植物异三聚体G蛋白研究进展
2024-01-07尚忠林康二芳李雨珂
尚忠林, 康二芳, 李雨珂, 付 玉
(河北师范大学 生命科学学院,河北 石家庄 050024)
作为一种古老且保守的信号转导系统,异三聚体G蛋白及其关联蛋白几乎存在于所有的真核生物中.异三聚体G蛋白一般由α,β和γ这3个结构不同的亚基组成,非刺激条件下,3个亚基结合形成的异三聚体蛋白定位在细胞质膜上.物理或化学因素可以作用于细胞质膜或细胞内的受体,活化的受体可以直接或间接作用于异三聚体G蛋白,Gα亚基结合GTP(鸟苷三磷酸)后活化并与Gβγ二聚体分离,Gα和Gβγ分别与细胞质膜或细胞质中的靶蛋白结合并触发信号级联反应,对于细胞质膜电位、跨膜离子流动、细胞骨架动态、细胞质关键酶的活性、转录因子活性进行调控,细胞代谢和基因表达模式的变化最终影响了细胞分裂、分化进程.随后,Gα中的GTP被水解为GDP(鸟苷二磷酸),无活性的Gα重新与Gβγ结合形成三聚体,结束信号传递过程[1].近年来,研究人员围绕植物体内异三聚体G蛋白信号系统的组分、功能以及异三聚体G蛋白活化及灭活机制开展了深入细致的研究,取得了令人欣喜的新成果.
1 植物异三聚体G蛋白的组成及结构特点
最初,研究人员参照动物细胞中异三聚体G蛋白的结构和基因特征,利用同源比对和分子克隆方法寻找典型的异三聚体G蛋白α,β和γ亚基,在模式植物拟南芥中只发现了1种α亚基、1种β亚基和2种γ亚基,这意味着拟南芥中最多只能组成2种类型的异三聚体G蛋白,研究人员一度对于异三聚体G蛋白能否参与多个生理过程的调控产生了怀疑[2].不过,近年来一个重要的进展是在多种植物中都发现了超大G蛋白(extra large G proteins,XLG),这些XLG可以与Gβ和Gγ结合形成三聚体,发挥类似Gα亚基的功能,每一个物种都含有多种XLG,XLG分子结构和编码基因具有明显的多样性[3];另一个重要的发现是植物细胞中存在多种类型的Gγ[4];这为植物异三聚体G蛋白形成多样化组合体、参与多种生理反应的调控奠定了结构基础.
拟南芥中的典型Gα(AtGPA1)由383个氨基酸组成,与动物细胞Gα相一致的特征是含有几个典型的保守结构域,如RAS样结构域、螺旋结构域,还含有N-豆蔻酰化位点和棕榈酰化位点(图1).在多种植物中都发现了与AtGPA1结构类似的蛋白,如大豆、豌豆及烟草都含有至少2个典型的Gα蛋白.Gα广泛存在于各种组织中,在细胞分裂旺盛的组织(如分生组织)中表达较丰富,在细胞内Gα可以定位于细胞质膜和细胞内膜上[5-7].
XLG与典型的Gα蛋白具有一定的相似性,都具有RAS结构域和螺旋结构域,XLG与典型Gα的显著区别是N端含有由300~500个氨基酸组成的延长区,这使得XLG分子量远超典型的Gα[7](图1).每一种植物中往往存在多种XLG,如拟南芥中有3种(XLG1~XLG3),玉米中有3种(ZmXLG1~ZmXLG3),水稻中也有3种(OsXLG1~OsXLG3),它们可参与形成多样化的异三聚体G蛋白,为响应多种刺激、启动多样化的细胞信号转导过程、精准调控生长发育和胁迫耐受反应奠定了基础[3,6-7].功能验证实验显示,拟南芥中XLG2/3,AGB1和AGG1/2参与介导由鞭毛蛋白引发的防御反应[8],玉米中的3个XLG参与调控株高和顶端分生组织的发育模式[9].在低等苔藓植物小立碗藓(Physcomitrellapatens)中没有发现典型的Gα蛋白,但是却发现了一些XLG,与典型的Gβ和Gγ组合形成的异三聚体参与小立碗藓生长发育调控,这一结果为理解异三聚体G蛋白结构进化提供了新的视角和线索[10-11].
植物细胞中的Gβ类型较少,在多个物种(包括拟南芥、玉米、水稻、燕麦、烟草等)中都只检测到了一种Gβ,不同物种中编码Gβ的基因序列具有极高的相似度(图1).拟南芥AGB1,玉米ZGB1及水稻RGB1存在于多个器官组织中,这些Gβ主要分布在细胞质膜和细胞内膜上,与Gα的定位高度重合[2,6].
植物细胞中的Gγ按照结构特征分成3个类型:Ⅰ型、Ⅱ型和Ⅲ型(图1).Ⅰ型一般含有100~120个氨基酸,分子中间具有卷曲螺旋结构域,含有与Gβ结合必需的、保守的DPLL基序,C端可以被异戊二烯化以确保Ⅰ型的Gγ定位在细胞质膜上.拟南芥中含有2个Ⅰ型Gγ(AGG1和AGG2),N末端可与Gβ相互作用,C末端的CaaX结构域可被异戊二烯或其衍生物修饰.Ⅱ型与Ⅰ型Gγ的主要区别在于C-末端没有异戊二烯基化位点,但这并不影响蛋白的定位.Ⅲ型Gγ的N端区域与Ⅰ型和Ⅱ型相似,但是C末端序列中富含半胱氨酸,可能参与结合XLG;Ⅲ型Gγ成员包括拟南芥中的AGG3,水稻中的DEP1,GS3和GCA2,大豆中的Gγ8~Gγ10等[2,4,6].
2 异三聚体G蛋白信号转导系统组分
2.1 G蛋白偶联受体
动物细胞质膜上存在多种异三聚体G蛋白偶联受体(G protein-coupled receptor,GPCR),这些GPCR一般含有7个跨膜结构域,结合配体分子后,活化后的GPCR作用于异三聚体G蛋白,引起Gα与Gβγ的分离[1].最初人们循着同样的思路,试图寻找植物细胞中的GPCR.虽然基因相似性分析结果显示拟南芥基因组中可能存在超过20个GPCR基因,但只有一种类似于动物细胞GPCR的蛋白GCR1被检测出来,GCR1能够与GPA1结合、互作,响应刺激后激活异三聚体G蛋白,在其他一些物种中也陆续发现了与GCR1类似的蛋白[12].
近几年研究人员在水稻、玉米和甘蔗中发现了一些具有9个跨膜结构域的蛋白,经检验这些蛋白具有典型的GPCR活性.这些蛋白可以定位于细胞质膜或内质网膜中,与传统的GPCR需要配体结合才能活化的这一特征不同,这些蛋白在物理胁迫(如低温)下就可以发生结构变化并活化,随后向下游激活异三聚体G蛋白组分,启动胁迫耐受性反应.水稻和玉米中的COLD1参与诱导植株耐低温能力[13-15],甘蔗中的ShGPCR1参与诱导植株对干旱、盐渍和冷胁迫的耐受能力[16].
2.2 G蛋白信号调节因子(regulator of G-protein signaling,RGS)
RGS是所有生物体中都存在的一类GTP酶激活蛋白(GTPaes activating protein,GAP),能够促进异三聚体Gα水解GTP,并抑制Gα与GTP的结合,对Gα发挥抑制作用.拟南芥的AtRGS1是第一个在植物中发现的RGS蛋白,能与Gα蛋白直接结合互作,并且优先与结合了GTP、处于活化态的Gα结合.植物体内的RGS蛋白C末端一般都有保守的RGS盒结构域,与该区域连接的N末端区域能够形成7次跨膜的结构域.由于七跨膜结构域是动物细胞质膜GPCR的重要特征,最初研究者们猜测RGS1可以作为一种GPCR发挥作用,但是2方面的证据否定了这种推测:1) RGS1本身具有GTP酶活性,将结合在Gα中的GTP分解为GDP,使Gα蛋白处于非活化状态,RGS1实际上是一种抑制蛋白而不是G蛋白的活化因素; 2) 目前能够与AtRGS1特异性结合的配体分子还没有得到确认[6-8,10].最近的研究结果显示,RGS1与异三聚体G蛋白、类受体激酶结合形成复杂的受体复合物,与配体结合、被活化的受体激酶将RGS1磷酸化,解除了 RGS1对异三聚体G蛋白的抑制作用,随后异三聚体G蛋白可以自动活化、解体并激活下游信号组分,这为解析RGS1以及其他RGS家族蛋白发挥功能的作用机制提供了新的视角和切入点[17-18].
2.3 磷脂酶
哺乳动物细胞质膜上的磷脂酶Cβ(phospholipase Cβ,PLCβ)能够与异三聚体Gα结合,加强Gα的GTPase活性,其功能类似于RGS蛋白.植物中尚未发现类似的PLCβ蛋白,但是已经发现拟南芥中的磷脂酶Dα1(phospholipase Dα1,PLDα1)可以结合并调节Gα蛋白活性[19-20].植物体内的磷脂酶家族成员众多,某些成员像PLDα1一样参与G蛋白信号转导的可能性是存在的,这也为今后探究异三聚体G蛋白信号级联机制提供了新的思路.
2.4 类受体激酶
类受体激酶(receptor-like kinase,RLK)和类受体蛋白(receptor-like protein,RLP)是细胞质膜上重要的信号转导组分,一个物种中往往含有几百甚至上千种RLK以及RLP(如拟南芥含有超过600种RLK、水稻含有超过1 100种RLK),这为细胞感受复杂环境刺激、精准调控细胞代谢奠定了坚实的基础.虽然大多数受体激酶(或受体蛋白)的功能和信号转导机制尚不清楚,但是仅就已经阐明的一些受体激酶而言,已经有充分的证据显示异三聚体G蛋白参与由RLK和RLP启动的信号转导过程[6,8,16].例如,鞭毛蛋白受体FLS2与异三聚体G蛋白组成受体复合物,病原菌鞭毛蛋白与其受体结合,可以迅速活化异三聚体G蛋白、诱发防御反应[8,16];FERONIA(FER)可以通过异三聚体G蛋白β亚基(AGB1)调节气孔开关以及对盐渍的响应[21];类受体激酶ZAR1(zygotic arrest1)通过AGB1调控受精卵分裂、分化形成胚胎的过程[22].随着累积的证据越来越多,一些学者大胆猜测,RLK大家族中的很多成员可以发挥GPCR的作用,启动异三聚体G蛋白信号转导[6,10,22].
3 植物异三聚体G蛋白的活化和灭活机制
在哺乳动物细胞中,细胞质膜上的GPCR感受信号分子,通过其鸟苷酸交换因子(guanine nucleotide exchange factor,GEF)活性使Gα蛋白结合的GDP脱离并结合GTP,异三聚体随即解离为活化的Gα和Gβγ(图2).植物细胞Gα的生化特性与动物细胞明显不同:首先,以拟南芥GPA1为代表的植物Gα能够高效率结合GTP,具备较强的自激活能力,不要求必需由GPCR对其进行激活;其次,某些植物特有的蛋白质发挥GPCR的作用,例如RLK和RLP家族成员,它们可以通过磷酸化RGS、解除其对G蛋白的抑制作用,也可能直接对异三聚体G蛋白进行磷酸化调控将某些亚基激活[6-7,16,21].
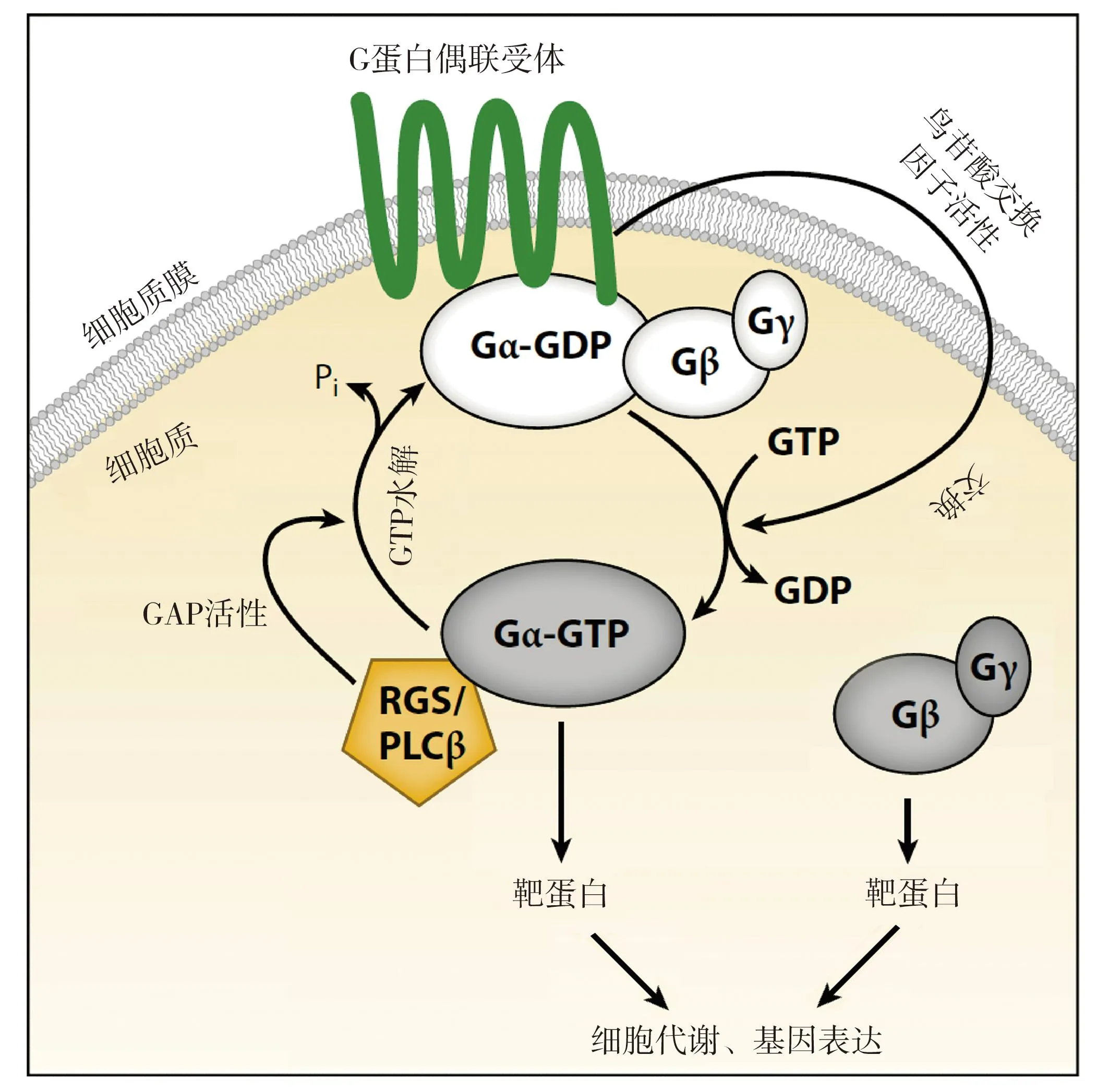
图2 植物异三聚体G蛋白活化和灭活机制(Pandy [6])Fig.2 The Activation and Inactivation of Heterotrimeric G Proteins in Plants
动植物细胞中异三聚体G蛋白灭活机制都是通过Gα中的GTP水解为GDP实现的.植物细胞中Gα的GTPase活性比动物细胞中的Gα低很多,难以保证GTP及时水解和Gα的及时灭活.而存在于植物细胞中的多种能结合异三聚体Gα并增强其GTP酶活性的GAP蛋白很可能在G蛋白失活过程发挥重要作用[5-7].RGS是GAP蛋白家族的重要成员,在绿藻、苔藓植物、裸子植物、被子植物中都发现了RGS,已经发现拟南芥中的RGS1可显著提高Gα的GTPase活性[6],拟南芥或大豆的Gα缺失突变体与RGS1缺失突变体表型相反,也证实了RGS作为G蛋白信号失活剂发挥作用[23].一些磷脂酶(如拟南芥PLDα1)也可以作为GAP来加强GPA1的GTPase活性[18].进一步研究发现,拟南芥中的RGS1和PLDα1存在互作,RGS1与PLDα1的结合抑制了它的磷脂酶活性;而磷脂酸(PLDα1催化产物)可以结合、抑制RGS1的GAP活性,2种GAP相互制衡的关系为异三聚体G蛋白信号强度调控提供了一种更为复杂和精准的方式[24].
4 异三聚体G蛋白参与调控的生理过程
异三聚体G蛋白活化后,可以作用于细胞质膜上的靶蛋白,如离子通道、磷脂酶C诱导产生第二信使;也可以作用于细胞质、细胞核中的靶蛋白,参与调控细胞骨架稳态、酶活性、功能基因表达.异三聚体G蛋白参与调控的细胞代谢、细胞分裂和分化模式的变化可以影响植物营养生长和生殖生长,影响植物的株型、种子数量、种子大小等性状.异三聚体G蛋白还广泛参与动员植物应对病原微生物(细菌、真菌、病毒等)引起的生物胁迫和环境因素(光、干旱、盐等)剧烈变化引起的非生物胁迫.在引起上述生理效应的过程中,异三聚体G蛋白的3个亚基单独或协同作用于下游靶蛋白,对异三聚体G蛋白生理功能的研究也是基于观察这些亚基基因缺失突变体或超表达株系的表型指标变化.由于遗传操作技术不断进步,对于异三聚体G蛋白功能的研究也成为进展最为迅速的领域.
4.1 参与营养器官生长发育
异三聚体G蛋白参与调控根的生长发育.例如,玉米中异三聚体Gα(CT2)可以被多肽信使CLV3的受体CLV1,CLV2结合并活化,参与调控顶端分生组织的细胞分裂和分化,CT2缺失突变引起玉米生长受阻、植株矮化[25].拟南芥Gβ或Gγ缺失突变体主根生长速度加快、侧根数量显著增多,Gβγ通过调节根尖生长素运输负调控主根生长和侧根发生[26].拟南芥Gα,Gβ缺失突变体比野生型产生更多、更长的侧根,当葡萄糖作为一种信号分子引起RGS1活化,RGS1抑制Gα活性也使得生长素促进侧根形成和伸长生长的效应得到了显著加强[27].在细胞外ATP抑制拟南芥主根生长、引起根尖弯曲的过程中,Gα通过激活Ca2+通道、调控功能基因表达发挥积极的作用[28].
异三聚体G蛋白参与调控下胚轴生长.研究发现,AGB1通过抑制转录因子BBX21活性促进下胚轴伸长生长,AGB1的基因缺失突变体下胚轴生长明显减缓[29].黄瓜中的GPA1活化促进了细胞分裂和伸长,使种子萌发、根和下胚轴生长速度都加快[30].拟南芥Gβ参与下胚轴的向光性反应,在Gβ缺失突变体中由蓝光诱导的下胚轴向光弯曲反应显著减弱[31].水稻中超表达Gγ(RGG1)的基因后,细胞中细胞分裂素含量下降,细胞对细胞分裂素的敏感性下降,细胞无法正常分裂导致生长受阻、植株矮化[32].
异三聚体G蛋白参与叶片发育.在水稻中,Gα基因缺失突变带来几方面的变化:1) 叶肉细胞细胞壁变薄、对CO2的通透性增强; 2) 叶肉细胞间联系变得松散,叶片内部表面积增大,有利于CO2吸收; 3) 叶绿体膜对CO2的通透性增强.这些变化有利于在气孔开度较小的条件下吸收足够的CO2用于光合作用,促使水分利用效率提高,对干旱的耐受能力增强.这些结果显示在这些发育指标的调控过程中,Gα发挥负调控作用[33].在水稻Gα(RGA1)的基因缺失突变体中,叶片避光性反应增强、光能利用效率提高,参与捕光色素复合体组建的蛋白合成量增加,植物对于强光的耐受性增强,显示RGA1在水稻的光保护反应过程中可能发挥负调控作用[34].
4.2 参与气孔发育和气孔运动的调控
异三聚体G蛋白参与调控气孔器发育.拟南芥Gα的基因缺失引起叶片表面气孔频度下降,超表达Gα则使气孔频度显著增加;Gβ对气孔发育的影响与之截然相反,Gβ缺失突变体气孔频度显著升高,而其超表达株系气孔频度显著降低[35].不过,另一个实验发现,与野生型相比,Gα缺失突变体气孔开度更大,二氧化碳吸收能力更强,蒸腾效率更高,其结果是虽然突变体叶片上气孔器数量少,但是其二氧化碳吸收量与野生型基本相当[36].
多种调控气孔运动的信号通过异三聚体G蛋白信号组分调控气孔运动.例如,植物激素脱落酸(abscisic acid,ABA)引起气孔关闭的过程中,ABA受体可以活化Gα、诱导活性氧形成,并通过活性氧敏感的Ca2+通道促进Ca2+内流,最终引起气孔关闭[37];ABA 还可以引起Gβ(AGB1)活化,Gβ作用于磷脂酶C(phospholipase C,PLC)催化产生第二信使IP3,诱导产生并维持Ca2+震荡,引起气孔关闭[38].在细胞外ATP诱导气孔开放的过程中,Gα通过活化质子泵、诱导Ca2+内流发挥积极作用[39].紫外线(UV-B)、乙烯可以激活G蛋白偶联受体GCR1,随后活化的Gα促进保卫细胞中活性氧和一氧化氮积累,最终引起气孔关闭[40].多肽信使RALF1作用于其受体FERONIA后引起气孔关闭,Gβ,Gγ和一些XLG参与了这一过程,但是经典的Gα不参与此过程[21].在细胞外钙调素促进气孔关闭的过程中,Gα通过诱导NO和H2O2积累发挥作用[41].高温和干旱诱导保卫细胞中NO积累、促进气孔关闭的过程中,Gβ发挥积极作用,豌豆Gβ在烟草中的异源表达增强了转基因烟草的高温和干旱胁迫耐受力[42].最新研究发现氢可以作为信号分子调控气孔运动,氢刺激拟南芥保卫细胞产生ROS和NO及促进气孔关闭的过程需要GPA1参与;在GPA1基因缺失突变体中氢刺激不能引起保卫细胞中ROS和NO积累,氢也不能引起气孔关闭[43].
4.3 参与生殖生长和胚胎发育
植物有性生殖过程影响果实、种子的数量和质量,直接关系到农业生产的经济效益.近年来研究发现,异三聚体G蛋白影响多种植物的花粉萌发、果实及种子发育.例如,水稻的Gα基因突变后,与分蘖、灌浆有关的多个基因表达量显著下降,影响水稻植株生长、抽穗和籽粒的生长并对产量造成不利影响[44].玉米的Gα及XLG表达量增加、活性增强可以使小穗密度、籽粒行数显著增加,叶形也有所改变,直立生长的叶片增多,这些农艺性状的变化都有利于玉米生产[9].水稻的XLG的基因缺失后,植株生长、花序发育、籽粒灌浆都出现障碍[45].水稻中的RGA1超表达后,花粉管对于弱光的适应性显著增强,在弱光下的糖吸收、利用能力显著增强,弱光对花粉管生长的抑制作用减轻,有利于完成双受精[46].在细胞外ATP促进拟南芥花粉萌发和花粉管生长的过程中,Gα通过维持钙通道活性发挥促进作用[47].水稻中RGB1调控胚乳细胞中的生长素合成酶活性、促进生长素合成,促进籽粒发育和淀粉积累,对产量形成具有重要作用[48].番茄中的一种Ⅱ型Gγ的基因缺失后,植株对生长素和ABA的敏感性发生变化,种子变小、果形变尖[49].亚麻荠中的Gβ参与种子发育和脂肪积累,Gβ缺失突变体的种子由细长形变成了短粗形,种子中脂肪含量也明显增加[50].
受精卵分化形成胚是种子发育过程的重要环节,拟南芥受精卵分化过程中,类受体激酶ZAR1可以作用于Gβ以调控受精卵不均等分裂,ZAR1和Gβ的基因缺失造成受精卵分裂形成的基细胞变短、顶细胞发育模式畸变[21];进一步研究发现,活化的Gβ可以作为架构蛋白将MAPK级联系统成员联系起来,使这些组分形成特异性的信号级联单元,对受精卵分化进行调控[51].
4.4 参与生物胁迫的响应
病原微生物(细菌、真菌、病毒等)侵害对于植物生长发育产生严重的负面影响,植物细胞可以感受病原微生物携带或释放的化学信息分子,并调动细胞内的免疫反应.已经有多项证据显示异三聚体G蛋白信号组分参与植物细胞免疫反应的动员过程.烟草中的异三聚体G蛋白基因缺失后,由超敏蛋白(harpin)引起的超敏反应、由激发子引发的气孔关闭过程都显著减弱,响应激发子刺激而表达的防御基因无法正常启动表达,显示异三聚体G蛋白在烟草防御反应过程中发挥关键作用[52].植物细胞对于细菌的鞭毛蛋白能够做出灵敏的响应,鞭毛蛋白受体FLS2及其共受体BAK1可以与异三聚体G蛋白及其调控蛋白RGS1组成复杂的受体复合物,FLS2与鞭毛蛋白flg22结合后引起BAK1活化、RGS1磷酸化,随后异三聚体G蛋白被释放并解离,活化的Gα可以直接作用于细胞质膜上的NADPH氧化酶,刺激产生活性氧爆发以诱导细胞的免疫反应;在此过程中,Gα,XLG2,AGB1,AGG1和AGG2都参与诱导免疫反应[8,53-55].拟南芥中的XLG1,XLG2,XLG3一同参与病原生物释放的模式分子触发的免疫反应,在XLG1/2/3三重突变体中,由模式分子引发的MAPK级联系统活化和抗病反应显著弱化[56].也有研究发现,flg22刺激造成的生长抑制过程中,Gβ发挥主要作用,AGB1缺失突变体对flg22刺激的抵抗力显著高于野生型[57].在拟南芥应对丁香假单胞菌侵害的过程中,细胞内的XLG2与Gβ一同发挥作用,动员对丁香假单胞菌的抵抗力[58].
4.5 参与非生物胁迫适应
在植物响应低温、高温、干旱和盐胁迫的过程中,多种异三聚体G蛋白信号组分参与诱导胁迫耐受性反应.如前所述,水稻、玉米中的GPCR(COLD1)和甘蔗中的ShGPCR1基因在低温、干旱、盐渍和冷胁迫下表达量增加,促进细胞中抗逆基因的表达,提高植株的胁迫耐受能力[13-15].番茄中Gα基因缺失造成冷胁迫耐受能力下降,由冷胁迫引起的活性氧积累增加,细胞受害严重;而超表达Gα的番茄细胞内活性氧清除系统活力显著增强,对于低温的耐受能力显著增强[59].黄瓜中的一种Gγ(CsGG3.2)可以使超氧化物歧化酶、过氧化氢酶、谷胱甘肽还原酶等活性氧清除酶活性增强,减少低温下活性氧积累和膜脂过氧化,增强黄瓜对低温的耐受能力[60].异三聚体G蛋白也参与响应高温胁迫,水稻Gα(RGA1)缺失突变体对高温的耐受能力显著增强,膜脂氧化减少,与糖代谢有关的酶活性显著增强,体内高能物质(如ATP)水平显著增加,显示在水稻耐受高温的过程中,RGA1可能发挥负调控作用[61].异三聚体G蛋白参与植物对干旱的响应,水稻中的RGA1的基因突变之后,植株对于干旱的适应能力显著加强,叶片水势提高、叶肉细胞中脯氨酸含量增加、叶片厚度增加,细胞内活性氧清除酶表达量增加、活性增强,显示在干旱适应过程中,异三聚体G蛋白发挥负调控作用[62].小麦Gβ的基因缺失突变体干旱胁迫耐受能力下降,Gβ超表达株系耐旱能力增强.干旱和盐胁迫下Gβ超表达株系小麦细胞中SOD活性更强,膜脂过氧化产物MDA更少,细胞中脯氨酸积累增加,增强了细胞对逆境的耐受能力[63].
水稻、玉米中的Gα缺失突变之后,植株对盐胁迫的敏感性降低,由盐胁迫引起的生长减缓、细胞衰老、叶绿素降解、细胞质渗漏等都得到了缓解,而且在突变体中盐胁迫对细胞分裂的抑制作用也显著减轻.这些结果显示Gα参与提高细胞对盐胁迫的敏感性,而降低Gα活性有助于提高植物对盐胁迫的适应能力[64].RALF1与其受体FER结合后,可以通过AGB1诱导盐胁迫耐受反应,在FER和AGB1缺失突变体中,由盐胁迫引发的ROS积累显著减弱,突变体的盐胁迫适应能力也显著减弱[65].
最新的一项工作中发现,一种非典型性的Gγ蛋白(AT1)参与作物对碱的感受和响应,敲除该基因后谷子、高粱、玉米和水稻对于碱的耐受能力显著增强.这种Gγ可以作用于水通道蛋白,抑制其磷酸化和对过氧化氢的外排,敲除该基因后水通道蛋白磷酸化加强、活性增强,能够更有效地将过氧化氢排出细胞,减少其对细胞的损害[66].这一发现为培育耐盐碱的作物提供了重要的切入点.
4.6 参与根瘤形成
豆科植物根瘤的形成对于其氮素供应和生长发育至关重要.研究发现,大豆中的异三聚体G蛋白对于根瘤形成至关重要,异三聚体G蛋白各亚基基因表达水平与根瘤形成数量呈正相关[67].进一步研究发现根瘤菌释放的结瘤因子与大豆细胞中的受体NFR1结合后,可以引起RGS蛋白磷酸化,活化的RGS发挥GTP酶活性,抑制异三聚体G蛋白活性,使得根瘤得以顺利形成[68].共生受体激酶(symbiosis receptor-like kinase,SymRK)可以和NFR1结合形成复合体,感受结瘤因子之后,SymRK将Gα磷酸化,使其无法与Gβγ二聚体结合,Gβγ可以独立发挥作用、促进根瘤形成[69].苜蓿和豌豆中的Gβ表达受抑制之后,根瘤形成数量显著减少,Gβ可能与磷脂酶C互作并调控细胞内第二信使的产生,参与调控根瘤形成[70].
5 展 望
近几年来,植物异三聚体G蛋白信号系统组分生理功能及作用机制的研究取得了令人振奋的进展,特别是研究材料从模式植物拟南芥转变为与人类生产生活密切相关的农作物(如水稻、小麦、大豆、黄瓜、玉米等)后,揭示了异三聚体G蛋白在控制作物生长发育、产量形成、品质改善、胁迫耐受性等方面发挥的作用,为未来采用遗传学手段改良农作物奠定了实验基础.今后一段时间,将现有成果整合起来,通过遗传操作手段改变异三聚体G蛋白信号转导组分表达水平、活性状态、组合方式以及对靶蛋白的作用强度,并全程跟踪遗传背景的变化对农作物生长发育、胁迫抗性的综合影响,将有望在创制生长发育良好、农艺性状出色、环境适应性好的作物品种方面取得新的突破.
