厚壳贻贝几丁质提取及几丁质相关基因筛查
2024-01-09袁俊杰李迈唐菊艳袁孟强李千千王晓娜张美溦陈伟王晓通
袁俊杰,李迈,唐菊艳,袁孟强,李千千,王晓娜,张美溦,陈伟,王晓通
(1.鲁东大学 农学院,山东 烟台 264025;2.烟台市海洋经济研究院,山东 烟台 264003)
几丁质是自然界中广泛分布的一类生物大分子,由D-N-乙酰氨基葡萄糖通过β-(1,4)糖苷键聚合而成[1],根据结构不同,分为α-、β-和γ-几丁质三种类型[2]。几丁质通常不以单体形式存在,其常与蛋白质、色素、碳酸钙等结合形成复合物[2],广泛存在于软体动物贝壳和内骨骼中,在贝壳形成和生物矿化中发挥重要作用。目前,已从乌贼、珠母贝、芋螺、江珧等软体动物中成功提取并鉴定到几丁质成分[3-7],研究发现软体动物几丁质具有良好的生物相容性、抗菌活性[7-8],在生物医学等方面有重要的应用前景。
几丁质作为软体动物贝壳有机组分之一,与其他蛋白质、糖蛋白、多糖等物质组成有机框架,在此基础上进行生物矿化[9]。其中,几丁质合成酶(chitin synthetase,CHS)、几丁质酶(chitinase,CHIA)、几丁质结合蛋白(chitin-binding protein,CBP)等与几丁质框架构建相关的蛋白,均是调控贝壳形成的重要蛋白,参与调控生物矿化[10-14]。在长牡蛎、紫贻贝和大扇贝的贝壳损伤研究中,三种双壳贝类的贝壳修复后,外套膜差异表达蛋白质中均鉴定到几丁质结合相关的结构域[15]。在紫贻贝贝壳修复和生物矿化的转录组分析中,也筛查到大量上调表达的几丁质代谢相关基因,包括几丁质合成酶(CHS)和几丁质酶(CHIA)等[16]。
厚壳贻贝(Mytilus coruscus)是我国重要的海水养殖贝类,具有较高的经济价值[17]。根据已有的研究结果:厚壳贻贝的贝壳包括珍珠质层、肌棱柱层和斜棱柱层[18],并通过蛋白质组学鉴定到60 余种壳基质蛋白[19],但几丁质以及几丁质相关基因在厚壳贻贝贝壳形成中的作用却鲜有报道。本研究从形态和组织学两方面详细观察了厚壳贻贝贝壳与外套膜结构,从厚壳贻贝的角质层和贝壳中提取并鉴定到几丁质,并构建外套膜三褶皱转录组,根据转录组数据筛查到几丁质合成、代谢、结合相关基因,并对三褶皱差异表达基因进行GO 富集分析,筛查出与几丁质功能相关的GO term。本研究有助于了解厚壳贻贝贝壳中有机骨架的成分,为几丁质在贝壳形成和生物矿化中的作用奠定研究基础。
1 材料与方法
1.1 实验样品
实验所用厚壳贻贝材料购于江苏省连云港市,实验前在海水养殖实验室内充氧暂养,暂养海水水温20 ℃,盐度25‰,养殖期间饲喂螺旋藻粉,每天换水一次。
1.2 贝壳及外套膜的形态及组织学观察
将腹缘端贝壳切下部分后,露出白色贝壳,并保留贝壳边缘的角质层,观察贝壳、角质层、外套膜三者生理学结构。将贝壳沿闭壳肌断开,取腹缘端外套膜边缘组织,经固定、脱水、透明、包埋后,在切片机(Leica,RM2255)上切成6 μm厚的切片。切片经脱蜡、复水后,用苏木精-伊红染液(南京建成,D006)染色,后经脱水、透明,用中性树脂封片。最后,在显微镜(Nikon,CI-L)下观察切片。
1.3 贝壳中几丁质的提取
将厚壳贻贝贝壳刷洗干净后,将表面的角质层剥离,烘干后研磨成粉末,放入2%盐酸浸泡30 min,用蒸馏水充分漂洗,高速离心分离得到不溶物并吸干水分。向不溶物加入2 mol/L的NaOH溶液,室温震荡24 h,之后高速离心30 min,蒸馏水清洗至中性,过滤得到棕褐色不溶物。H2O2漂白后,蒸馏水漂洗。60 ℃烘干,得到固体粉末。
将角质层剥离后的贝壳清洗并烘干,也研磨成粉末,并放入2%盐酸浸泡脱钙,漂洗至pH 为中性后,高速离心,向沉淀物加入2 mol/L 的NaOH 溶液,室温震荡24 h。高速离心、洗涤、滤纸过滤,得到脱蛋白不溶物。脱色、漂洗、烘干,得到粉末状固体。
1.4 几丁质提取物的傅里叶红外光谱鉴定
各取0.01 g 几丁质提取物,在NICOLET IS50型傅里叶红外仪器下,经32次全反射扫描在400~4 000 cm-1波数范围内测得吸收峰,使用OMNIC软件绘制吸收峰图。
1.5 基于外套膜转录组的几丁质相关基因鉴定
随机选取3 只厚壳贻贝,分别采集外套膜边缘的外褶皱、中褶皱、内褶皱组织,液氮速冻,放入超低温冰箱保存。提取总RNA 后,抽提mRNA 构建RNA-seq 文库并测序。原始数据经质量过滤后,通过Trinity 软件进行De novo拼接,对组装好的Unigene进行常见功能数据库注释,根据功能注释和BLAST 比对,筛选出几丁质合成、代谢、结合相关的基因,进行表达量(FPKM)分析,并对其蛋白序列进行功能域预测(http://smart.embl-heidelberg.de/)。对内、中、外褶皱间差异表达基因进行GO富集分析,筛查出与几丁质合成、代谢、结合等功能相关的GO term。
2 实验结果
2.1 厚壳贻贝贝壳及外套膜的组织特征
通过形态观察,厚壳贻贝贝壳主要由角质层(贝壳硬质蛋白)和贝壳层(碳酸钙晶体)两部分组成(图1(a)),而外套膜边缘部分主要分为三个褶皱(图1(b)、(c)),外褶皱(of)、中褶皱(mf)以及内褶皱(if),其中外褶皱形态在三个褶皱中最小,其边缘紧紧贴附在贻贝的贝壳边缘,内褶皱靠近其他软组织部,且是三褶皱中形态最大的,其上分布大量外套膜触手,而中褶皱则位于内、外褶皱中间,组织大小也介于二者之间。通过石蜡切片进一步观察,中褶皱又分为一大一小2 层褶皱,靠近外褶皱一侧有1 个非常小的褶皱,与外褶皱共同形成一个沟状结构,此处可分泌贝壳的角质层膜(图1(c)、(d1))。此外,根据组织切片观察,发现三个褶皱的上皮细胞类型也有较大差别:壳膜沟处的外褶皱具有单层柱状上皮细胞,其形态细长,细胞核细长位于基部,细胞质中存在少量褐色的色素(图1(d1));而靠近壳膜一侧的中褶皱(mf1 和mf2)其细胞形态为矮柱状,细胞核椭圆形位于基部,这些上皮细胞中并未观察到棕色的色素(图1(d1)),而在远离壳膜一侧的中褶皱上皮细胞中则可观察到大量棕色色素(图1(d2));内褶皱也具有单层柱状上皮,这些上皮细胞的核细长位于细胞基部,在胞质中可观察到大量深棕色的色素(图1(d3)、(d4))。

图1 厚壳贻贝贝壳及外套膜的形态及组织学观察
2.2 贝壳几丁质提取及FTIR特征
从厚壳贻贝角质层粉末(图2(a))中,提取并鉴定到几丁质(图2(b)):在 3 272 cm-1处有1个明显的N-H 伸缩振动吸收峰,在2 800~3 000 cm-1处,有两个C-H 伸缩振动吸收峰,在1 626 cm-1处有1 个较大的酰胺I 带吸收峰,在1 516 cm-1处出现1个较大的酰胺Ⅱ带吸收峰[6],在1 234 cm-1处出现1 个较小的酰胺Ⅲ带吸收峰。此外,在1 430~1 445 cm-1具有CH2弯曲和CH3变形的特征吸收峰,在1 370~1 390 cm-1具有CH 弯曲与对称CH3变形而出现特征吸收峰[20]。从贝壳粉末(图2(c))中,也得到几丁质提取物,并鉴定到几丁质特征吸收峰(图2(d)):在3 269 cm-1处具有N-H 伸缩振动吸收峰,在2 800~3 000 cm-1处,也出现两个C-H伸缩振动吸收峰,在1 623 cm-1处出现一个酰胺I 带吸收峰,在1 513 cm-1处有1 个酰胺Ⅱ带吸收峰,并在1 560 cm-1处还出现一个较小的酰胺Ⅱ带吸收峰,在1 232 cm-1处出现1 个较小的酰胺Ⅲ带吸收峰。此外,在1 430~1 445 cm-1也具有CH2弯曲和CH3变形的特征吸收峰,在1 370~1 390 cm-1处也具有CH 弯曲与对称CH3变形而出现特征吸收峰。

图2 厚壳贻贝角质层和贝壳中几丁质的提取及鉴定
2.3 几丁质相关基因的筛查及表达分析
根据厚壳贻贝外套膜三褶皱的转录组数据,筛查出34 个几丁质相关基因,其中几丁质合成相关的基因有7 个(图3(a)),几丁质降解相关的基因有16 个(图3(b)),几丁质结合相关基因有11个(图3(c))。根据功能域预测,7 个几丁质合成酶基因(CHS)均具有chitn_synth_2 结构域。而筛查到的几丁质酶主要包括18家族和20家族糖苷水解酶两类,在16 个几丁质酶中,有10 个具有Glyco_18 结构域(18 家族糖苷水解酶),其余6 个具有Glyco_hydro_20 结构域(20 家族糖苷水解酶)。其中McCHIA1、McCHIA4、chitotriosidase-1-like 还具有2 或3 个几丁质结合结构域(ChtBD2);还 有4 个β 氨 基 己 糖 苷 酶(Bhexosaminidase)除了具有Glyco_hydro_20 结构域外,还具有一个特殊的碳水化合物结合域CHB_HEX结构域。厚壳贻贝11个几丁质结合相关的基因,均具有1到5个ChtBD2结构域(图3(c)),其中,Coadhesin 除了ChtBD2 结构域,还具备3 个血小板反应蛋白TSP1 结构域;Chitin binding/VWA1、Chitin binding/VWA2 还各自具备1 个与几丁质结合相关的VWA 结构域;mucin-2-like 还具有1 个层粘连蛋白LamG 结构域;McCBP1 还具备1个凝血因子5/8 C-末端FA58C结构域。
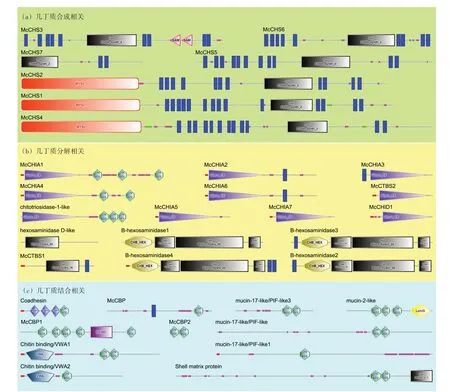
图3 厚壳贻贝几丁质相关基因的结构域预测
为进一步研究外套膜三褶皱与几丁质相关功能的关系,对筛查到的几丁质相关基因进行表达量分析(图4),并在三褶皱差异表达基因的GO富集结果中筛查几丁质相关的GO term(图5)。根据三褶皱中几丁质相关基因的表达量分析结果,发现几丁质合成相关基因主要在内、中褶皱高表达;而几丁质结合相关基因的表达模式则与之相反,主要在外褶皱高表达;筛查到的16 个几丁质酶基因,则主要呈现两种表达模式:一类是在外褶皱中高表达,一类在内、中褶皱中高表达。其中,在外褶皱中高表达的几丁质酶为壳三糖苷酶(McCHIA),属于18 家族糖苷水解酶;而在内、中高表达的几丁质酶多为二-N-乙酰壳二糖酶(McCTBS)、β 氨基己糖苷酶(B-hexosaminidase)等。此外,从外套膜三褶皱差异基因的GO富集分析结果中,筛查出多个几丁质相关的GO term(图5),发现内、中褶皱的差异高表达基因主要富集到几丁质生物合成相关的GO term,而外褶皱则主要富集到几丁质结合、代谢、角质层色素沉着相关的GO term。
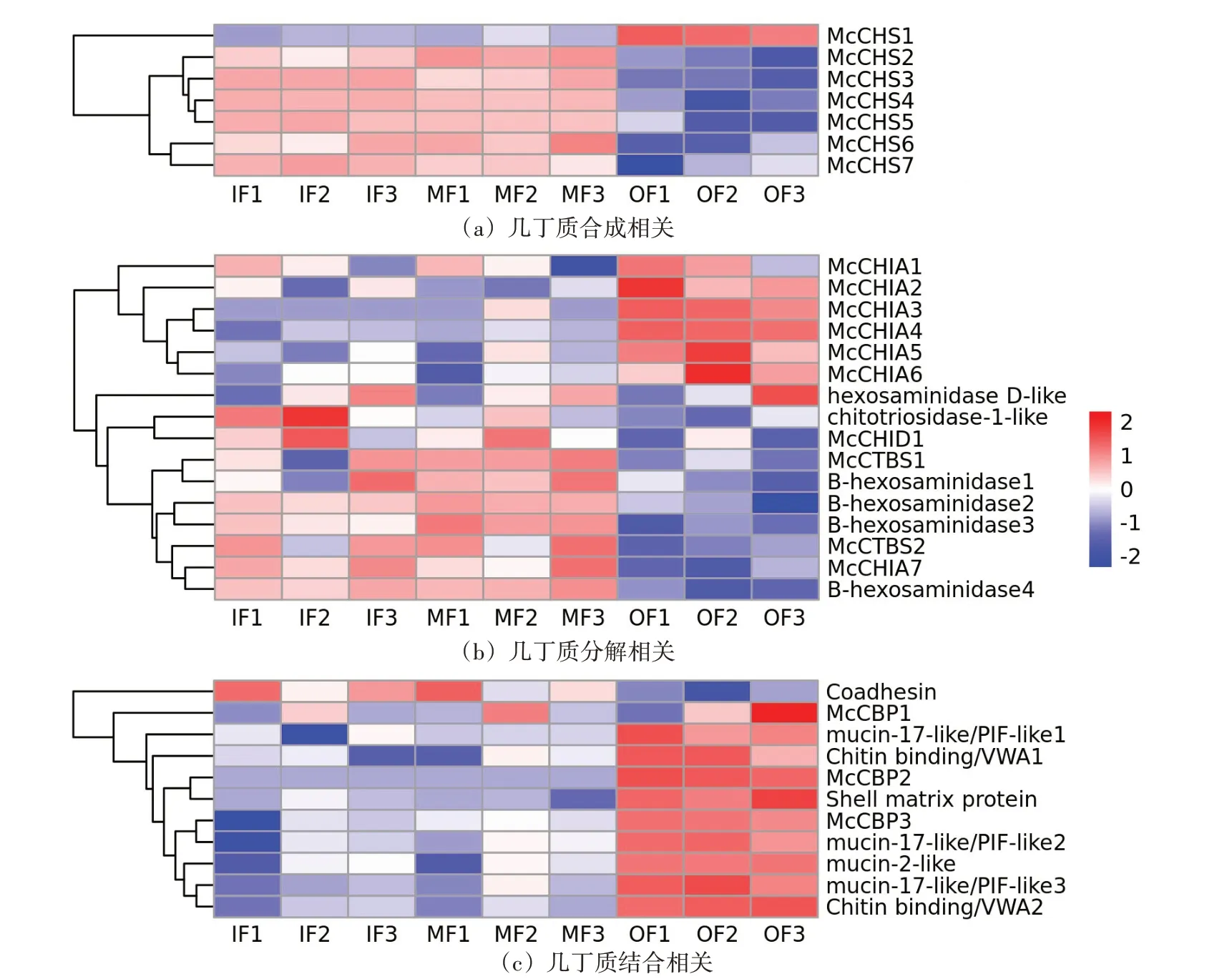
图4 厚壳贻贝几丁质相关基因在外套膜三褶皱中的表达分析
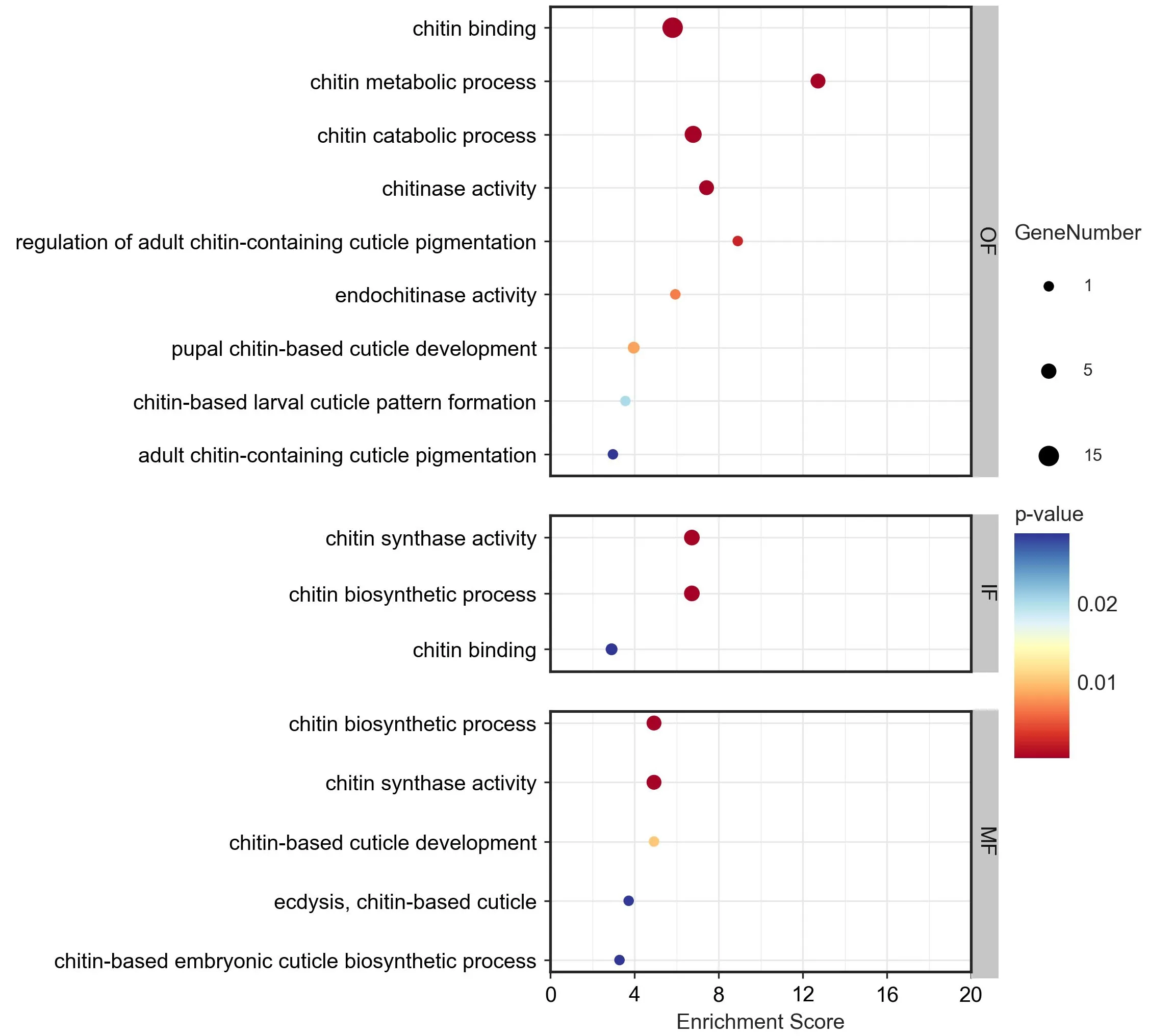
图5 厚壳贻贝外套膜三褶皱中高表达基因的GO富集分析
3 讨论
在自然界中,碳酸钙晶体会嵌入以几丁质为主成分的有机基框架中进行生物矿化,因此,本研究在提取几丁质时,进行了脱钙、脱蛋白等过程。根据FTIR 结果,本研究成功在厚壳贻贝的角质层和贝壳层提取到几丁质,二者分别在3 272 cm-1和3 269 cm-1处均存在单一吸收峰,这表明贻贝角质层和贝壳层中提取的几丁质均为β-几丁质而非α-几丁质[3-4,21]。但由于贝壳层与角质层中有机质和矿物质的构成存在差异,两种几丁质提取物的FTIR 吸收峰发生了一定程度的左右平移。此外,贝壳层中几丁质提取物在1 560 cm-1处还多了一个较小吸收峰(图2(d),蓝色箭头所示),二者在吸收峰的大小上,角质层提取物在1 513~ 1 626 cm-1处的吸收峰也大于贝壳层提取物,这表明,角质层中几丁质含量要更高于贝壳层中几丁质含量,且所含其他杂质也较少。几丁质作为贝壳有机基质的重要组成,在生物矿化中具有关键调控作用,在牡蛎、珠母贝、芋螺等贝类贝壳中均鉴定到几丁质的成分[5-7,21],且这些贝类贝壳中的几丁质提取物均是具有β-几丁质的特征吸收峰。
除几丁质外,贝壳基质蛋白也是贝类贝壳有机基质的重要组成,此前,在长牡蛎[22]、贻贝[19]、海螂[23]、栉江珧[24]、珠母贝[25]、栉孔扇贝[26]等双壳贝类的贝壳基质蛋白研究中,均鉴定到含有几丁质结合结构域的蛋白质以及几丁质代谢相关蛋白。在紫贻贝的研究中,也发现几丁质在幼虫外壳的形成和功能中发挥着重要作用[25],在翡翠贻贝贝壳蛋白质组分析中,还发现大量具有几丁质结合结构域的蛋白质[27]。因此,探究几丁质相关的壳基质蛋白有助于更好地了解厚壳贻贝贝壳有机骨架的形成。本研究利用外套膜边缘三褶皱的转录组数据,筛查出厚壳贻贝中几丁质合成、代谢、结合相关基因。在筛查到的三类几丁质相关基因中,几丁质酶数量最多。自然界中,几丁质酶主要包括几类糖苷水解酶(18、19、20、23、48 家族),而在厚壳贻贝中鉴定到了18 家族和20 家族两类糖苷水解酶。结合这三类几丁质相关基因在三褶皱中的表达量分析,发现这些几丁质酶在三个褶皱中的表达模式有所区别:β 氨基己糖苷酶、二-N-乙酰壳二糖酶等几丁质酶基因主要在内、中褶皱表达;而壳三糖苷酶类的几丁质酶主要在外褶皱表达。此外,根据表达模式,发现几丁质合成酶基因主要在内、中褶皱高表达,而几丁质结合蛋白相关基因的表达模式却与之相反,主要在外褶皱高表达。这表明外套膜三层褶皱在几丁质相关的功能上可能存在差异。
此前,在珠母贝[12]、三角帆蚌[14]、长牡蛎[21]等双壳贝类的研究发现几丁质结合蛋白能与几丁质和碳酸钙结合,并指导碳酸钙晶体的结晶并参与调控贝壳的形成。在长牡蛎、紫贻贝和大扇贝的贝壳损伤研究中,贝壳修复后外套膜差异表达的蛋白质中,均发现具有几丁质结合相关的VWA 结构域、LamG 结构域[15-16],这些研究表明具有几丁质结合相关的VWA 结构域、LamG 结构域蛋白能够参与贝壳损伤后的修复。结合在厚壳贻贝中的研究结果,几丁质结合蛋白主要在厚壳贻贝在外褶皱高表达,在这些外褶皱高表达蛋白中也发现几丁质结合相关的VWA 结构域、LamG 结构域的存在,由此推测,厚壳贻贝可能通过外褶皱调控几丁质框架的形成,参与贝壳的形成与修复。此外,长牡蛎壳基质蛋白的研究中也鉴定到几丁质酶和几丁质合成酶[28],在酸化海水处理的长牡蛎中,几丁质合成酶表达水平被显著抑制,从而抑制几丁质的生物合成来抑制牡蛎钙化壳的形成[29]。在厚壳贻贝中,几丁质合成酶主要在内、中褶皱中高表达,贻贝可能通过内、中褶皱来调控几丁质合成,从而影响贝壳的形成。在贝类几丁质酶的研究中,珠母贝的几丁质酶能参与贝壳的形成[30];在静水椎实螺中,也发现几丁质酶会影响壳的形成[31]。在厚壳贻贝中,不同类型的几丁质酶基因在不同褶皱高表达,三个褶皱可能都能通过几丁质的代谢来影响壳的形成,但具体的作用方式可能不同,还有待深入研究。最后,外套膜三褶皱GO功能富集分析的结果也表明内、中褶皱主要参与调控几丁质的生物合成过程,而外褶皱可能主要参与调控角质层壳膜的形成。此外,在外褶皱中还富集到“含几丁质角质层色素沉着”的GO term,结合组织切片观察结果,壳膜沟处的外褶皱上皮细胞中能观察到棕色色素,而此处的中褶皱上皮细胞却未观察到色素,推测壳膜的色素沉着可能主要由外褶皱来调控。
4 结论
根据贝壳和外套膜边缘的形态观察,本研究发现厚壳贻贝外套膜边缘具有1 个较大的内褶皱及较小的中、外褶皱。在三褶皱中,外褶皱最小,其牢固贴附于壳上,外褶皱和中褶皱之间产生角质层壳膜并延伸包裹在贝壳表面,根据组织切片观察,三个褶皱的上皮细胞形态也存在明显不同。此外,本研究还从厚壳贻贝的角质层和贝壳中提取并鉴定到几丁质成分,并根据三褶皱转录组数据鉴定到几丁质合成酶(CHS)、几丁质酶(CHIA)和几丁质结合蛋白(CBP)三类几丁质相关基因。根据功能域和表达量分析,发现几丁质合成酶基因、β氨基己糖苷酶、二-N-乙酰壳二糖酶等几丁质酶基因主要在内、中褶皱高表达,而与之相反的,几丁质结合蛋白基因和壳三糖苷酶类的几丁质酶基因主要在外褶皱中高表达。三褶皱间的GO富集分析也表明厚壳贻贝的内、中褶皱可能主要参与调控几丁质的生物合成,而外褶皱则主要参与调控几丁质结合、代谢和角质层色素沉着等过程。综上所述,厚壳贻贝外套膜三褶皱不仅在形态结构上存在显著差异,其在与几丁质合成、代谢、结合等功能方面也存在差异,其中,外褶皱可能在厚壳贻贝的几丁质骨架形成和色素沉着中发挥更重要的作用。本研究有助于了解厚壳贻贝贝壳有机骨架的几丁质成分和几丁质合成及代谢等相关基因,为厚壳贻贝贝壳形成和生物矿化的研究奠定了基础。
