采食高淀粉高油日粮奶牛的泌乳性能、乳成分变化及乳腺差异表达基因筛选
2024-01-15韩印如刘肃然毕研亮刘德武郭勇庆
吴 柯,韩印如,李 斌,刘肃然,贾 坤,毕研亮,刘德武,郭勇庆
(1 华南农业大学 动物科学学院,广东 广州 510642;2 华南农业大学 兽医学院,广东 广州 510642;3 中国农业科学院 饲料研究所,北京 100081)
近年来,我国奶业发展迅速,奶牛养殖业和乳品加工业逐渐成为国民经济中重要支柱产业之一[1]。乳脂作为牛奶营养的重要物质之一,是衡量牛奶品质优劣的重要指标,其含量直接影响原奶品质和售价。然而,现行生鲜乳国家标准中要求的乳脂率仅为3.1%,我国原奶乳脂率的整体水平为3.69%,远低于欧盟(4.04%)、澳大利亚(4.11%)和新西兰(4.78%) 等乳业发达国家生鲜乳中的乳脂率[2]。乳脂率受遗传、饲粮、泌乳阶段等多种因素影响,其中受饲粮组成影响最大。乳脂降低综合征(Milk fat depression,MFD)是指日粮因素造成奶牛乳脂率或乳脂产量明显降低,而对产奶量和乳蛋白产量影响较小的一种现象[3]。我国优质粗饲料资源短缺,为了使奶牛充分发挥产奶性能,生产中不得不饲喂高淀粉高油饲粮来满足奶牛能量需要,造成乳脂合成抑制多发[4]。过去的研究通常采用单独的高精料或高油饲粮来诱导奶牛产生MFD,而给奶牛同时饲喂高精料高油饲粮的研究相对较少。
日粮组成会影响反刍动物乳腺中乳脂肪和乳蛋白的合成。研究表明,在日粮诱导奶牛发生MFD 过程中参与脂质合成的关键酶下调,启动子中有一个受固醇反应元件SREBP1调节[5],表明SREBP1在调节牛乳腺上皮细胞脂肪酸合成的信号通路中具有重要作用。RNA 测序技术(RNAseq) 已成为基因表达和转录组分析的重要手段,Cui 等[6]以荷斯坦奶牛乳蛋白和乳脂肪含量作为差异性状,对奶牛的乳腺上皮细胞进行转录组测序,推测出了调控乳中蛋白和脂肪含量的关键基因。因此,利用RNA-seq 技术阐明牛乳腺转录组对于确定影响奶牛乳成分性状的候选基因至关重要[7]。
本研究旨在探究同时饲喂高淀粉高油日粮对奶牛生产性能和乳成分的影响,并结合乳腺组织的转录组表达情况,筛选出参与脂质调控的候选基因,为进一步阐述MFD 的分子机制和科学配制日粮提供数据支持。
1 材料与方法
1.1 试验动物和饲养管理
选取来自广东某奶牛场的8 头头胎荷斯坦奶牛为试验动物,其泌乳日龄(215±34) d,体质量(575±23) kg。全混合日粮(Total mixed ration,TMR)饲喂,每天饲喂时间为07:00 和17:00,挤奶时间为06:30 和16:30,自由采食和饮水,保证剩料量(w)5%~10%。试验期间,每头奶牛单独拴系饲养于栏舍内,地面铺有橡胶垫和细沙,每日清粪2 次。
1.2 试验设计
试验开始前预饲10 d,奶牛均饲喂低淀粉低油饲粮。试验期将8 头奶牛随机分为对照组和处理组(每组4 头),采用2×2 反转试验设计,进行两期试验,每期23 d(诱导期16 d,恢复期7 d),两期间对照组和处理组对调。诱导期时,对照组奶牛饲喂低淀粉低油日粮,精粗比(m:m) 50:50,处理组奶牛在对照组日粮基础上添加了266 g/kg(干物质基础)细粉碎玉米(过1.2~1.5 mm 筛) 和46 g/kg(干物质基础)的大豆油,精粗比(m:m) 62:38;恢复期,两组奶牛均饲喂低淀粉低油饲粮。诱导组奶牛在每期试验的前4 d 进行饲粮过渡。饲粮组成和营养成分见表1。
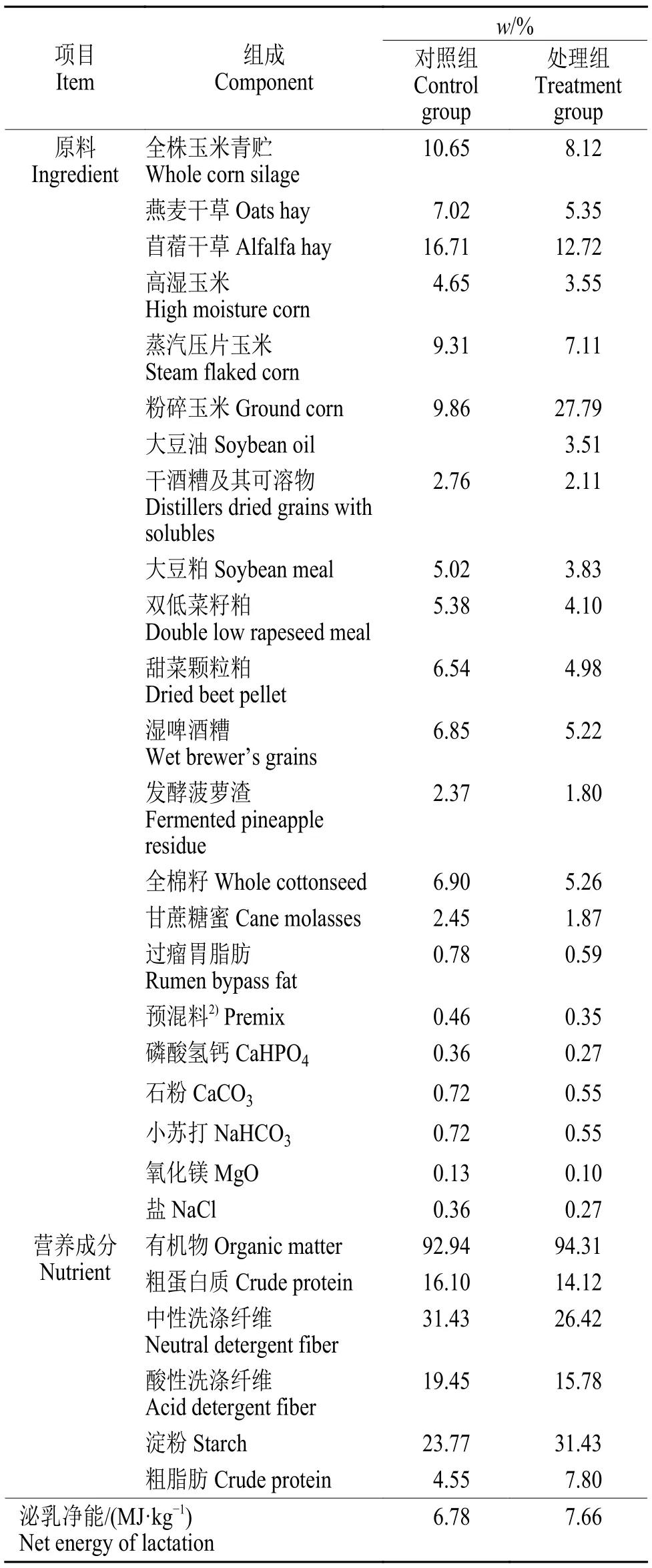
表1 试验饲粮组成和营养水平(干物质基础)1)Table 1 Composition and nutrient levels of experimental diets (DM basis)
1.3 样品采集
1.3.1 饲料样品采集和测定 饲养过程中每天记录每头牛的TMR 饲喂量和剩料量,测定第1、4、7、10、13、16、19 和22 天的干物质采食量(Dry matterintake,DMI)。于每期的第13~15 天采集饲料原料和TMR,65 ℃烘干后粉碎,过18 目筛后,参照《饲料分析及饲料质量检测技术》[8]中的方法测定干物质(Dry matter,DM)、有机物(Organic matter,OM)、粗蛋白质(Crude protein,CP)、粗脂肪(Ether extract,EE)、中性洗涤纤维(Neutral detergent fiber,NDF)和酸性洗涤纤维(Acid detergent fiber,ADF)含量,采用比色法测定淀粉含量[9]。
1.3.2 奶样采集和测定 测定每期第1、4、7、10、13、16、19 和22 天的产奶量,并采集奶样。将每天采集的早晚奶样按照1:1 体积比均匀混合后4 ℃保存,利用多功能乳成分分析仪(LactoStar 3560;FUNKE GERBER,Germany)测定乳脂、乳蛋白、乳糖和非脂固形物含量。
1.3.3 乳腺组织采集与测定 于饲养期第17 天由经过培训的专业兽医人员对奶牛进行乳腺组织的活体采样,其中每期每个处理组随机采集乳腺样本2 个(共8 个)。采集乳腺组织时,选择奶牛乳房后区上四分之一中点处为采样部位。皮下注射体积分数为1%的盐酸普鲁卡因(每头注射20~30 mL)进行麻醉。麻醉起效后,使用12 号活检针移除适量乳腺组织,用生理盐水洗去血液,放入冻存管,存于液氮中待测。观察伤口不再出血时(活检后约5 min)将其缝合。转录组测序由北京诺禾致源科技股份有限公司完成,测序文库使用NEBNext®Ultra™Directional RNA Library Prep Kit for Illumina®(NEB,USA)生成,高通量测序平台为Illumina HiSeq 4 000(Illumina,USA)。
1.4 数据统计分析
采用Excel(2016)进行所有数据的基本处理。奶牛的DMI、产奶量和乳成分结果折线图采用GraphPad Prism 7 绘制;各指标采用SAS version 9.4 的MIXED 程序进行统计分析,使用t检验比较差异显著性,根据以下模型进行分析:Yijkl=μ+Si+Tj+Nk(Si) +Pl+εijkl,其中Yijkl为试验牛在不同处理下的因变量值,μ为总体均值,Si为次序i的随机效应(i=1、2),Tj为日粮j处理效应(j=1、2),Nk为嵌套次序的试验牛k的随机效应(k=1~8),Pl为采样时期效应(l=1、2),εijkl为随机误差。使用Kenward-Rogers语句校正自由度。剔除学生化残差(Studentized residuals) 超过±3 的数据点。采用的软件方法为GOseq 的GO 分析,使用KOBAS(2.0)和KEGG 数据库对差异表达基因进行通路显著性富集分析,确定差异基因的主要生化代谢途径和信号转导途径。基于皮尔逊相关系数对奶牛的平均泌乳性能与部分乳腺差异表达基因进行相关分析。
2 结果与分析
2.1 高淀粉高油日粮对奶牛DMI、产奶量及乳成分变化的影响
由于饲粮对奶牛的影响是渐进的过程,因此本试验进行了分阶段采样,以观察饲养期间奶牛相应指标的变化情况(图1)。由图1a 可以看出,处理组DMI 在诱导期呈波动性变化,整体低于对照组,其中第13、16 天显著低于对照组(P<0.05),恢复期逐渐回升。图1b 表明,试验初期处理组产奶量略高于对照组,第4 天后处理组逐渐下降,其中第13、16 天显著低于对照组(P<0.05),恢复期有所回升。乳成分方面,与对照组相比,饲喂处理组饲粮7 d 后乳脂率和乳脂产量整体呈降低趋势,且第10、13、16 和19 天显著降低(P<0.05)(图1c、1d);乳蛋白、乳糖和非脂固形物的含量先升高后降低(图1e、1g、1i),而产量先降低后回升(图1f、1h、1j),其中第13、16、19 天差异显著(P<0.05)。
2.2 高淀粉高油日粮对奶牛乳腺组织转录组的影响
2.2.1 RNA 抽提结果与质量检测 共提取了泌乳奶牛8 个乳腺组织RNA 以供建库使用。乳腺处理组和对照组每组各取4 个样本,通过Nanodrop 和Agilent 2 100 检测总RNA 的质量浓度、纯度和完整性,结果表明,乳腺样本R N A 完整值(R N A integrity number,RIN)≥6.8,质量浓度、纯度和完整性较好,检测结果为A 级,达到测序样品质量要求,可用于后续转录组测序。
2.2.2 测序数据总体分析 在本研究中,8 个乳腺测序文库共计得到477.47 M Raw reads,每个样品的Raw reads 总数均达47 M 以上,每个样品保留下来的Clean reads 最少约占总数的97.33%。每个乳腺组织样的Clean bases 总数均大于7.76 G,检测碱基含量分布发现,GC 含量为49.3%~52.23%,Q20≥97.14%,Q30≥92.24%,说明测序质量较高,可进行后续分析。
2.2.3 差异表达分析 通过Cuffdiff 同种组织各组间差异表达基因进行筛选,发现乳腺组织共筛选出235 个显著差异表达的mRNA,其中上调表达的有64 个,下调表达的有171 个。筛选条件为P<0.05,|log2(Fold change)|>1(Fold change 表示差异倍数),乳腺组织差异基因火山图如图2所示。
对乳腺235 个差异表达基因的表达量做FPKM(Fragments per kb per million reads) 层次聚类(Hierarchical clustering)分析后得到差异基因聚类图,FPKM 指每百万fragments 中来自某一基因每千碱基长度的fragments 数目,同时考虑了测序深度和基因长度对fragments 计数的影响。如图3 所示,可以看出饲粮处理后乳腺组间差异基因的聚类明显分开,8 个样本聚成2 个类别,对照组样本和处理组样本分别聚为一类,说明这些聚集基因可能具有相似的功能注释或处于相同的代谢通路,各组组内样本之间也有明显的差异。
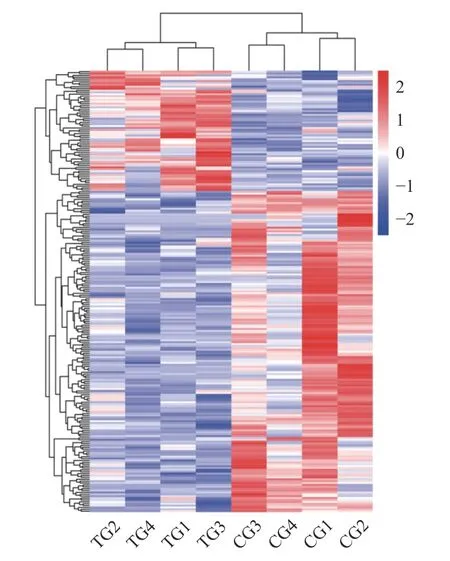
图3 乳腺组织差异表达基因热图Fig.3 Heat map of differentially expressed genes in mammary gland tissue
2.2.4 测序结果的qPCR 验证 对差异表达mRNA 进行荧光定量PCR,以验证测序结果的准确性。选取乳腺中参与脂质调控的关键基因包括FASN(脂肪酸合成酶)、SREBF1(固醇调节元件结合转录因子),另外又随机选取4 个基因在乳腺中进行验证,包括SCGB1D、PLA2G16、RNASE1和BICDL1。如图4 所示,实时荧光定量PCR 结果与测序结本基本一致,说明测序结果可靠性较高。
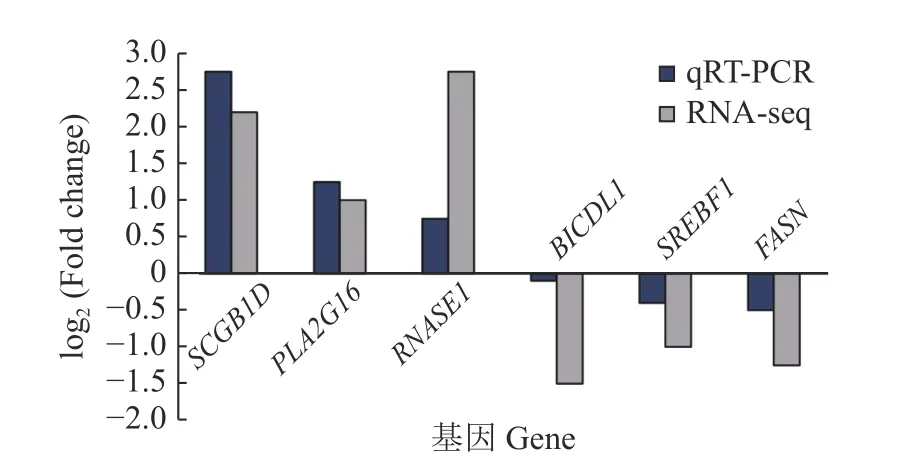
图4 乳腺组织部分差异表达mRNA 荧光定量PCR 验证Fig.4 Fluorescence quantitative PCR validation of partial differentially expressed mRNA in mammary gland tissue
2.2.5 mRNA 功能富集分析 GO 分析结果表明,筛选出的乳腺差异表达基因显著富集到609 个GO条目(P<0.05),其中包含454 个生物学过程条目、111 个分子功能条目和44 个细胞成分条目。图5为乳腺差异mRNA 显著富集的前30 个GO 条目。对差异表达基因进一步富集分析发现,上调基因主要与炎症反应、氨基酸代谢和有机氮化合物代谢等过程有关。下调基因主要与脂质代谢、脂质生物合成、脂肪细胞分化、信号调节以及细胞的增殖、分化、发育等过程有关。
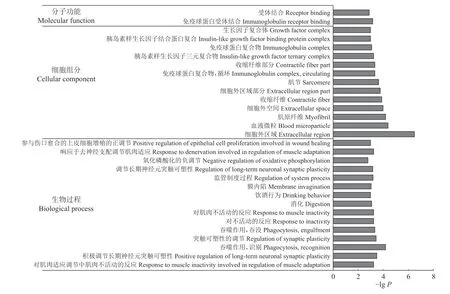
图5 乳腺差异mRNA 显著富集的前30 个GO 条目Fig.5 The first 30 GO terms with significant enrichment of differential mRNAs in mammary glands
对差异表达基因进行KEGG 通路分析(图6)发现,共有20 个基因之间有互作关系,而乳腺上调差异基因显著富集到6 条通路上(P<0.05),包括核糖体、亚油酸代谢和花生四烯酸代谢等;下调差异基因显著富集到14 条通路上(P<0.05),包括轴突导向、钙信号通路、原发性免疫缺陷、NF-κB 信号通路、泛酸和CoA 生物合成以及用于IgA 生成的肠道免疫网络等。

图6 乳腺差异表达基因KEGG 富集图Fig.6 KEGG enrichment map of differentially expressed genes in mammary glands
筛选出的差异显著基因中,FCGR1A、SAA3、NLRC4和CATHL5参与炎症反应和免疫反应;CD40LG和CCL21参与NF-κB 信号非经典通路;FASN、SREBF1、MLXIPL、HTR2B、ETNK2、FGFR4和VDR参与脂质生物合成过程;CCL21、CYP1A2、LRAT和TYRP1参与脂质代谢过程调节;SREBF1、VDR和FGFR4参与调节类固醇代谢过程;AGAP2、NEURL1、IGFBP5、FASN和VDR参与乳腺发育过程;HTR2B、ETNK2和SREBF1参与甘油脂合成过程。
2.2.6 奶牛泌乳性能与差异基因表达的相关分析 本研究选取了差异显著性TOP 10 的基因,并根据功能富集分析筛选了与氨基酸代谢、脂质代谢、炎症反应等过程相关的差异表达基因,将乳腺中筛选出的差异基因分别与奶牛的平均泌乳性能数据做了相关性分析,由表2 可知,乳腺中,RNASE1、Novel02780和TECTB表达与乳脂率和乳脂产量呈显著负相关(P<0.05 或P<0.01);GSTA2、FCGR1A、SAA3、RNASE1、Novel02780和NLRC4表达与DMI 呈极显著负相关(P<0.01),其中FCGR1A、SAA3和NLRC4参与炎症与免疫反应;GPA33、LRRC75A、GPR37L1、COL26A1、APLN、SREBF1、VDR、FGFR4、CCL21和NEURL1表达与产奶量和乳脂产量呈显著正相关(P<0.05);FBXO10、HTR2B、TYRP1、CD40LG、AGAP2和IGFBP5表达与乳脂产量呈显著正相关(P<0.05)。

表2 奶牛泌乳性能与部分乳腺差异基因表达的相关分析1)Table 2 Correlation analysis of lactation performance and expression of selected differential genes in mammary glands of dairy cows
3 讨论与结论
3.1 高淀粉高油日粮对奶牛DMI 和泌乳性能的影响
DMI 受日粮适口性、能量水平等多种因素影响。本研究得出,处理组奶牛的DMI 和产奶量在13 和16 d 显著降低。早期研究显示,每天采食高浓度油时,DMI 较低,产奶量也有所下降[10]。但Boerman 等[11]饲喂添加大豆油或大豆脂肪酸馏出物饲粮时发现,奶牛的DMI 下降,产奶量却提高。此外,Oba 等[12]研究发现,在奶牛瘤胃中灌注丙酸来替代乙酸可导致奶牛DMI 下降。本试验处理组日粮中添加了粉碎玉米,较高的淀粉含量可在瘤胃中产生较多的丙酸,参与到肝脏的氧化代谢,抑制动物机体采食,进而导致奶牛DMI 下降。
产奶量受生理、环境、营养、疾病等多种因素影响。在瘤胃健康时,高精料饲粮通常可提高奶牛的产奶量,但是当瘤胃健康受损时,高精料饲喂则会降低奶牛的产奶量[13]。另一方面,泌乳后期奶牛在妊娠过程中食欲较差,采食量下降,产奶量也会降低[14]。本试验得出,诱导初期处理组奶牛的产奶量略高于对照组,之后呈下降趋势,可能与DMI 的显著降低及瘤胃健康有关。
本试验处理组奶牛饲粮添加粉碎玉米和大豆油饲喂7 d 后,乳脂率和乳脂产量开始下降且差异显著。研究表明,用高精料诱导奶牛MFD 时乳脂率显著降低了25%,乳脂产量显著降低了27%[15];饲粮中添加4%(w)豆油或4%(w)胡麻油后,奶牛均出现了乳脂降低现象[16];高淀粉饲粮添加600 g 葵花籽油饲喂2 周以上,可引起奶牛的乳脂合成抑制[17],说明高淀粉高油饲粮可降低乳脂合成,与本试验结果一致。本试验处理组奶牛第13、16 和19 天的乳蛋白、乳糖和非脂固形物含量显著提高。然而部分研究显示,饲喂高精料植物油饲粮时并未观察到乳糖、乳蛋白和非脂固形物有显著影响[18],与本试验结果不一致,这可能与引起乳脂降低的饲粮组成及其对产奶量的影响不同有关。有研究表明,乳蛋白含量与饲粮的能量水平有关,适当增加饲粮精粗比可以提高乳蛋白含量[19]。乳糖含量增加,可能是由于奶牛采食高淀粉日粮后,饲粮中的过瘤胃淀粉进入肠道生成葡萄糖量相应增加[20]。
3.2 高淀粉高油日粮对奶牛乳腺组织脂代谢相关基因表达的影响
对乳腺差异mRNA 进行富集分析后发现,上调mRNA 显著富集到的GO 条目主要与炎症反应和免疫反应、氨基酸代谢和有机氮化合物代谢有关;下调mRNA 显著富集到的GO 条目主要与脂质生物合成与脂质代谢、细胞增殖分化与细胞发育以及乳腺发育有关。
3.2.1 高淀粉高油日粮影响奶牛乳腺脂质代谢的作用机制 通过筛选得到与炎症反应和免疫反应相关的基因有FCGR1A、SAA3、NLRC4和CATHL5。FCGR1A 是免疫球蛋白FcγR 受体家族的主要成员之一,可参与调节免疫反应,它可通过激活NF-κB 信号传导增加NLRP3 炎症小体的形成以及炎性细胞因子(IL-1β 和IL-18)的释放,从而加剧免疫炎症[21]。SAA3(血清淀粉样蛋白A3)是奶牛乳腺上皮细胞(Mammary epithelial cell,MEC)分泌的主要血清淀粉样蛋白A (Serum amyloid A,SAA)亚型,在炎症期间于大多数MEC 中高水平表达,能在宿主防御中发挥作用。此外,研究表明,SAA3 也能由脂肪细胞产生,且SAA3 参与奶牛分娩前后的炎症反应调节[22]。NLRC4 在幽门螺杆菌感染中,可通过控制IL-18 的产生来降低宿主的免疫反应,从而导致炎症[23]。CATHL5 是抗菌肽家族的成员,具有抗菌活性和炎症反应调节功能,还可触发宿主的特定防御反应,对哺乳动物的免疫反应有重要作用[24]。除此之外,CD40LG和CCL21基因表达下调并显著富集到NF-κB 信号非经典通路,CD40LG 在免疫应答中起主要作用,能诱导激活NF-κB[25];CCL21 是一种趋化因子,对淋巴组织内原始淋巴细胞的返回和运输具有重要作用[26]。本研究中,CCL21表达与乳脂产量之间具有高度正相关性,因此CCL21表达量的减少可能是乳脂产量降低的影响因素之一。
3.2.2 高淀粉高油日粮影响奶牛乳腺的脂质调控 本试验结果显示,多个基因参与脂质合成、代谢等生物学过程。
奶牛饲喂高淀粉高油饲粮后,乳脂合成降低与关键脂质合成基因(FASN)和转录因子(SREBF1、MLXIPL)的表达下调有关。FASN 是参与脂肪酸合成的关键酶,其转录本显著降低直接影响乳腺脂肪酸的从头合成。SREBF1 可通过调控涉及胆固醇和脂肪代谢的众多基因的转录来调节脂质的生物合成和脂肪形成[27]。MLXIPL 又被称为碳水化合物反应元件结合蛋白(ChREBP),是乳腺中调节脂肪生成的另一个转录因子;研究表明MLXIPL 对脂肪生成有积极作用,对于脂肪酸和体内甘油三酯合成的协调控制至关重要[28]。本研究发现HTR2B、ETNK2均富集到磷脂合成、甘油磷脂合成的GO 条目中。HTR2B(5-羟色胺受体2B)被发现于研究人类脂肪形成和脂肪储存的新基因中,当HTR2B表达显著下调12.25 倍时,脂肪细胞表现出脂质积累增加;用HTR2B拮抗剂处理过的细胞进行验证发现,细胞中性脂质水平显著增加[29]。ETNK2(乙醇胺激酶2)是乙醇胺激酶(EKI)家族成员,EKI 通过二磷酸胞苷(CDP) 乙醇胺途径催化磷脂酰乙醇胺(PE)生物合成的第1 步,该酶参与甘油磷脂代谢。FGFR4 是跨膜酪氨酸激酶受体,在调节肝胆汁酸、全身脂质代谢、葡萄糖稳态以及细胞信号传导中起重要作用[30]。VDR(维生素D3 受体)是配体诱导型转录因子的核激素受体超家族的成员,VDR 直接与NLRP3 相互作用,抑制NLRP3 炎症细胞的组装,减少NLRP3 介导的IL-1β 和IL-18 的分泌[31],提示VDR 可能是一种潜在的抗炎因子。
LRAT(卵磷脂视黄醇酰基转移酶)催化视黄醇转化为视黄酯,在人乳腺组织中的研究表明LRAT在人的乳房生理中起重要作用[32]。CYP1A2 是细胞色素P450 酶超家族的成员之一,CYP1A2 可存在于内质网囊泡产生的脂质微区中但似乎并不影响其形成。TYRP1(酪氨酸酶相关蛋白1)参与黑色素生物合成途径,Randhawa 等[33]研究发现黑色素生成相关基因在内脏脂肪组织中的表达,首次证明了黑色素的生物合成途径在脂肪组织中起作用。此外,本试验相关性分析结果显示TYRP1表达与乳脂产量之间有极显著的正相关性。
本研究中AGAP2、NEURL1和IGFBP5参与了乳腺发育过程,说明高淀粉高油饲粮对奶牛乳腺发育有一定的影响。AGAP2 参与细胞凋亡、细胞存活和受体运输相关的信号通路[34]。NEURL1(神经化E3 泛素蛋白连接酶1) 能通过泛素的赖氨酸残基K27 导致其蛋白酶体介导的降解来调节PDE9A 蛋白水平,从而可能对细胞cGMP 水平产生积极影响。IGFBP5(胰岛素样生长因子结合蛋白5)对调节乳腺发育和泌乳后复性过程中的细胞凋亡和细胞增殖有重要作用[35]。
以上基因中,CCL21既参与脂质代谢调节也参与了炎症调节,而由于之前在SREBP1的启动子区域中发现了NF-κB 的结合位点[36],因此推测炎症信号通路可能影响了脂质代谢的调节,有待进一步验证。结合差异表达基因与泌乳性能的相关性分析,基因FGFR4、VDR、HTR2B、CCL21和TYRP1虽不在常规的脂肪代谢过程中,但其表达量与乳脂产量具有较高的正相关,对乳腺发育和乳腺脂质代谢具有重要调控作用,上述基因表达量的下调可能是引起乳脂含量降低的重要因素,因此可作为研究奶牛MFD 脂质代谢调控的候选基因。
3.3 结论
通过饲喂高淀粉高油日粮,降低了奶牛的DMI、产奶量、乳脂率,提高了乳蛋白、乳糖和非脂固形物含量。测序结果显示,乳腺中参与脂肪合成的mRNA 的表达下调是造成乳脂率和乳脂产量降低的主要因素。结合差异表达基因与泌乳性能的相关性分析,筛选出了乳腺中可能与脂质代谢调控存在紧密联系的基因FGFR4、VDR、HTR2B、CCL21和TYRP1作为研究MFD 的候选基因,可为研究荷斯坦奶牛MFD 的分子机制奠定基础。
