的翻耕和除杂等管理措施,干扰土壤环境的稳定性。c-p值较大的高营养级类群或k-策略者数量对食物网复杂性与稳定性发挥重要作用,但生命周期较长,在干扰后恢复速度较慢;而c-p值较小的r-策略者(快速生长的机会主义者),生命周期较短、繁殖率高、适应能力强,在扰动后能够快速恢复[52]。其他研究也发现,土壤扰动对处于高营养级的捕食/杂食性线虫影响较强,对食真菌线虫影响较小[30]。因此,建植多年生禾本科人工草地后土壤食物网结构稳定性受到一定程度的影响。此外,垂穗披碱草和老芒麦人工草地与天然草地间CI指数差异不显著且均低于50(图5),表明该区域的建植人工草地没有改变土壤食物网原有的能流渠道,仍以细菌分解为主。由于人工草地的地上生物量大部分被转移,易造成人工草地生态系统的物质输出大于输入;再加上在全球变化背景下青藏高原温度呈上升趋势,对土壤细菌群落的更替速率有促进作用[53],将加快土壤有机质分解和养分释放速率。最终会导致人工草地土壤养分亏缺,牧草生产力下降,这可能是高寒人工草地在建植4年后生产力下降的主要原因。
4 结论
高寒地区建植多年生禾本科牧草人工草地能够改变土壤线虫群落组成结构,但对群落密度及多样性影响不显著。随建植年限增加,土壤线虫密度呈上升趋势,均匀度指数呈下降趋势。线虫群落营养结构随建植年限变化明显,优势营养类群在第1和第3年是食细菌线虫,在第2年是植物寄生线虫和食细菌线虫。建植多年生禾本科牧草人工草地能够提高土壤食物网的资源富集能力,降低土壤食物网的结构稳定性。植物根系全钾、粗脂肪和粗纤维含量是影响土壤线虫群落的主要因子。建议减少对多年生禾本科人工草地施加钾肥,以提高土壤食物网的抵抗力。
