尾巨桉人工林主要生态效应研究*
2024-01-14任世奇杜阿朋顾大形韦振道李昌荣陈健波
任世奇,杜阿朋,胡 刚,顾大形,伍 琪,韦振道,李昌荣,陈健波
(1.广西壮族自治区林业科学研究院,南宁桉树森林生态系统广西野外科学观测研究站,广西南宁 530002;2.中国林业科学研究院速生树木研究所,广东湛江 524022;3.南宁师范大学环境与生命科学学院,广西南宁 530100;4.广西壮族自治区中国科学院广西植物研究所,广西桂林 541006)
桉树是桃金娘科(Myrtaceae)杯果木属(Angophora)、伞房属(Corymbia)和桉属(Eucalyptus)树种的统称,共有800多种[1],天然分布于澳大利亚、印度尼西亚东部岛屿、东帝汶和菲律宾南部[2]。桉树适应能力强、生长速度快、加工用途广、经济价值高,其木材可用于生产人造板和制浆造纸;叶片精油可用于护肤品生产、食品添加;树皮可提炼鞣质。世界范围内有120多个国家和地区引种栽培桉树,面积多达3亿亩[3](1亩≈666.67 m2)。
我国引种栽培桉树已有130多年历史,大规模引种栽培桉树始于1982年中国与澳大利亚的桉树合作示范项目,其中引种驯化最为成功的桉树树种当属尾叶桉(Eucalyptusurophylla)和巨桉(E.grandis),利用这两个桉树树种杂交开发具有自主知识产权的尾巨桉(E.urophylla×E.grandis)和巨尾桉(E.grandis×E.urophylla)杂交种,这两个杂交种是近20年我国种植范围最广、面积最大的桉树品种,种植范围覆盖广西、广东、海南、福建等省区,种植面积超过8 000万亩,以此成就了全国桉树木材年产量占全国木材年产量40%以上的贡献。广西是我国桉树产业化发展最好的省区之一,2021年广西桉树人工林面积达4 550万亩,居全国第一位[4]。
自2000年以来,随着桉树人工林种植面积的持续扩大,其大面积连片纯林、短轮伐期等经营方式带来的生态效应受到广泛地关注和质疑。已有研究表明,通过将人工纯林调整为单一树种占较大比率的混交林可提高林地的经济回报和提供更多元化的产品[5],降低病虫害发生率[6],改善土壤理化性质[7]。杨承栋[8]深入总结单个树种针叶纯林连作效益并提出:有群落结构的混交林是维护、恢复和提高森林土壤功能,提高森林土壤生产力,实现可持续经营的关键。为真实了解桉树人工纯林及其混交林的生态效应,客观认知桉树人工纯林及其混交林生长过程对自然生态环境的影响,本研究围绕桉树人工纯林和混交林的生态系统组成、结构与功能,通过对实施不同经营模式和技术措施形成的不同类型桉树人工林进行定位观测研究,初步评价桉树人工林的主要生态功能,以期为桉树人工林生态和经济综合效益的发挥提供生态高效经营技术方案,最终为实现桉树人工林的科学化发展奠定基础。
1 材料与方法
1.1 研究区域
本研究依托南宁桉树森林生态系统广西野外科学观测研究站(以下简称南宁桉树野外站,包括南宁站点、柳州站点)和湛江桉树森林生态系统国家定位观测研究站,并以桂林、北海两个地区作为植物多样性调查辅助样点。观测范围跨越北纬21°-25°,覆盖桂北丘陵山地、桂中盆地平原、桂东南-粤西沿海台地3个区域的典型桉树人工林观测样带。气候条件分属中亚热带湿润季风气候、南亚热带季风气候和北热带海洋性季风气候,其地带性植被分别为常绿阔叶林、季风常绿阔叶林和季节性雨林;土壤类型以红壤、赤红壤、砖红壤为主。
1.2 方法
1.2.1 尾巨桉单木、林分的水分消耗及林区水质观测
(1)尾巨桉单木蒸腾耗水量。以南宁桉树野外站(南宁站点)的中龄尾巨桉为研究对象,利用树干茎流仪(TDP,Dynamax公司,美国)定位观测尾巨桉单木的树干液流密度,通过伐倒木圆盘测定尾巨桉单木边材面积,利用Granier公式[9]量化分析尾巨桉单木林冠蒸腾水分,即尾巨桉单木蒸腾耗水量[10]。
(2)尾巨桉林分蒸散耗水量。以南宁桉树野外站(南宁站点)的中龄尾巨桉林分为研究对象,使用自动气象站(CR1000,Campbell公司,美国)观测林外大气降水,利用坡面径流场量化地表水分流失量,使用树干茎流仪、自动雨量计(UA-003-64,Hobo公司,美国)分别采集树干茎流量和穿透水量,通过林外降水与林内穿透水差值获取林冠截留量。根据水量平衡方程由上述样地尺度水分循环分量计算出尾巨桉林分蒸散耗水量。此外,应用彭曼模型(Penman-Monteith)估算尾巨桉林分蒸散耗水量,并将水量平衡方程实测的尾巨桉林分蒸散耗水量与彭曼模型模拟结果进行比较,评价彭曼模型模拟尾巨桉林分蒸散耗水量的适用性[11]。
(3)尾巨桉活立木根系吸水来源。使用同位素比率质谱仪(MAT 253,Thermo Fisher Scientific 公司,美国)量化一代植苗林、二代萌芽林(一次萌芽林)、三代萌芽林(二次萌芽林)、四代萌芽林(三次萌芽林)4种不同连栽代次尾巨桉纯林的土壤水、林分周边地下水、林分外降水等水环境的δD 比率,使用IsoSource多元混合模型分割量化不同水环境对尾巨桉根系吸水的贡献[12,13],剖析不同连栽代次尾巨桉根系对土壤水、地下水、降水等水环境的吸收策略。
(4)尾巨桉林区地表水水质演变。在尾巨桉三代萌芽纯林(Three generations of pureE.grandis×E.urophylla,Eck)、尾巨桉采伐迹地丢荒(Cutover land ofE.grandis×E.urophycla,WL)、尾巨桉+灰木莲水平带状混交(Horizontal mixed stand ofE.grandis×E.urophyclaandMagnoliaceaeglanca,EMH)、尾巨桉+灰木莲垂直带状混交(Vertical mixed stand ofE.grandis×E.urophyclaandM.glanca,EMV)、尾巨桉+灰木莲株间混交(Interplant mixed stand ofE.grandis×E.urophyclaandM.glance,EMI)、灰木莲纯林(PureM.glancastand,Mck)6种土地利用类型中分别安装地表水采集装置,连续7个月采集6种土地利用类型的地表水水样,分析地表水水质演变特征。
1.2.2 尾巨桉林地土壤养分循环观测
(1)尾巨桉植苗纯林凋落物量及分解归还。连续12个月按月收集5年生、7年生和9年生尾巨桉植苗纯林凋落物,以枝条、叶片、果皮分类测定生物量及养分含量,同时将枝条与叶片按不同比率混合分装在720个分解袋中并随机放置于该林分的地表,每月随机抽取3个分解袋称重并化验袋中剩余凋落物的养分含量[14]。
(2)尾巨桉三代萌芽纯林及改培混交林的土壤理化性质及微生物群落结构。采集不同龄组尾巨桉纯林、尾巨桉混交林的土壤样品,分析其土壤微生物群落结构,剖析不同林分类型对土壤地力维持机制[15]。
1.2.3 尾巨桉人工林生物多样性调查
在桂林、南宁、北海3个不同纬度地区选择1-3年、4-6年和7-9年3个龄组尾巨桉人工林并区划60个样方,调查尾巨桉林下植物的种类、多度、盖度等指标,解析尾巨桉人工林林下植物多样性特征[16]。
1.2.4 桉树人工林碳储量及林冠CO2交换量观测
以第九次(2018年)全国森林资源清查结果为数据源,利用政府间气候变化专门委员会(IPCC)推荐的生物量估算模型估算广西桉树人工林的碳储量[17]。依托南宁桉树野外站(柳州站点),通过在综合观测塔上安装开路式涡度相关系统(Li-7500A,LIcor Co.,Ltd.,USA)并连续3年观测鹿寨区域的中龄尾巨桉人工林林冠与大气交界面的CO2通量,估算尾巨桉人工林生态系统的碳汇能力。
2 结果与分析
2.1 尾巨桉单木、林分的蒸腾耗水量及林分水质演变特征
2.1.1 尾巨桉单木蒸腾耗水量
尾巨桉单木日均蒸腾耗水量为1.32 mm,其中春、夏、秋、冬四季典型晴天的尾巨桉单木日均蒸腾耗水量分别为1.58、1.61、1.23、0.84 mm,其差异表现在早晨液流提速时刻、中午液流峰值到达时刻、傍晚液流低速时刻和白天液流持续时间。在尾巨桉种植密度为83株/亩的情况下,单位土地面积的尾巨桉单木全年蒸腾耗水量约为349 mm,约占当年降水量的27%。
2.1.2 尾巨桉林分蒸散耗水量
彭曼模型估算结果显示,中龄尾巨桉人工林的日均林分蒸散耗水量3.5 mm,季节平均蒸散耗水量289 mm (其中夏季蒸散耗水量445 mm、冬季蒸散耗水量117 mm),全年蒸散耗水量占当年降水量的82%。同时,全年的每日土壤相对含水量处于适宜或偏湿状态,土壤水分不是中龄尾巨桉人工林蒸散耗水的限制因子。
2.1.3 尾巨桉人工林水分循环组成在中龄尾巨桉人工林叶面积指数约为1.82的背景下,林分水量平衡分量特征表现为降水输入再次分配的林冠截留量约占22%,穿透水量约占77%,树干茎流量约占1%;水分输出分量特征表现为林冠截留蒸发量约占22%,林冠蒸腾量约占27%,林下蒸散量约占46%,土壤入渗和地表径流约占5%。
2.1.4 尾巨桉活立木根系吸水来源
一代植苗尾巨桉根系吸水主要源于0-60 cm 土层,该层水分贡献占82%,其中0-10 cm 土层的水分贡献最大,占37%。二代萌芽尾巨桉根系吸水也主要源于0-60 cm 土层,该层水分贡献占87%,根系对该土层的水分吸收较均匀。三代萌芽尾巨桉根系吸水主要源于0-80 cm 土层,该层水分贡献占84%,根系对该土层的水分吸收较均匀。四代萌芽尾巨桉根系吸水主要源于0-40 cm 土层,该层水分贡献占85%,其中0-20 cm 土层土壤水贡献最大,占39%。综上,尾巨桉人工林的根系吸水主要源于土层深度80 cm 以内的土壤水。
2.1.5 尾巨桉人工林地表水水质演变
定位观测结果显示,尾巨桉人工林地表水未出现“黑水”现象。尾巨桉人工林地表水的p H 值约为5.8。通过分析尾巨桉纯林不同改培模式地表水的化学需氧量(COD)和五日生化需氧量(BOD5)(图1)可知,尾巨桉纯林和灰木莲纯林地表水的化学需氧量变化相似,二者平均值约为55 mg·L-1,最大值为120 mg·L-1;伐桉迹地丢荒地表水的化学需氧量最小,平均值仅为35 mg·L-1;尾巨桉与灰木莲3种方式混交林地表水的化学需氧量均比尾巨桉纯林的略高,总体平均值小于130 mg·L-1。由五日生化需氧量变化规律可知,尾巨桉纯林及其不同改培模式林分地表水的五日生化需氧量在5月份时最高,其波动范围为40至100 mg·L-1,高于其他月份(<60 mg·L-1)。尾巨桉纯林及其不同改培模式林分地表水的p H 值、化学需氧量、五日生化需氧量3 项指标均达《农田灌溉水质标准》(GB 5084-2021)规定的限定值。
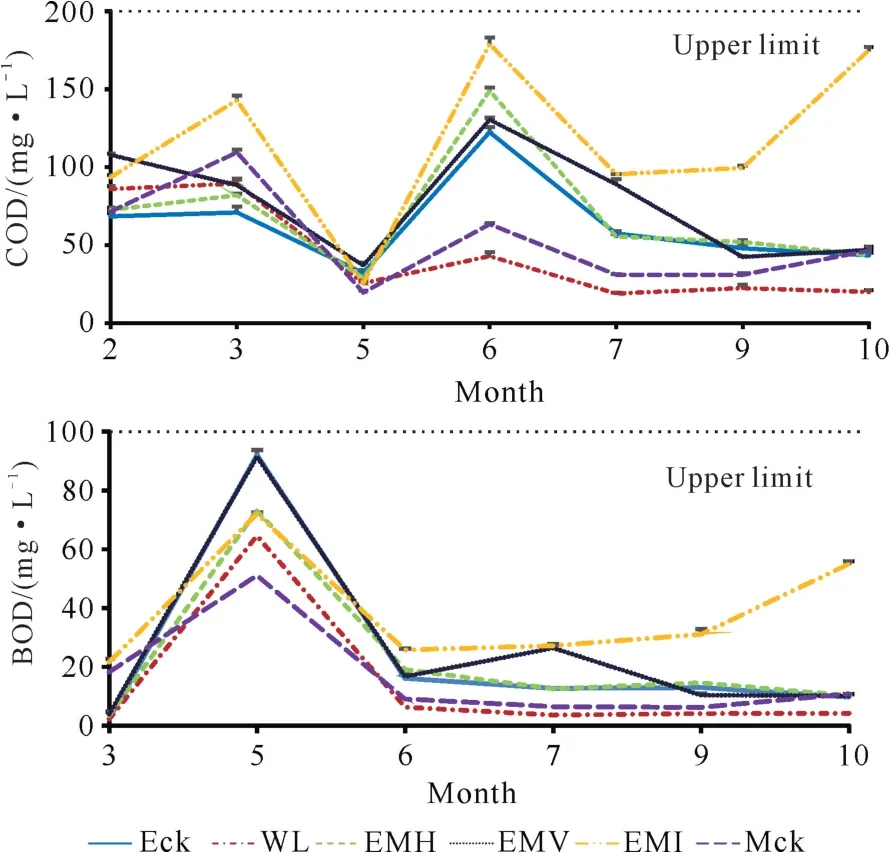
图1 尾巨桉纯林及其不同改培模式地表水的化学需氧量和五日生化需氧量Fig.1 Surface water COD and BOD5 of different forest types shifting from E.urophylla×E.grandis
2.2 尾巨桉纯林、混交林的土壤性质变化特征
2.2.1 尾巨桉植苗纯林凋落物量及分解养分归还特征
通过对5年生、7年生、9年生3个成熟期的尾巨桉植苗纯林凋落物量及其养分含量连续12次的月度监测,发现叶片是凋落物的主要成分,占凋落物总量的47%-54%。明确凋落物养分归还以氮元素为主,归还量55.48-58.13 kg·hm-2;其次为钾元素,归还量16.48-20.37 kg·hm-2;磷元素的归还量最小,仅2.61-3.66 kg·hm-2。
2.2.2 尾巨桉三代萌芽纯林改培混交林的土壤化学性质特征
由表1可知,尾巨桉纯林、尾巨桉采伐迹地丢荒、尾巨桉+灰木莲株间混交林、灰木莲纯林4种林分的土壤化学性质差异显著。与尾巨桉采伐迹地丢荒比较,尾巨桉纯林土壤的有机质及各养分元素含量显著下降,然而,改培的尾巨桉+灰木莲株间混交林的土壤化学性质有显著改善,且混交林的土壤有机质、全氮、速效氮含量显著提高。值得注意的是尽管尾巨桉+灰木莲株间混交林的速效钾、有效镁含量(39.47、7.96 mg·kg-1)显著高于尾巨桉纯林,但是均显著低于尾巨桉采伐迹地丢荒和灰木莲纯林。同时,尾巨桉采伐迹地丢荒和灰木莲纯林的土壤p H 值显著提升,土壤酸度降低。
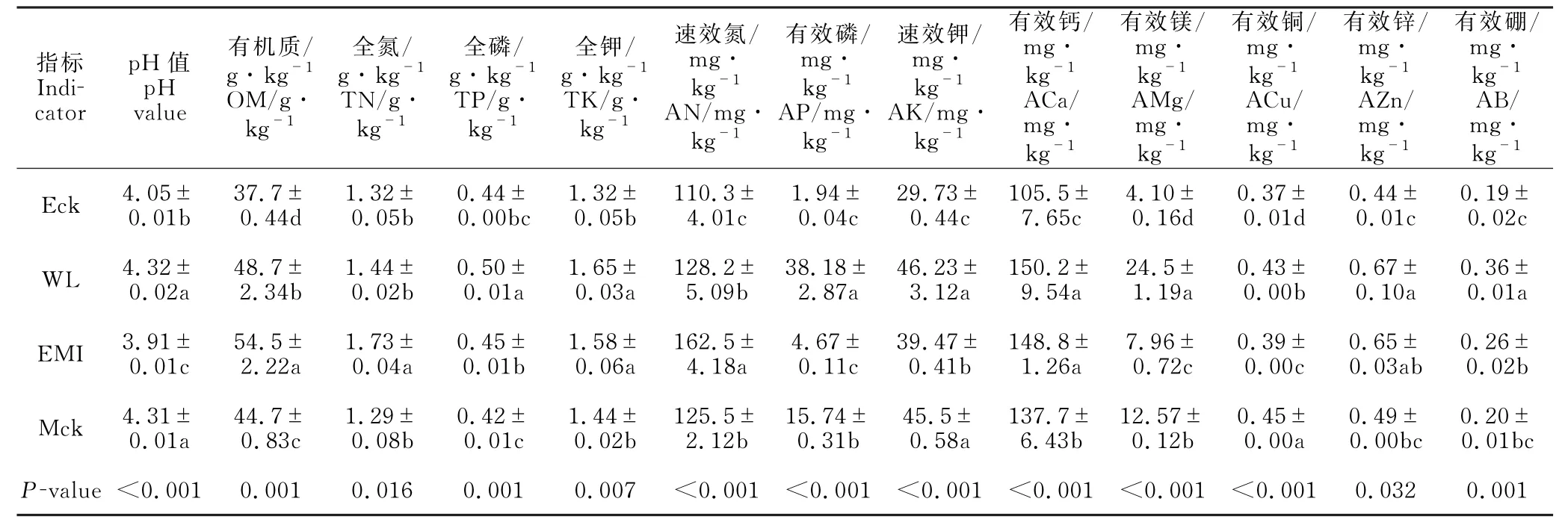
表1 尾巨桉纯林不同改培模式林分的土壤化学性质Table 1 Soil chemical property of different types shifting from E.urophylla×E.grandis
2.2.3 尾巨桉三代萌芽纯林改培混交林的土壤物理性质特征
由图2可知,尾巨桉纯林、尾巨桉采伐迹地丢荒、尾巨桉+灰木莲株间混交林、灰木莲纯林4种林分的土壤容重差异不显著,总体表现为尾巨桉纯林>尾巨桉采伐迹地丢荒>尾巨桉+灰木莲株间混交林>灰木莲纯林,多重比较显示尾巨桉纯林与灰木莲纯林呈极显著差异。不同林分的土壤质量含水量呈显著差异,其中尾巨桉+灰木莲株间混交林的最高,尾巨桉纯林的最低。不同林分之间的土壤最大持水量表现为显著差异,其中灰木莲纯林的最大,其极显著高于尾巨桉纯林,而灰木莲纯林与尾巨桉+灰木莲株间混交林、采伐迹地丢荒之间的差异不显著。4种林分之间的土壤毛管持水量和最小持水量变化规律一致,总体差异不显著,从大到小顺序依次为尾巨桉+灰木莲株间混交林>灰木莲纯林>尾巨桉采伐迹地丢荒>尾巨桉纯林。
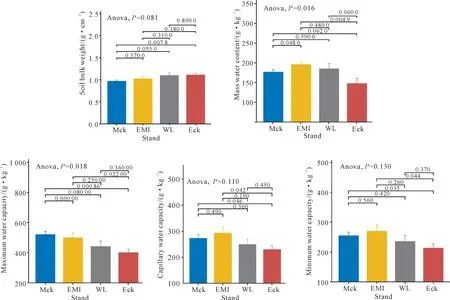
图2 尾巨桉纯林不同改培模式林分的土壤物理性质Fig.2 Soil physical properties of different types shifting from E.urophylla×E.grandis
2.2.4 尾巨桉三代萌芽纯林改培混交林的土壤微生物群落结构特征
尾巨桉纯林改培为混交林后,土壤细菌、真菌的群落结构发生显著变化。灰木莲纯林、尾巨桉采伐迹地丢荒2种林分的土壤细菌群落多样性和丰富度均显著高于尾巨桉+灰木莲株间混交林和尾巨桉纯林;尾巨桉纯林、灰木莲纯林、尾巨桉采伐迹地丢荒、尾巨桉+灰木莲株间混交林4种林分之间的土壤真菌群落多样性差异不显著,但真菌丰富度表现为灰木莲纯林>尾巨桉采伐迹地丢荒>尾巨桉+灰木莲株间混交林>尾巨桉纯林。
2.3 尾巨桉人工林植物物种多样性特征
2.3.1 尾巨桉人工林的林下植物物种情况
通过样方调查发现,60个样方共记录2 482株植物,隶属58科101属142种(被子植物127种、蕨类植物13种、裸子植物2种)。其中,禾本科(Poaceae)植物最多,有9属11种;其次为菊科(Asteraceae)植物,有7 属8 种;茜草科(Rubiaceae)植物有5 科7属。单科单种有28 科28 属,占调查样方总科数的48.28%,包括闭鞘姜科(Costaceae)、番荔枝科(Annonaceae)、钩吻科(Gelsemiaceae)、夹竹桃科(Apocynaceae)、金丝桃科(Hypericaceae)等。101 属中有38属为单种属,占调查样方总属数的37.62%。此外,通过广泛调查收集广西桉树林下植物物种组成发现,广西桉树林中有维管植物120 科369 属584 种(其中被子植物101科337属529种,蕨类植物15科28属49种,裸子植物4科4属6种),分别占广西维管植物科、属、种总数的38.83%、18.35%、6.37%。桉树林中植物以灌木(201种)为主,其次为草本(186种)和乔木(106种),藤本(91种)最少。从区系成分看,科包括8个分布区正型,7个变型,属包括14个分布区正型,15个变型,科、属的R/T 值(热带成分/温带成分)分别为3.20和5.37。
2.3.2 尾巨桉人工林的林下植物物种多样性变化特征
尾巨桉人工林的林下灌木层物种α多样性随林龄变化的规律与草本层的相反,其中灌木层物种α多样性整体上随林龄增长而增加,当尾巨桉人工林为7-9年生时,灌木层物种α多样性最大,表明7-9年生的尾巨桉人工林林下灌木层物种分布较均匀;草本层物种α多样性整体上随林龄增长而降低,当尾巨桉人工林为7-9年生时,草本层物种α多样性最小,表明草本层物种丰富度随林龄增长而降低,尾巨桉人工林4-6年生时的草本层物种分布较为均匀。不同林龄尾巨桉人工林的林下草本层彼此之间的相似系数随林龄增长而升高。广西尾巨桉人工林的林下植物群落具有明显的垂直地带性,其中海拔、土壤全钾是影响其分布的主要因子,其次尾巨桉林分的郁闭度很大程度上也影响着林下植物的分布。
2.4 桉树人工林的固碳能力
根据第九次(2018年)全国森林资源清查结果显示,广西桉树人工林面积256万公顷,其中以幼龄桉树人工林的面积最大,占45.23%;以中龄桉树人工林的蓄积量最大,占总蓄积量1.09 亿立方米的50.48%。基于南宁桉树野外站(柳州站点)于2018-2020年连续3年观测鹿寨区域的中龄尾巨桉人工林林冠与大气界面CO2净交换量(NEE)表明,尾巨桉人工林白天为碳汇,夜间为碳源,夏季和秋季吸收固定CO2的能力最强,该区域中龄尾巨桉人工林平均每日净固定CO2约为11.0 g·m-2,转化为每年净固定碳约为10.9 t·hm-2。
3 讨论
3.1 桉树人工林蒸腾耗水特征
通过表2列举的几个主要造林树种的单木蒸腾耗水量发现,4年生尾巨桉在湿季的蒸腾耗水量仅为2.0 L·d-1,比毛白杨的还少;5年生和6年生的尾巨桉在湿季的蒸腾耗水量为8.0-10.2 L·d-1,少于湿加松、柳杉等树种;10年生尾叶桉和11年生粗皮桉的蒸腾耗水量也小于马尾松、木荷、锥栗等树种,由此说明桉树人工林的蒸腾耗水量并未显著高于其他树种。
3.2 尾巨桉人工林对水质的影响
根据分析结果,尾巨桉纯林及其不同改培模式林分地表水的p H 值、化学需氧量、五日生化需氧量3项指标均达《农田灌溉水质标准》(GB 5084-2021)规定的限定值。同时,2022年7月18日,中华人民共和国生态环境部发布的2022 年第二季度(4-6月)和1-6月全国地表水环境质量状况显示,在1-6月全国地级及以上城市中,排前10名的城市依次是柳州市、黔东南苗族侗族自治州、桂林市、崇左市、丽水市、嘉峪关市、河池市、梧州市、衢州市、百色市,其中第1名柳州市、第3名桂林市、第4名崇左市、第7名河池市、第8名梧州市和第10名百色市均属于广西,证实广西具有优质的地表水资源。根据广西桉树人工林栽培区分布情况,地表水水质排名第1的柳州市、第4的崇左市、第7的河池市、第8的梧州市和第10的百色市均是广西桉树人工林的主栽区,这反映出当前桉树人工林的经营方式未降低地表水水质,也验证了通过延长桉树主伐年龄,降低人为活动对桉树人工林的干扰强度,同时引入乡土树种将桉树纯林改培为混交林等科学经营措施可以提升桉树人工林的生态功能,更好发挥桉树林的生态效益。
3.3 尾巨桉人工林对土壤理化性质的影响
3个年龄序列成熟期尾巨桉人工林凋落物量及其养分含量的12次连续监测结果显示,凋落物以氮元素归还为主,凋落物养分归还总量随林龄增长而逐渐增加,建议适当延长尾巨桉人工林的培育周期有利于凋落物分解养分还土。通过将尾巨桉三代萌芽纯林改培为尾巨桉与灰木莲混交林,其土壤的物理性质、化学性质和微生物群落结构均得到显著改善,与Li等[26]的研究结果一致。
3.4 尾巨桉人工林的林下植物物种多样性变化规律
广泛调查广西尾巨桉人工林的林下植物物种组成发现,维管植物有120科369属584种,其中以灌木为主,达到201种;其次为草本植物,有186种;乔木106种;藤本植物最少,仅有91种。任世奇等[27]也通过植物样方调查和相关文献资料发现,桉树人工林林下植物有190种,其中木本植物32科67属103种,藤本植物16科23属26种,草本植物22科53属61种。与此同时,对60 个调查样方的研究分析发现,桉树人工林的林下灌木层物种多样性整体随林龄增长而增加,其中在7-9年生的桉树林下的灌木植物多样性最丰富;草本层物种多样性整体随林龄增长而降低,其中在7-9年生的桉树林下的草本植物多样性最低。通过比较桉树人工林与其他树种的林下植物多样性(表3)发现,成熟桉树人工林的林下植物物种丰富度并不低,且总体上灌木多样性比草本多样性高。
3.5 桉树人工林的固碳功能特征
我国主要造林树种的碳汇能力年均约2.2 t·hm-2(图3),而桉树人工林每年每公顷可吸收固定碳约4.0 t[33],其碳汇能力远大于其他主要造林树种。对鹿寨县黄冕乡的中龄尾巨桉人工林固碳效率的评估结果为10.9 t·hm-2·a-1,因此,应挖掘桉树人工林碳汇潜力,以更大程度地发挥桉树人工林的固碳释氧功能,为国家实施双碳战略应对CO2减排,实现2060年碳中和目标发挥重要作用。

图3 我国主要造林树种的碳汇能力Fig.3 Carbon sink capacity of chief tree species in our country
4 结论
针对桉树生长的水分消耗,桉树林对地表水水质的影响,桉树林地土壤理化性质、土壤酶活性、微生物群落结构,桉树林下植物物种多样性,以及桉树的固碳增汇能力等方面内容开展了较系统的研究工作,总结出了阶段性研究结论:(1)桉树在生长过程中的单木蒸腾并未消耗大量水分;(2)桉树未显著降低地表水水质;(3)桉树纯林改培混交林可显著提升林地的土壤质量;(4)桉树林下植物物种多样性丰富度随林龄增长呈增加趋势;(5)桉树的碳汇功能强于我国其他主要造林树种。上述结论揭示了桉树种植对生态环境影响的客观规律。目前,广西年生产桉树木材3 000万立方米以上,年采伐量3 000万立方米以上,广西以占全国森林面积的1.4%生产了全国32%的木材,极大地缓解了我国木材的供需矛盾及木材资源供给不足压力,2020年桉树也被列入国家储备林建设树种名录[34]。因此,建议从景观尺度上,适当延长桉树主伐年龄,新造林或萌芽林适当套种乡土树种形成同龄或异龄桉树混交林,同时对林木兼施人工修枝,以此从定向培育关键技术措施上优化桉树经营方案,既利用其速生丰产特性,又充分发挥其生态服务功能,实现桉树的生产力与生态服务功能双增效益,更好地为发展广西林业万亿元绿色产业和建设国家储备林核心基地服务。
