高寒草甸圆穗蓼叶解剖结构与草酸钙含量对生境干旱化的响应
2024-01-01阳佳英蔡惠李忠馗张大才






摘要:为揭示高寒草甸生境干旱化过程中侵入种圆穗蓼(Bistorta macrophylla)的生境适应能力,在滇西北白马雪山高寒草甸选择湿润、半湿润和干旱生境设置样方、采集实验材料。利用单因素方差分析、主成分分析等方法,分析叶片解剖结构和草酸钙含量与土壤水分之间的关系。结果表明:叶片栅栏组织紧密排列为2层,主脉中5~9个维管束呈环形排列,且木质部朝中心方向分布。叶片、上表皮及角质层、下表皮、栅栏组织与海绵组织、主脉与维管束、木质部与韧皮部等结构的厚度以及叶草酸钙含量在生境间差异显著(P<0.01),且干旱生境中数值最大,其中叶片草酸钙含量对植株耐旱能力贡献最大。下表皮角质层厚度、栅海比在生境间的差异不显著。圆穗蓼通过增加保水结构(表皮与角质层)厚度以减少水分蒸发,增加输水结构(维管束、木质部)厚度提高输水效率,增加草酸钙含量增强贮水能力。即采取增强保水和输水结构功能、提高解剖结构与草酸钙含量协同响应等策略抵御生境干旱化。
关键词:解剖结构;保水结构;输水结构;土壤水分梯度;草酸钙含量
中图分类号:Q944.56;S545""" 文献标识码:A"""" 文章编号:1007-0435(2024)06-1944-09
Response of Leaf Anatomy and Calcium Oxalate Content of Bistorta
macrophylla to Habitat Drought in Alpine Meadow
YANG Jia-ying, CAI Hui-wen, LI Zhong-kui, ZHANG Da-cai*
(Key Laboratory of National Forestry and Grassland Administration on Biodiversity Conservation in Southwest China,
Forestry College, Southwest Forestry University, Kunming, Yunnan Province 650224, China)
Abstract:In order to reveal the habitat adaptive capacity of the invasive species Bistorta macrophylla in the process of drought in alpine meadows,we selected humid,subhumid and drought habitats in the alpine meadows of the Baima Snow Mountain in northwestern Yunnan Province to set up sampling plots and collect experimental materials. Changes in leaf anatomy and calcium oxalate content along habitat drought were investigated using one-way ANOVA and principal component analysis. The results showed that the palisade tissues were tightly arranged in 2 layers,5~9 vascular cylinder in the main veins were arranged in a ring,and the xylem was distributed towards the center of the ring. Leaf thickness,upper epidermal and stratum corneum thickness,lower epidermal thickness,palisade(P) and spongy(S) tissue thickness,main vein and vascular cylinder thickness,xylem and phloem thickness,and leaf calcium oxalate content differed significantly among habitats and had the greatest values in the drought habitats (P lt;0.01),with the leaf calcium oxalate content contributing the most to the drought tolerance of B. macrophylla. Lower epidermal stratum corneum thickness and P/S did not differ significantly among habitats. Under drought stress,B. macrophylla reduced water evaporation by increasing the thickness of water retaining structures (epidermis,epidermal stratum corneum),improved water transport efficiency by increasing the thickness of water transporting structures (vascular cylinder,xylem),and enhanced the water storage capacity by increasing the calcium oxalate content. Namely,the strategies of enhancing the function of water retaining and water transporting structures and increasing the coordinated response of anatomical structures and calcium oxalate content were used to resist the drought of the habitat.
Key words:Anatomical structure;Water retention structure;Water transport structure;Soil moisture gradient;Calcium oxalate content
叶片是植物与生境接触面积最大的器官,对生境变化敏感。叶片解剖结构特征与植物生境适应能力密切相关[1-3],尤其是草本植物,叶片解剖结构随生境干旱化而显著变化[4-5]。近年来,在全球气候变化和人为干扰的双重影响下[6],高寒草甸生境干旱程度加剧,退化演替严重,群落原建群种的生存空间逐渐被压缩,且原生植物群落中的一些偶见种重要值逐渐增加成为侵入种,甚至在部分生境成为建群种[7]。在此过程中,侵入种的叶片解剖结构如何响应生境干旱化的机制仍不明晰。
植物受到干旱胁迫时,叶解剖结构全面响应,其功能也会相应的增强或减弱[8-9]。与此同时,保水结构(表皮、角质层)会增加厚度以防止水分蒸发,增强叶片的保水功能[10],是植物面对逆境时维持生长的关键;叶肉结构分为栅栏组织和海绵组织,是光合作用的场所,也是气体交换的地方,其厚度关系着植物光合效能[11]。生境干旱时,栅栏组织越厚,越有利于光合作用的提高,同时栅栏组织与海绵组织的比例关系着植物对生境变化的适应性[12],例如密穗马先蒿(Pedicularis densispica)在高海拔干旱生境中叶片不仅增加栅栏组织厚度,且更大程度增加海绵组织厚度,降低栅海比以增强对强光生境的适应[13]。输水结构(维管束、木质部)厚度越厚则植物运输效率越高,抗旱能力也越强[14-15]。可见,叶解剖结构的改变在逆境胁迫中作用巨大,能直接影响植物的生境适应能力。
草酸钙是由植株内源合成的草酸与外源吸收的钙离子共同反应而形成的生物矿物,大部分聚集在植物体内的晶异细胞中[16]。草酸钙是植物长期适应进化的结果,具有钙离子调控[17]、光照的聚集和反射[18]、植物防御[19]、抗逆境胁迫[20]等作用。苏志孟等对堇菜(Viola verecunda)叶片的研究发现,草酸钙是遭遇生境干旱时主要的束水物质[21];干旱区的灰绿藜(Chenopodium glaucum)、猪毛菜(Salsola collina)、碱地肤(Kochia scoparia var. Sieversiana)和荒漠中生长的灌木叶片草酸钙含量都很高[22-23]。由此可见,草酸钙对植物抗干旱生境胁迫有至关重要的作用,较高的草酸钙含量是反映植物具有较强抗旱性的重要指标。
白马雪山高寒草甸面积大,以线叶嵩草(Carex capillifolia)和蕨状嵩草(C. filispica)为建群种,然而,随着生境干旱化程度增加,高寒草甸面积快速减小[24]。原生植物群落退化演替,圆穗蓼(Bistorta macrophylla)等侵入种在部分生境取代原建群种成为优势种。在生境干旱化过程中,侵入种叶片解剖结构功能性状对植被发育和生态保护有着十分重要的意义。本研究沿生境干旱化梯度采集圆穗蓼为试验材料,通过研究叶片解剖结构和草酸钙含量随着生境干旱化的变化规律,以揭示叶解剖结构特征与生境适应的关系,以及叶解剖结构、草酸钙含量在不同土壤水分生境之间的差异。
1 材料与方法
1.1 研究区域概况
研究区域位于滇西北白马雪山垭口附近的高寒草甸,地理坐标为28°22′41″N,99°0′46″E,海拔4 262~4 328 m(图1a),属于寒温带山地季风气候,地形为高原山地,两山之间地段平缓,低凹区域有一条溪流[25]。高寒草甸分布于沟谷平坦处,并沿坡面分布于中下坡位,线叶嵩草(Carex capillifolia)和蕨状嵩草(C. filispica)为建群种,并且伴生有高山早熟禾(Poa alpina)、矮羊茅(Festuca coelestis)等(图1b)。近年来受全球气候变暖和人为放牧的双重干扰,其生境变化较大,原生植物群落退化演替,在部分生境中圆穗蓼、珠芽蓼(Bistorta vivipara)取代原建群种成为优势种(图1c)。
1.2 样方设置与材料采集
从溪边起至高寒草甸方向设置3个5 m×5 m的样地,1号样方为湿润生境样方,土壤表面湿润,用手指按压地表将渗出水分,该样地中圆穗蓼为偶见种;2号样方为半湿润样方,地表较湿润,但地表受压不会渗出水分,该样地中圆穗蓼为共优种;3号样方为干旱生境样方,地表干燥,且部分地表出现岩石裸露,该样地中圆穗蓼为优势种。用TS-TW土壤温湿度测定仪在样方内随机取5处样点测量土壤体积含水率,取平均值作为该样方的土壤体积含水率。调查记录每个样方的海拔、地形、地理坐标、优势种等信息(表1)。每个样方内选择5个1 m×1 m的小样方,使用数码相机以镜头垂直于地面进行拍摄获取图像,采用图像法测定圆穗蓼盖度[26]。实验材料采集于2023年8月,每个生境样方随机取长势均一的圆穗蓼50株,其中30株取叶片标记后放入FAA(formaldehyde-acetic acid-ethanol fixative)固定液(70% 乙醇∶冰醋酸∶甲醛=90∶5∶5)中保存;20株取叶片标记后放入液氮罐中保存。
1.3 叶解剖结构
将叶片自固定液取出,从叶片中间位置剪切含主脉和不含主脉的5 mm×5 mm的方块各1个。首先用70%酒精洗涤,经70%,85%,90%,100%系列叔丁醇-乙醇溶液脱水,每级2 h。而后用100%叔丁醇进行透明,共3次,每次2 h。透明后即刻浸蜡,将材料浸入纯石蜡中,放置在63℃烘箱中2~3 d。浸蜡后使用KD-BM II型包埋机包埋,用Leica RM 2145型切片机切片,切片厚度8~10 μm,切片在40℃烤片机上烤干,再用番红固绿对染,树胶封片。每个样地取15株样品,每株取1片完整无虫害的叶子,共15片叶,每片叶主脉和不含主脉方块各3个切片重复,3个样地共计270张切片。切片在Leica DM 500型光学显微镜下观察,并用LAS V4.4软件采集图像,共获得照片400余张。使用Image J软件测量以下指标:叶片厚度(μm),上下表皮厚度(μm),上下表皮角质层厚度(μm),栅栏组织厚度(μm),海绵组织厚度(μm),主脉厚度(μm),木质部厚度(μm),韧皮部厚度(μm),并计算栅海比(P/S)=栅栏组织厚度/海绵组织厚度。
1.4 晶体类型与含量测定
从液氮取出圆穗蓼叶片,研磨成粉末,将粉末样品置于载玻片上,使用X射线衍射仪(日本Rigaku Ultima IV)进行测试。实验条件设置:光源Cu-Kα射线,管电压40 kV,电流40 mA,测试范围10°~80°,每分钟为扫速5°[27]。测定结果表明晶体的化学成分是草酸钙(图2)。
精确称取待测植物叶片0.3 g(精确到 0.000 1 g),置于 15 mL透明离心管中,加入提前预冷过的 0.1 mol·L-1 盐酸 2 mL,匀浆 120 s,放置振荡器上振荡 1 min,室温超声提取 30 min,取出 4 000 r·min-1 离心 10 min,上清液转移到新的离心管中,重复提取一次,合并两次上清液,用 0.1 mol·L-1 盐酸定容至 5 mL,振荡混匀,取适当上清液过 0.22 μm微孔滤膜过滤,待液相色谱测定。采用安捷伦1200液相色谱系统(Agilent Technologies Inc.Santa Clara,USA)测定其含量。测定条件,色谱柱[VWDC18(250 mm×4.6 mm;0.5 μm)];柱温30℃;流速1 mL·min-1;流动相为0.1%磷酸∶甲醇(97.5∶2.5);进样量10 μL;波长210 nm[28]。使用草酸钙(购于sigma公司)作为标准,制作标准曲线,并根据标准曲线计算草酸钙含量。
1.5 数据分析
数据经Microsoft Excel软件记录并整理后,用SPSS 26. 0分析叶解剖结构、草酸钙含量与土壤含水率之间的相关性,判断哪些指标受土壤水分变化的影响较显著。对叶片解剖结构和草酸钙含量数据标准化处理,然后进行主成分分析,得到原始数据相关矩阵的特征值、方差贡献率、累计方差贡献率、荷载矩阵[29]。提取特征值大于1.00的因子作为主成分因子,确定主成分数量。以每个主成分所对应的特征值为权数,根据主成分表达式F=(λ1F1+λ2F2+λ3F3)/(λ1+λ2+λ3),计算综合评价模型F[30]。对综合模型中各因子的系数进行排序,系数大的确定为主成分,筛选出对物种耐旱性贡献率最大的指标。用Origin 2018绘图。
2 结果与分析
2.1 叶片解剖结构特征
圆穗蓼叶片上下表皮均由一层长椭圆形、圆形细胞紧密排列组成;栅栏组织由2层不规则的长柱形细胞排列而成,细胞间排列紧凑;海绵组织相对栅栏组织排列较为疏松,由近似椭圆的细胞组成,大小不一,细胞间隙大;栅栏组织与海绵组织中均有晶体分布(图3a)。叶片主脉区域由5~9个维管束环绕排列(图3b,c),木质部朝中心方向排列,韧皮部在木质部外面;维管束之间有薄壁组织(图3b)。
2.2 不同生境中叶解剖结构的差异
圆穗蓼叶片厚度为46.46~128.82 μm(图4a);上表皮角质层厚度为0.53~1.46 μm(图4b);上表皮厚度为4.91~16.24 μm(图4c);下表皮厚度为3.29~9.38 μm(图4c),这4个指标在生境间差异均显著(P<0.01,图4a~c),且干旱生境中最厚。下表皮角质层厚度为0.36~1.04 μm,生境间差异不显著(图4b)。栅栏组织厚度为19.44~54.65 μm;海绵组织厚度为15.29~58.05 μm;这2个指标在生境间差异均显著(Plt;0.05;Plt;0.01,图4d),且在干旱生境中最厚。栅海比为1.11~1.14,生境间差异不显著(图4e)。
圆穗蓼叶片主脉厚度为114.14~471.22 μm(图3a);维管束厚度为11.65~56.75 μm(图4f);木质部厚度为4.58~27.56 μm(图4g);韧皮部厚度为6.29~30.46 μm(图4g),这4个指标在生境间差异均显著(图4a,f,g,Plt;0.01),且在干旱生境中最厚。
2.3 不同生境中草酸钙含量的差异
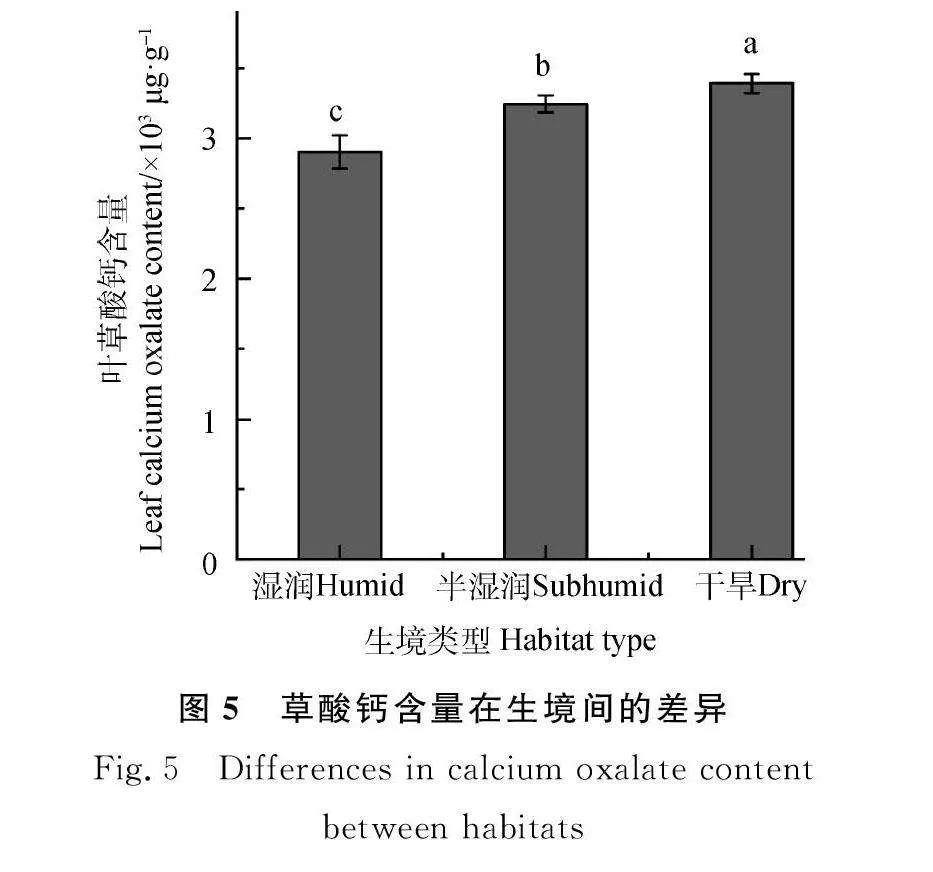
圆穗蓼叶片内的草酸钙含量为2 785.19~3 486.74 μg·g-1,干旱生境草酸钙含量显著高于湿润和半湿润生境(Plt;0.01,图5)。
2.4 叶片耐旱指标分析
圆穗蓼耐旱性3个主成分的贡献率分别为64.11%,9.55%,8.44%,累计贡献率为 82.10%(表2)。因此,3个主成分可以代替原始所有指标的绝大部分耐旱信息。F1中上表皮厚度、海绵组织厚度和下表皮厚度具有较高荷载,主要反映圆穗蓼光合作用和保水能力方面的信息。F2中栅海比和草酸钙含量具有较高荷载,主要反映圆穗蓼贮水和光合作用能力方面的信息。F3中上表皮角质层厚度荷载比较高,上角质层有利于减少植物的蒸腾作用,越厚保水性越强。根据主成分的因子荷载及特征值计算综合模型并排序,得到对圆穗蓼耐旱性贡献最大的3个指标是草酸钙含量、上表皮厚度和海绵组织厚度。
3 讨论
3.1 叶解剖结构特征与生境适应的关系
圆穗蓼在高寒草甸退化过程中,从偶见种到优势种,这其中与叶片的解剖结构特征密切相关。第一,叶肉中栅栏组织发达,与干旱生境中野生胡杨(Populus euphratica)发达的栅栏组织相似[31],以此增强植株的耐旱能力。第二,圆穗蓼叶片主脉的组成十分特殊,主脉区域由5~9个维管束环绕排列,木质部朝中心方向分布。不同于常见的旱生植物叶片主脉仅有1个维管束[32],木质部朝着上表皮方向排列,韧皮部朝着下表皮方向排列。推测圆穗蓼主脉中维管束这种特殊的分布方式,使得维管束数量更多,在一定程度上增加了输水通道的数量,极大增加了输水效率,使得植株生境适应能力更强,为其在干旱生境成为优势种提供保障。
3.2 叶片解剖结构对生境干旱化的适应
圆穗蓼叶片各解剖结构都在干旱生境中最厚。表皮有保护叶片组织结构的功能,它是叶片组织对抗复杂外部环境的重要屏障[33],角质层有利于反射强光、减少水分蒸发[34]。在生境干旱过程中,圆穗蓼叶片增加上、下表皮和上表皮角质层厚度可以减少高温带来的伤害,保持叶片水分。表皮和角质层同属于保水结构,而上表皮厚度对圆穗蓼的耐旱能力贡献较大,表明在干旱生境下圆穗蓼是增加对保水结构的投入以减少水分散失。栅栏组织是主要的光合作用场所[35-36],圆穗蓼叶片增加栅栏组织厚度以此加快光合作用效率,与抗旱性强的茶树一致[37]。下表皮角质层厚度不受土壤水分的影响,推测圆穗蓼叶背面的绒毛,在一定程度上削弱了下表皮角质层的作用。理论上较小的栅海比可以减弱叶片对光照的吸收,减少过剩光能对叶片的破坏[38],但土壤水分没有影响到圆穗蓼叶片的栅海比,而海绵组织对圆穗蓼的耐旱性贡献大,这与栅栏组织的协同增厚密切相关,以此提高维管束到表皮间的水分输送效率[39]。相较于减少对过剩光能的吸收,圆穗蓼更倾向于增加对输水效率的投入来抵御干旱胁迫。这些叶片组织结构的同时增厚,使得圆穗蓼在干旱生境中有了更厚的叶片,极大增强了叶片抵御外界复杂环境的能力。
主脉各解剖结构均在干旱生境中最厚。木质部与韧皮部主要是输送水分与养分的结构,其越厚则表明输送效率越高[40-41],如在面临干旱生境时费菜(Sedum aizoon)有更厚的木质部和韧皮部,因此水分和养分输送效率高于同生境的其他植物,适应能力也更强[42]。圆穗蓼也是如此,在干旱胁迫下增加木质部和韧皮部的厚度,以拓宽输送通道的方式增大输送效率抵御生境胁迫。叶片主脉除具有输送水分和养分的功能外,还有保水作用[43],生长在高寒草甸的圆穗蓼为了适应高海拔会出现的植物生理缺水这一现象[44],增加叶片主脉厚度以增强保水性,以确保植株不受生理缺水的迫害。
3.3 叶片草酸钙含量对生境干旱化的响应
干旱生境中草酸钙含量最高,且草酸钙含量对圆穗蓼耐旱能力贡献大。首先,草酸钙能通过调节渗透势维持植物细胞的水分平衡,在荒漠植物群落的建群种比典型草原建群种的草酸钙含量高,荒漠地区干旱程度高,草酸钙能提供水分使得植物抗旱能力更强[45]。圆穗蓼同样如此,随着生境干旱化,增加草酸钙含量,以此为植物内部保留更多的水分,是抵御干旱胁迫的一种策略。其次,草酸钙可以作为植物的动态碳库[46],生境干旱时,圆穗蓼增加海绵组织厚度,不利于外界CO2等气体从气孔下室到光合作用场所的传导,叶片内的草酸钙还可以提供CO2,维持植物的光合作用,因此圆穗蓼植株在水分亏缺条件下光合作用不受影响,进而保持良好的生长状态,从而占据更多生存空间。
4 结论
圆穗蓼发达的栅栏组织和主脉呈环形排列的维管束,为其适应干境生境提供了结构基础。叶肉组织结构增厚提供生理活动的基础;保水结构(表皮、角质层)增厚为叶片保留水分;输水结构(维管束,木质部)增厚,保证更高的输水效率;草酸钙含量的增加为光合作用和水分贮存提供条件。叶片保水和输水结构功能的增强,并与草酸钙含量协同响应干旱生境,使圆穗蓼的生境适应能力更强,在高寒草甸生境干旱化中取代原来建群种,从而成为优势种。
参考文献
[1] OLARANONT Y,STEWART A B,TRAIPERM P. Effects of crude oil on plant growth and leaf anatomical structures in a common coastal plant[J]. International Journal of Phytoremediation,2021,23(2):162-170
[2] 高钿惠,尚佳州,宋立婷,等. 小叶杨叶片光合特性与解剖结构对干旱及复水的响应[J]. 中国水土保持科学,2021,19(6):18-26
[3] 李萍. 光照强度对3种大型卷瓣凤梨叶片解剖结构及光合色素含量的影响[J]. 西北植物学报,2020,40(12):2065-2074
[4] 王坤,韦晓娟,李宝财,等. 金花茶组植物叶解剖结构特征与抗旱性的关系[J]. 中南林业科技大学学报,2019,39(12):34-39
[5] MONJE P V,BARAN E J. Characterization of Calcium Oxalates Generated as Biominerals in Cacti[J]. Plant Physiology,2002,128(2):707-713
[6] YANG Z,ZHU Q,ZHAN W,et al. The linkage between vegetation and soil nutrients and their variation under different grazing intensities in an alpine meadow on the eastern Qinghai-Tibetan Plateau[J]. Ecological Engineering,2018,110:128-136
[7] ZHANG Y,LIU X,CONG J,et al. The microbially mediated soil organic carbon loss under degenerative succession in an alpine meadow[J]. Molecular Ecology,2017,26(14):3676-3686
[8] 田培林,刘师,李登武. 祁连圆柏针叶解剖结构对高原寒旱环境的适应性分析[J]. 西北植物学报,2022,42(6):1030-1041
[9] 吴青松,刘英卉,李硕,等. 乌苏里鼠李茎叶的解剖结构及其生态适应性[J]. 植物研究,2023,43(3):461-469
[10]马静,贺熙勇,陶亮,等. 基于叶片解剖结构的澳洲坚果种质资源抗旱性评价[J]. 热带作物学报,2023,44(7):1392-1399
[11]周雨珩,刘慧,张世柯,等. 基于叶片解剖性状探究29种植物对热带珊瑚岛的适应策略[J]. 热带亚热带植物学报,2023,31(6):747-756
[12]王维睿,苏世平,李毅,等. 6个地理种群红砂(Reaumuria soongocica)叶片生态解剖特征及抗旱性评价[J]. 中国沙漠,2015,35(4):895-900
[13]LIU W,ZHENG L,QI D. Variation in leaf traits at different altitudes reflects the adaptive strategy of plants to environmental changes[J]. Ecology and Evolution,2020,10(15):8166-8175
[14]韩喆,张永强,张浩浩,等. 干旱胁迫对伊犁绢蒿幼苗生长及叶片解剖结构的影响[J]. 草地学报,2024,32(1):105-112
[15]肖晴,林轸荣,姜风岩,等. 放牧强度对高寒草甸优势植物叶片解剖结构的影响[J]. 草地学报,2023,31(10):3018-3025
[16]李秀丽,张文君,鲁剑巍,等. 植物体内草酸钙的生物矿化[J]. 科学通报,2012,57(26):2443-2455
[17]SAMEH S,MOHAMMAD M G,EL-KEBLAWY A A,et al. Mechanical and phytochemical protection mechanisms of Calligonum comosum in arid deserts[J]. Plos One,2018,13(2):e0192576
[18]BAUER P,ELBAUM R,WEISS I M. Calcium and silicon mineralization in land plants:Transport,structure and function[J]. Plant Science,2011,180(6):746-756
[19]KORTH K L,DOEGE S J,PARK S H,et al. Medicago truncatula mutants demonstrate the role of plant calcium oxalate crystals as an effective defense against chewing insects[J]. Plant Physiology,2006,141(1):188-95
[20]袁佳秋,孙大伟,杨玲,等. 东京四照花钙组分与渗透调节物质对盐胁迫的响应[J]. 生态环境学报,2023,32(4):687-696
[21]苏志孟,张习敏,马琳,等. 堇菜叶片草酸钙晶体与水分维持的关系[J]. 广西植物,2019,39(6):720-728
[22]李梦琦,何兴东,杨祥祥,等. 半干旱区两种草原类型植物体内草酸钙研究[J]. 草地学报,2018,26(6):1421-1427
[23]CI H C,LIE X D,LI R,et al. Characteristics of plant calcium fractions for 25 species in Tengger Desert[J]. Sciences in Cold and Arid Regions,2010,2(2):168-174
[24]霍佳娟,朱珏妃,宋明华,等. 青藏高原高寒草甸退化演替进程中植被斑块特征[J]. 草地学报,2022,30(11):3113-3118
[25]彭新华,杨绕琼,尹云丽,等. 滇西北白马雪山高山松(Pinus densata)径向生长对气候因子的响应[J]. 生态学报,2023,43(21):8884-8893
[26]关法春,梁正伟,王忠红,等. 方格法与数字图像法测定盐碱化草地植被盖度的比较[J]. 东北农业大学学报,2010,41(1):130-133
[27]杨世颖,邢逞,张丽,等. 基于粉末X射线衍射技术的固体制剂晶型定性分析[J]. 医药导报,2015,34(7):930-934
[28]刘晓婷,姚拓,马亚春,等. 植物根际促生菌发酵液中植物激素含量的液液萃取-高效液相色谱测定法[J]. 草业科学,2022,39(10):2222-2228
[29]朱凯琳,李嘉宝,陈昕. 龙苍沟国家森林公园7种花楸属植物的叶解剖特征及其环境适应性[J]. 植物研究,2022,42(2):174-183
[30]杨春娇,韩雨圳,李忠馗,等. 藏东南高寒草甸两种嵩草根系导管解剖结构对生境干旱化的响应[J]. 草业学报,2022,31(2):76-87
[31]丁伟,杨振华,张世彪,等. 青海柴达木地区野生胡杨叶的形态解剖学研究[J]. 中国沙漠,2010,30(6):1411-1415
[32]刘慧斌,朱周俊,赵君茹,等. 6个高州油茶无性系叶片形态结构及耐热性比较[J]. 热带作物学报,2023,44(4):737-745
[33]BINKS O,MEIR P,ROWLAND L,et al. Limited acclimation in leaf anatomy to experimental drought in tropical rainforest trees[J]. Tree Physiology,2016,36(12):1550-1561
[34]CHEN M,ZHU X,ZHANG Y,et al. Drought stress modify cuticle of tender tea leaf and mature leaf for transpiration barrier enhancement through common and distinct modes[J]. Scientific Reports,2020,10(1):6696
[35]ZHU J,ZHU H,CAO Y,et al. Effect of simulated warming on leaf functional traits of urban greening plants[J]. BMC Plant Biology,2020,20(1):139
[36]吴建波,王小丹. 高寒草原优势种紫花针茅叶片解剖结构对青藏高原高寒干旱环境适应性分析[J]. 植物生态学报,2021,45(3):265-273
[37]仇杰,高超,罗洪发. 贵州西北喀斯特区古茶树叶片解剖结构及抗旱性评价[J]. 西北植物学报,2023,43(7):1170-1184
[38]VELIKOVA V,ARENA C,IZZO L G,et al. Functional and Structural Leaf Plasticity Determine Photosynthetic Performances during Drought Stress and Recovery in Two Platanus orientalis Populations from Contrasting Habitats[J]. International Journal of Molecular Sciences,2020,21(11):3912
[39]GONZALEZ P L,RAVETTA D A. Relationship between photosynthetic rate,water use and leaf structure in desert annual and perennial forbs differing in their growth[J]. Photosynthetica,2018,56(4):1177-1187
[40]刘万鹏,李悦煊,李志勇,等. 内蒙古四种不同生态类型扁蓿豆叶片解剖性状与抗旱性关系的研究[J]. 中国草地学报,2023,45(6):32-43
[41]HU Y,YANG L,GAO C,et al. A comparative study on the leaf anatomical structure of Camellia oleifera in a low-hot valley area in Guizhou Province,China[J]. Plos One,2022,17(1):e0262509
[42]李雪,吴青松,许少祺,等. 景天科3种植物的叶片形态结构与抗旱性评价[J]. 东北师大学报(自然科学版),2023,55(3):114-121
[43]MALES J. Adaptive variation in vein placement underpins diversity in a major Neotropical plant radiation[J]. Oecologia,2017,185(3):375-386
[44]刘梦瑶,李伟琼,王铁娟. 差不嘎蒿茎叶解剖特征及其与气候因子的关系[J]. 草地学报,2021,29(5):1118-1124
[45]徐平宜,张嘉悦,何兴东. 过渡带近距离同地分布不同植物群落建群种叶片草酸钙特征[J]. 草地学报,2021,29(5):929-935
[46]TOOULAKOU G,GIANNOPOULOS A,NIKOLOPOULOS D,et al. Alarm Photosynthesis:Calcium Oxalate Crystals as an Internal CO2 Source in Plants[J]. Plant Physiology,2016,171(4):2577-2585
(责任编辑 彭露茜)
