噬菌体裂解酶及其在畜禽生产中的应用研究进展
2024-01-09谢自强宋瑞铭郝柳杭高庆羽张永英石玉祥
谢自强,宋瑞铭,郝柳杭,高庆羽,赵 越,李 悦,张永英,石玉祥
( 河北工程大学生命科学与食品工程学院,河北 邯郸 056000 )
细菌耐药性导致了严重的公共卫生安全问题,为响应农业农村部减抗、替抗、无抗的战略提案,急需开发出新的、绿色安全的替抗产品。噬菌体(bacteriophage,phage)是能够感染细菌、真菌、放线菌或螺旋体等微生物的病毒的总称[1],因其以细菌为寄主,也称为细菌病毒,根据噬菌体与宿主相互关系可分为裂解和溶原两种类型[2]。裂解性噬菌体具有杀菌效果,可直接进入宿主细菌进行复制和裂解[3-4],在感染细菌后期控制编码产生裂解酶,破坏细菌细胞壁肽聚糖层,最终使细菌破裂死亡。噬菌体遗传物质中可能携带耐药和毒力基因以及特异性杀菌能力制约了噬菌体的临床应用,且细菌菌株通过突变、受体修饰、被动适应、限制性修饰、规律间隔成簇短回文重复序列(clustered regularly interspaced short palindromic repeats associated systems,CRISPR/Cas)和假溶原等方式对噬菌体产生耐药性[5]。为克服这一影响,有研究尝试利用基因重组技术对裂解酶蛋白进行表达,结果显示了一定的抑菌效果,且没有关于细菌对噬菌体裂解酶产生耐药性的报告[6]。目前,针对噬菌体裂解酶的研究主要通过修饰改造其裂解谱,及提高抗菌活性,从而实现对不同种属耐药菌株的杀菌效果。此外,噬菌体裂解酶制剂在治疗细菌性疾病时具有特异性强、无残留、效果显著等优势,可用于畜禽细菌性疾病的治疗,解决耐药菌所致感染疾病,还可作为饲料添加剂用于改善畜禽肠道菌群、提升畜禽抗氧化能力。基于此,本文综述了裂解酶的结构、作用方式及应用概况等,以期为裂解酶制剂的开发提供参考。
1 噬菌体裂解酶概述
1.1 裂解酶的分类及结构
裂解酶(endolysin 或lysin)又称内溶素(lysozyme)、胞壁质酶(muramidase),是噬菌体编码的酶,在侵染细菌的后期产生,可降解细菌细胞壁内的特定键,从而释放子代病毒。一般根据裂解活性可分为不同类型:乙酰壁酰胺酶(acetylmuramidases)、转糖基酶(transglycosylases)、氨基葡萄糖苷酶(glucosaminidases)、酰胺酶(amidases)和内肽酶(endopeptidases),见图1。它们可以降解不同细菌的细胞壁,其作用的靶点是细菌的肽聚糖层[7-8]。
肽聚糖聚合物由交替的N- 乙酰壁酸(Nacetylmuramic acid,MurNAc)和N-乙酰氨基葡萄糖(Nacetyl-D-glucosamine,GlcNAc)残基组成,通过β-1, 4 糖苷键连接。相邻的聚糖链通过短的相互连接的肽交联,每个肽通过涉及肽的第一个氨基酸的酰胺键与MurNAc 残基连接(通常为L-丙氨酸)[9]。乙酰壁酰胺酶(如N-乙酰基β-D-壁酰胺酶)、氨基葡萄糖苷酶(如N-乙酰-β-D-氨基葡萄糖苷酶)、转糖基酶(如裂解转糖基酶)是糖苷酶的亚类,其通常会切割连接在肽聚糖层的MurNAc和GlcNAc交替聚合结构的β-1, 4 糖苷键。酰胺酶(如N-乙酰胞壁酰-L-丙氨酸酰胺酶)可催化MurNAc 和肽茎部分中的第一个氨基酸L-丙氨酸之间的酰胺键的断裂。内肽酶(如L-丙氨酰-D-谷氨酸内肽酶)可切割干肽的两个氨基酸之间的键,在肽桥内发生键的断裂[10]。
由于革兰氏阳性菌和革兰氏阴性菌细胞壁结构的差异,不同细菌噬菌体裂解酶在结构上存在区别。革兰氏阳性菌噬菌体裂解酶通常呈现模块化结构,由两个具有不同活性的结构域组成:一个N端酶活性结构域(EAD)和一个C端细胞壁结合结构域(CBD),由一个短而灵活的连接体连接,见图2。CBD负责底物识别,附着在细胞壁上的特定配体分子上,裂解酶的特异性通常由CBD决定。在细胞裂解后,一些CBD会与细胞壁碎片上的底物紧密结合,阻止它们过早裂解周围可能含有预装配噬菌体成分的细菌细胞。EAD负责催化特定PG键的断裂以破坏细胞壁。
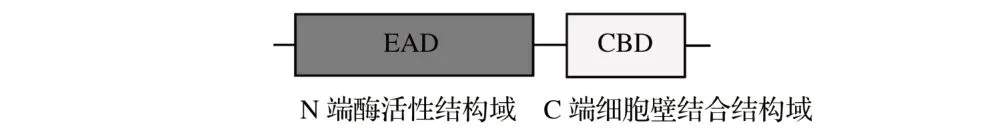
图2 针对革兰氏阳性细菌的模块化裂解酶的一般结构Fig.2 General structure of modular lyase for Gram-positive bacteria
由于外膜可有效保护革兰氏阴性细菌免受游离裂解酶的攻击,革兰氏阴性菌噬菌体裂解酶在宿主细胞裂解后不需要CBD 与细胞碎片结合,以最大限度地减少过早裂解。因此,革兰氏阴性菌噬菌体裂解酶通常具有球状结构,带有单个EAD以破坏细胞壁。一些革兰氏阴性菌噬菌体裂解酶被发现具有模块化结构(见图3),如由沙门菌噬菌体编码的PVP-SE1gp146[11]、SPN1S_0028[12]和Gp110[10]。与革兰氏阴性菌噬菌体裂解酶不同,模块化革兰氏阴性菌噬菌体裂解酶均具有C 端EAD 和N 端球形蛋白结合域(PBD),其识别革兰氏阴性细菌中常见的PG成分[13]。
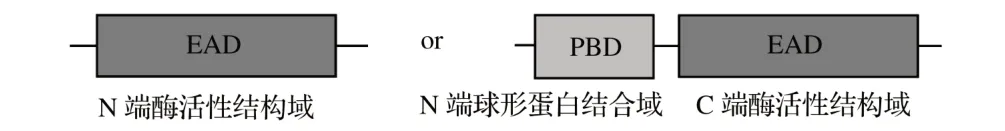
图3 针对革兰氏阴性细菌的球状和模块化裂解酶的结构安排Fig.3 Structural arrangement of spherical and modular lyases for Gram-negative bacteria
1.2 裂解酶作用机制
噬菌体介导的裂解有两种基本策略:一种策略是单链DNA(ssDNA)或单链RNA(ssRNA)噬菌体通过抑制细菌细胞壁肽聚糖的合成,依靠单基因编码的蛋白质分解细菌;而另一种策略是95%以上的已知双链DNA(dsDNA)噬菌体(如λ噬菌体)利用至少两种蛋白质,如绝大多数采用“穿孔素(holin)-裂解酶”裂解系统。大部分裂解酶没有信号肽,因此它们不会通过细胞质膜转位攻击肽聚糖中的底物,这种运动由裂解系统中的第2个噬菌体基因产物穿孔素控制。在穿孔素的作用下造成细菌细胞膜穿孔,使裂解酶顺利作用于肽聚糖靶点,导致宿主细胞溶解[13]。随着噬菌体在被感染细菌中发育,裂解酶在细胞质中积累,等待噬菌体成熟。在基因的调控下,穿孔素分子被插入细胞质膜形成斑块,最终导致细胞膜的广泛破坏[14]。裂解酶快速降解肽聚糖层,导致其渗透溶解,革兰氏阳性细菌容易受到裂解酶的影响,而革兰氏阴性细菌则具有天然的抵抗力。将外膜透过剂与噬菌体裂解酶共同作用于革兰氏阴性菌,可提高其抗菌活性,发挥裂解酶的作用[15]。且细菌能够被以重组蛋白形式进行外源表达的裂解酶杀死[16],这就可以通过基因工程手段克隆到表达载体进行大量制备,并配合外膜透过剂进行杀菌。因此,噬菌体裂解酶以一种普遍的物种特异性方式快速切割肽聚糖的独特能力,使其成为有前途的潜在抗菌剂。
2 噬菌体裂解酶在动物疾病临床诊疗中的应用
国内外已利用基因重组技术,先后制备出多种噬菌体重组裂解酶,包括大肠杆菌、金黄色葡萄球菌、沙门菌、布鲁菌、链球菌噬菌体裂解酶等,可用于治疗金黄色葡萄球菌、链球菌导致的奶牛乳腺炎;养殖环境的消杀;肉、蛋等畜禽产品的净化;此外,噬菌体重组裂解酶作为饲料添加剂应用于畜禽养殖还可提升畜禽产品质量等[17]。杨瑞思等[18]以3 日龄白羽肉鸡为试验对象饲喂复合噬菌体裂解酶,试验28 d后检测肠道菌群结构和肝脏抗氧化物酶的变化。结果显示,复合噬菌体裂解酶可提高肠道有益菌群数量和肝脏抗氧化能力。王子晶[19]建立了2型猪链球菌致小鼠菌血症模型,使用200 µg/只的噬菌体裂解酶Ply1228 进行治疗后,可显著降低各个组织器官及血液的载菌量,显著减轻炎症及病理变化,对小鼠的保护率高达100%。张楸杨[20]构建了高效表达噬菌体裂解酶LySP2 毕赤酵母菌株,通过诱导后,其体外抗菌效果良好,对鸡白痢沙门菌雏鸡感染模型的保护率高达70%,对肝脏及肠道沙门菌的清除率为100%和50%。Zhang等[21]研究发现,噬菌体裂解酶LysGH15具有对表皮葡萄球菌、溶血性葡萄球菌和人型葡萄球菌形成的葡萄球菌浮游细胞和生物被膜的清除能力。有研究通过构建表皮葡萄球菌菌血症小鼠模型进行体内试验,结果显示,腹腔注射20 μg/只的LysGH15 24 h后,小鼠血液中表皮葡萄球菌细胞计数比未处理的对照小鼠降低4个对数单位,器官中降低3个对数单位;同时血液中的细胞因子水平也有所降低,器官病理变化得到改善。综上,LysGH15表现出治疗由不同葡萄球菌引起的生物膜相关或非生物膜相关感染的潜力。
Ding 等[22]鉴定了一种新的内切酶LysSE24 并进行异源表达和纯化,发现其在0.1 µmol/L 浓度下对肠炎沙门氏菌ATCC 13076 具有最佳杀菌活性;在体外与外膜渗透剂联合培养10 min 后,沙门氏菌群显著减少。此外,内切酶LysSE24 还显示了对23 种经测试的多重耐药沙门菌菌株的抗菌活性。Xu 等[23]发现了1 种新型的噬菌体裂解酶PlyEc2,对关键的革兰氏阴性菌(包括大肠杆菌、沙门菌、志贺氏菌、不动杆菌和假单胞菌)具有强大的杀菌活性;在一定条件下,PlyEc2对产志贺毒素大肠杆菌O157:H7显示出较高的杀菌活性。此外,这种裂解酶还能够在体外将几种致病菌株的细菌滴度降低5个对数单位以上,从而实现完全灭菌。刘晓航[24]通过分子生物学技术将亚栖热菌和栖热菌噬菌体的两种裂解酶基因MMP phg和TSP phg进行拼接,后将融合片段克隆到表达质粒上,得到两种融合酶的表达系统,结果表明,融合酶对两种噬菌体宿主菌均具有明显的裂解作用。噬菌体对细菌具有高度特异性,只能够裂解其对应的菌株,而大多数表达纯化的噬菌体裂解酶的抗菌谱则更广,这表明更多的细菌种类具有某一裂解酶的细胞壁识别位点。
3 噬菌体裂解酶的改造及相关应用
噬菌体编码的裂解酶被认为是对抗革兰氏阳性菌的有效替代品,为了发挥裂解酶在对抗多重耐药细菌中的全部潜力,人们越来越关注能够使裂解酶克服靶向革兰氏阴性菌的外膜屏障[25]。由噬菌体裂解酶衍生的新型抗菌剂也不断出现。抗菌肽(antimicrobial peptides,AMPs)是由生物体产生的阳离子肽,通常由20~60个氨基酸残基组成。在许多结构-功能关系定量研究中,AMPs的正电荷数和疏水性是膜破裂的关键结构参数。虽然裂解酶在抑制革兰氏阴性菌中的应用有限,但AMPs 是噬菌体裂解酶未能突破革兰阴性菌外膜的潜在解决方案。Ning 等[26]设计的阳离子肽嵌合裂解酶Lysqdvp001-5aa、Lysqdvp001-10aa 和Lysqdvp001-15aa 是基于副溶血性弧菌噬菌体qdvp001 中的溶血素Lysqdvp001 设计的,通过将阳离子肽融合到Lysqdvp001 的C 末端,增强其对副溶血性弧菌的杀菌能力。这些嵌合裂解酶对副溶血性弧菌的抗菌活性以及内外膜通透性优于原噬菌体裂解酶,且其抗菌谱比噬菌体qdvp001 宽得多,Lysqdvp001-15aa 的MIC 和MBC 分别使牡蛎中副溶血性弧菌的数量减少了3、4 个对数单位。MBC 的Lysqdvp001-15aa 清除了约50%的副溶血性弧菌生物膜,并抑制了90%以上的细菌生物膜的形成。Nie等[27]通过生物信息学分析和设计,获得了4 种新型嵌合裂解酶(P361、P362、P371 和P372),它们是沙门菌噬菌体裂解酶和新型抗菌肽LeuA-P的融合。重组嵌合裂解酶在大肠杆菌BL21(DE3)菌株中表达,对沙门氏菌表现出高度特异性的抑制作用。Lu 等[28]将抗菌肽A 肽残基与噬菌体MK54的内溶素LysG24连接,构建1种新的肽修饰内溶素LysCA,研究LysG24 和LysCA 的抗菌活性。结果表明,LysG24 和LysCA 的裂解谱比噬菌体MK54 的裂解谱宽。体外试验表明,LysG24和LysCA在6 h内杀死了99%的宿主菌,LysCA 的溶解能力和环境适应性明显强于LysG24。体内试验表明,LysG24 和LysCA 在感染肺炎克雷伯菌(LPKP)的小鼠肺炎模型中均能够有效减少肺部炎症反应,治疗组肺炎克雷伯菌(LPKP)载菌量明显低于细菌组,其中LysCA治疗效果更明显。Wang等[29]从耐甲氧西林金黄色葡萄球菌(MRSA)噬菌体JD007中鉴定出1种细胞壁水解酶JDlys,将JDlys 与细胞穿透肽(CPPTat)融合,用于消灭细胞内耐甲氧西林金黄色葡萄球菌感染。结果发现,CPPTat-JDlys 融合后几乎不影响JDlys 的溶菌活性,能够有效消除细胞内耐药金黄色葡萄球菌,减轻MRSA引起的炎症反应和细胞损伤。抗菌肽-裂解酶嵌合体策略激发了噬菌体裂解酶在体外无法特异性抑制外膜致密的革兰氏阴性菌,开发通用嵌合裂解酶跨越革兰氏阴性菌的外膜屏障,克服针对裂解酶的免疫反应。
除了修饰改造外,裂解酶与抗生素联用也可提高抗菌活性、降低细菌耐药性[30]。Indiani 等[31]研究发现,链球菌裂解酶CF-301与达托霉素或万古霉素的组合具有协同作用,与人体血液中的两种特定成分(即溶菌酶和白蛋白)之间也存在协同作用,该结果证明了CF-301 作为静脉内给药抗微生物剂治疗金黄色葡萄球菌菌血症和心内膜炎的潜在有效性。纳米封装可通过保护蛋白质不被降解、提供持续释放,从而提高裂解酶的稳定性、保质期和治疗效果,从而帮助实现更好的治疗效果。Kaur等[32]研究发现,藻酸盐-壳聚糖纳米颗粒(Alg-Chi-NPs)可做作为裂解酶LysMR-5的载体,通过钙离子诱导藻酸盐核的预凝胶化及其与壳聚糖的络合,制备了LysMR-5负载纳米颗粒,包封后裂解酶结构完整生物活性不变,对金黄色葡萄球菌具有杀菌效果。
4 结论
与抗生素相比,噬菌体裂解酶治疗细菌感染具有以下明显优势:具有一定的特异性,避免了生物失衡和引发继发感染;不会使细菌产生耐药性;快速发挥杀菌作用;与针对耐药菌的新型抗生素相比,裂解酶制剂研发时间更短。虽然不同种类的裂解酶靶向特异性差异很大,但通过分子生物技术的改造可以发挥其抗菌潜力。
许多裂解酶在治疗抗生素耐药细菌方面显示出良好的效果,但其应用与后续研究同样面临挑战。外膜的存在导致裂解酶对革兰氏阴性菌的治疗活性较低;裂解酶体内半衰期短,使用时引发免疫反应,在体内易失去酶解活性;噬菌体治疗缺乏可靠的临床试验数据和相应的监管批准;给药方式、剂量不明确;裂解酶蛋白外源表达中存在诸多问题等。具有广谱裂解能力的酶筛选及改造、裂解酶辅助因子的探索、裂解酶规模化生产、裂解酶克服体内代谢的成药方式、成药后用药方式剂量等均是未来值得深入研究的方向。面对饲料端禁抗的大背景,噬菌体裂解酶势必会成为新一代对抗细菌性疾病的“利刃”,在对抗耐药细菌、清除生物被膜、作为饲料添加剂应用于畜禽养殖中发挥其潜力。
