上海市14 种常见园林树种根系形态结构聚类研究∗
2024-01-16张德顺陈一家姚鳗卿李屹楠陈莹莹
张德顺 陈一家 张 振 姚鳗卿 郭 丽 李屹楠 陈莹莹
1 同济大学建筑与城市规划学院高密度城市人居环境生态与节能教育部重点实验室 上海 200092
2 四川农业大学风景园林学院 成都 611130
3 安徽新华学院 合肥 230088
根系作为植物六大器官之一, 不仅承担植物吸收水分和养分、 固定植株等多种生长发育功能[1], 还是植物发挥防风固沙、 涵养水源、 水土保持等生态功能的主要组织[2]。 根系形态结构(Root system architecture, RSA)[3-4]是植物根系正常进行生理活动和实现综合效益的关键生物学特性和生态习性, 是植物基因表达和环境影响共同作用的结果。 不同树种具有不同的根系类型, 植物学上一般将根系分为直根型和须根型两大基础类别。 在此基础上, 历经多年研究, 部分学者提出了更为细致的根系形态分类[5-7], 如台湾学者颜正平[5]基于根系的分支形式, 将根系形态结构分为PH 型、 V 型、 VH 型、 R 型和M 型5 种基本型。 其中, PH 型根系指大部分根系水平横向生长, 即大部分根为水平根; V 型根系指垂下根较为发达, 侧根不伸展, 水平根与斜出根少; VH型根系水平根、 斜出根与垂下根均生长良好; R型根系多斜出根, 根幅较宽; M 型根深较浅, 多须根, 分布于土壤上层。 苅住曻[1]对日本457 种木本植物进行了根系结构研究, 将根系结构分为4 种类型, 即浅根集中型、 中心型、 深根主根型和深根鱼尾型。 这种分类方法较为简洁, 且每一类根系的特征都较为明显。
近年来, 根系研究更为关注其形态指标与锚固特性的关联性[8-9]。 常见根系形态的定量化指标包括根系长度、 体积、 面积、 根系分支结构、根尖数、 平均根径、 比根长、 比根面积和根密度等[10-13]。 然而, 由于试验条件和试验目的的差异, 各项研究对根系形态的描述重点各不相同。 根系形态参数选择侧重点的差异限制了不同研究之间的可参考性。 传统的根系分类要适应定量化指标体系的发展和现代研究需求而更新, 从而提高不同根系研究间的参考性, 增强城市绿地系统中植物应用与选择的科学性及其对气候变化的适应性。 因此, 参考前人研究成果, 试图整理总结出一套较为全面综合、 易于获取的根系形态指标量化体系, 便于对根系形态结构系统性的描述和概括, 进而为树木根系形态聚类分析提供依据。
1 研究区概况
试验场地位于上海市宝山区的森林植被种质资源基地(31°23′33″N, 121°19′55″E)。 试验场地中的土壤质地为黄棕壤, 黏度适中, 30 cm 深度下土壤紧实度为2 005~2 011 kPa, 土壤容重为1.30~1.37 g•cm-3, 土壤性质较为稳定。
2 研究方法
2.1 树种选择
本研究共筛选了14 种上海园林树种, 包括三角槭(Acer buergerianum)、 天竺桂(Cinnamomum japonicum)、 野漆 (Toxicodendron succedaneum)、细叶青冈(Quercus shennongii)、 女贞(Ligustrum lucidum)、 枫香树(Liquidambar formosana)、 北美枫香(L.styraciflua)、 水紫树(Nyssa aquatica)、豆梨 (Pyrus calleryana)、 德州栎 (Quercus nuttallii)、 南京椴 (Tilia miqueliana)、 乌桕(Triadica sebifera)、 榉树(Zelkova serrata) 和培忠杉(Taxodiomera peizhongii)。 每树种选择3 株共42 株苗木作为试验对象, 用于重复试验以提高数据代表性, 所有试验对象生长环境一致、 生长情况良好, 且胸径控制在5.5 cm ± 0.7 cm, 高度控制在3.5 m ± 0.5 m, 以减少土壤环境差异、 树种规格等因素对试验结果的影响。
2.2 根系结构获取
试验在2021 年5 月下旬进行, 试验开始前一周均为晴天。 “完全挖掘法” 具有操作简单、 结果准确和试验限制因素少等优势[14], 本研究选用此法以获取较为精确的根系结构数据。 通过预挖掘确定直径1.5 m, 深0.8 m 的圆形树坑以尽量减少对根系的机械损伤, 并收集被试树种的完整根系结构。 挖掘出树根后, 去除根系间的土壤,拍摄以获取完整准确的树根结构图像, 图像经Photoshop、 CAD 等软件处理后用于量化分析根系结构的相关参数(图1)。 根系结构获取中, 榉树的一个试验个体的根系有明显人为锯断的痕迹,故将其排除。
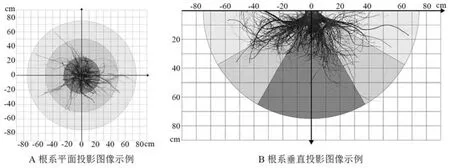
图1 根系图像获取
2.3 数据分析
基于CAD 和Photoshop 处理根系图像, 统计得到各指标数据后, 运用SPSS 26.0 结合其根系形态结构指标体系数据进行聚类分析。
3 结果与分析
3.1 根系形态结构指标体系的建立
根系结构的量化分析是了解其在物种间差异的重要手段。 根系的各种形态变量越丰富, 描述的根系形态结构特征也就越全面。 广义上讲, 根系形态相关变量主要包括根系分区(水平根、 斜出根和垂下根)、 垂直分布(浅根型、 中间型和深根型)、 水平分布 (分散性、 中间型和集中型)、 细根数量(密生型、 中间型和疏生型)、 细根大小(纤细型、 中间型和肥厚型)、 根系发育(优秀、 良好、 中、 不良和极差) 等。
为了方便测量数学建模, 目前已有的研究中也采用了根横截面尺寸、 根的分布、 隆起的根/土壤体积(Uplifted root/soil volume)、 树木断裂模数(Greenwood modulus of rupture) 和土壤/树木密度等更为直观的参数[15]。 细化到根系某一部分的结构时, 部分学者则使用了一些更为具体的变量,包括主根基底直径、 主根深或浅根数量等[16]。 根角度也是常用的形态指标, 依据根与地面产生的夹角不同, 可分为水平根、 斜出根与垂下根[17-18]。
本研究结合文献综述,整理出由经典指标构成描述根系形态的指标体系,包括根系总生物量、根系结构分布、根系直径、根系密度4 类共16 项指标(表1)。
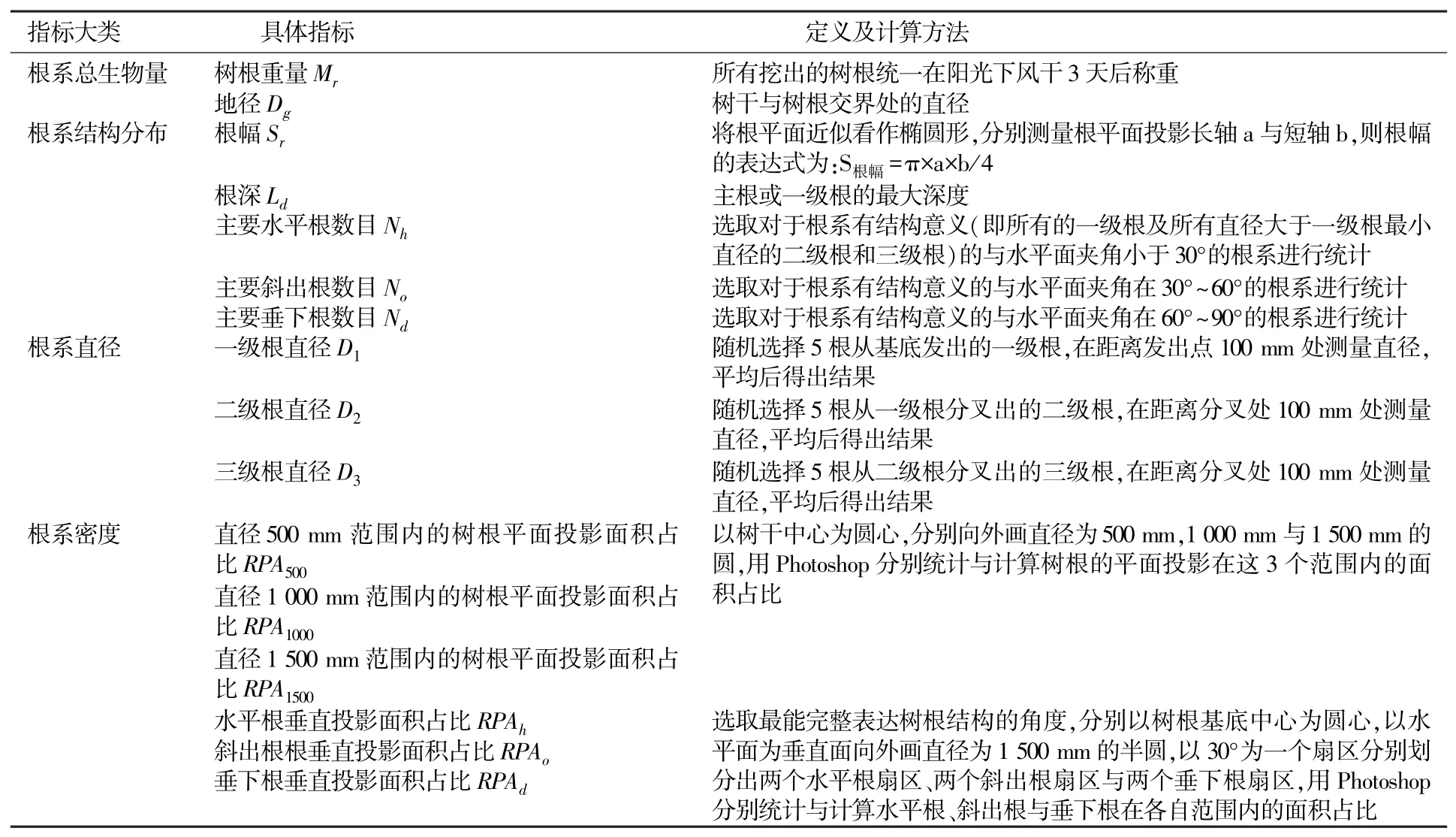
表1 根系形态结构指标体系
3.2 14 种树种根系形态结构特征
对比分析不同树种的根系图像, 可以得到不同树种的根系形态特征(表2)。 从树根挖掘的整体情况上看, 不同树种的根系差异比较明显, 同一种树的树根虽然也会显示部分差异, 但其生长基本都遵循了类似规律。 部分同科的植物, 如北美枫香与枫香树、 德州栎与细叶青冈, 其根系也呈现出了一定的相似性。
3.3 基于新的根系形态结构指标体系的聚类分析
根据表1 根系形态指标体系统计整理14 种树种根系形态结构参数(表3), 参考前人的根系分类方法与实际根系形态结构, 最终将树根分为4大类(图2)。
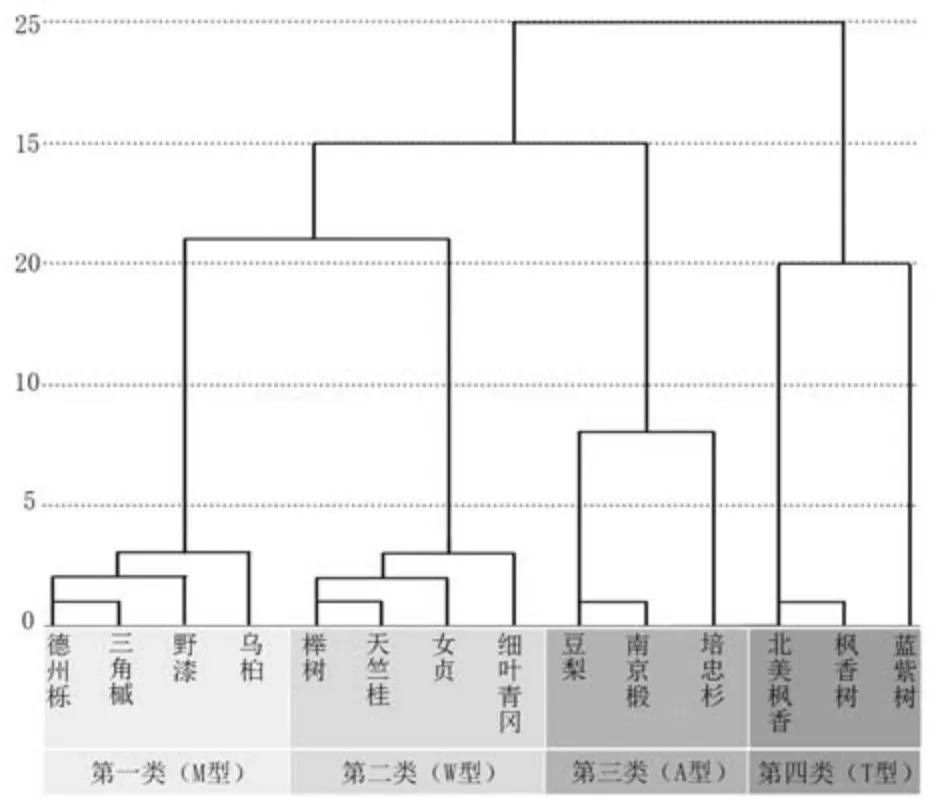
图2 树种根系聚类分析
第一类(图3A): 德州栎、 三角槭、 野漆、乌桕。 主根明显, 水平根、 斜出根和垂下根均分布密集, 根幅大, 根深较深, 根整体的体积比较大, 对应于苅住曻[1]描述的深根主根型根系特征。因为从立面来看根系的整体轮廓与英文字母M 较为相似, 故将此类根系称为M 型根系。
第二类(图3B): 女贞、 榉树、 天竺桂、 细叶青冈。 没有明显主根, 树根分叉较多, 绝大部分的根系结构都集中在距离地表40 cm 以内的深度, 但根幅较大, 水平根较多, 符合苅住曻[1]描述的浅根集中型特征。 因为从立面来看根系轮廓与英文字母W 较为相似, 故将此类根系称为W型根系。
第三类(图3C): 培忠杉、 豆梨、 南京椴。没有明显主根, 但一级根较为粗壮, 须根较少,多斜出根, 水平根与垂下根分布较为均匀, 根幅偏大, 符合苅住曻[1]描述的中心型根系特征。 因为从立面来看根系的分叉形式与英文字母A 较为相似, 故将此类根系称为A 型根系。
第四类(图3D): 北美枫香、 枫香树、 水紫树。 主根粗壮, 多垂下根, 少水平根与斜出根,有须根, 根幅较小但根深较深, 符合苅住曻[1]描述的深根鱼尾型特征。 因为从立面来看根系的分叉形式与英文字母T 较为相似, 故将此类根系称为T 型根系。
4 讨论
4.1 根系形态结构聚类结果的匹配性
本研究通过对根系样本的聚类分析, 同时参考前人的研究成果, 最终将树种根系形态结构分为M 型根、 W 型根、 A 型根和T 型根4 种。 关于根系形态结构类型的分类体系, 最被国内学界认可与接受的是颜正平的分类方法, 他将根系结构分为PH 型、 V 型、 VH 型、 R 型和M 型5 种[19]。试验中获得的根系样本基本能被归纳为前4 种类型, 而只分布于土壤表层、 多须根的M 型根在本次试验中并没有遇到。 从另一个角度来看, 颜正平[5]的M 型根系与PH 型根系在描述与示意上仍较模糊, 考虑到两者均多水平根, 且分布于浅层土壤中, 其差别更多的是根系的生物量, 这就给鉴别与分类带来了一定的困难。 比如颜正平[5]将枫香树归为PH 型根系, 但在本研究聚类中枫香树与T 型根系更为接近。 相比较而言, 苅住曻[1]的分类方法更为简洁明了, 即将根系结构分为浅根集中型、 中心型、 深根主根型和深根鱼尾型4种, 且每种根系间均有较为明显的区别。 本研究涉及的三角槭、 女贞、 枫香树、 乌桕、 榉树5 种树种均与苅住曻[1]的研究分类结果一致。 整体看来, 本研究的聚类结果与苅住曻[1]的分类匹配度更高。
从分类体系的角度来看, 颜正平[5]的指标包括细根不同深度分布类型、 细根数量、 主根深度及其分布、 主根数量、 根深、 根系分支方式等8项指标。 而苅住曻[1]进行了更大量的样本调查和统计分析, 分类指标也更为全面, 包括垂直分布、水平分布、 根系分支方式、 细根生物量及根径、根毛类型。 比较之下, 苅住曻对根系形态的考量更为全面, 水平投影上根系分布特征、 根系分支方式等都是颜正平的评价体系没有涵盖到的。 对比本研究的指标体系, 也与苅住曻[1]更加一致,比如根系结构采纳了苅住曻的水平根、 斜出根、垂下根体系, 根系分级描述直径, 以及综合水平面与垂直面考虑根系密度。 尽管苅住曻[1]对木本植物根系展开了广泛研究, 但其分类依据复杂且缺乏定量化统计的论证, 本研究恰好从数理统计的角度为苅住曻的分类提供了论证。 当然, 要获得更为准确可靠的结论还需要对更多树种样本进一步研究与分析。
4.2 植株聚类与树种聚类的差异
考虑到部分树种存在明显的个体差异, 本研究也补充了以试验个体为单位的聚类分析, 以探究个体差异对根系聚类结果的影响。 对比个体和树种聚类结果发现, 根系结构性状较为稳定的树种为枫香树、 北美枫香、 水紫树、 培忠杉和榉树,而其他9 个树种存在个体差异而导致两种聚类结果不同的情况。
4.2.1 水紫树的聚类差异
对树种聚类时, 水紫树与枫香树、 北美枫香同时被归结为T 型根, 而对试验个体聚类时, 水紫树和后二者差距甚远。 仔细对比三者的根系结构发现, 这种差异主要是由于聚类标准是否细致决定的。 水紫树拥有数量少、 粗且长的平行根,且根深更深, 根幅更广, 确实与北美枫香、 枫香树存在一定的差别, 但这3 种园林树种与其他树种对比, 则均拥有十分发达的垂下根结构。 因此,参考前人的研究成果, 倾向于将水紫树、 北美枫香和枫香树的根系结构统一归纳为T 型根, 以避免出现分类过繁的情况。
4.2.2 植株差异的类型
9 种差异显著的树种包括: 细叶青冈、 女贞、天竺桂、 南京椴、 乌桕、 豆梨、 德州栎、 三角槭和野漆。 进一步观察这9 种树种的根系结构样本,发现生长差异是造成植株个体差异的最主要原因。虽然试验前尽可能排除试验个体的规格、 生长环境等因素导致的误差, 但依然会出现因为隐性的生长环境差异而出现生长发育上的差异性[20-22],如个体差异最大的细叶青冈, 造成聚类差异的指标是根系生物量, 但从根深、 根幅、 根系直径等其他指标和根系结构投影图均可判断三个样本均符合W 型根系的结构特征, 其他树种亦然。 这说明虽然生长差异导致的个体聚类差异客观存在,但通过进一步比较分析可以确定树种根系结构间不存在本质差异, 可以归结为同一结构类型。 第二个可能的原因是实生苗的遗传特性差异。 相关研究已证明实生苗根系结构受到基因型的影响而表现出明显的差异[18], 而同一无性系的植株则具有更为稳定相似的根系形态, 如野漆3 号根系的须根比1 号和2 号植株个体更多、 2 号根垂下根未发育的情况。 由于本研究中同一树种的样本量较为有限, 且未涉及基因层面的探究, 因此尚难以判断这种个体性状差异的幅度和频率。 未来研究应对这两种树种根系结构进一步广泛取样, 以获得更加全面的数据和结论。
从科属的角度看, 同科植物的亲缘关系更近,植物根系结构应更为相似。 比如, 枫香树和北美枫香同属于金缕梅科枫香树属的植物, 其根系结构都为T 型根, 垂下根多且粗壮, 而且根系的结构形状也都十分稳定。 同时也存在这种现象, 即德州栎和细叶青冈虽然同科, 但根系结构却归属不同类别。 因此, 树种的科属关系只能作为一种补充手段, 在试验前期的树种选择与后期的结构分类中起到辅助作用。
5 结论
园林树木根系形态对其个体的生长发育、 群体的生态功能都有重要影响。 本研究梳理出一套较为全面的根系形态结构指标体系, 包括根系总生物量、 根系结构分布、 根系密度、 根系直径等4 类共16 项指标。 基于该体系分析14 种园林树种的根系形态量化数据并进行聚类分析, 得到M型、 W 型、 T 型和A 型四种根系结构类型, 该聚类结果与苅住曻的分类结论较为匹配, 从数理统计角度为根系分类提供了佐证。 园林树种根系形态指标体系的归纳有助于推进根系研究体系的构建, 同时也需要更多研究补充和加深根系分类领域的认识。 本研究结果亦可对进一步园林树种根系的功能性状研究和应对气候变化的人居生态建设提供理论参考。
