铁皮石斛DobHLH51基因克隆及表达特性分析
2024-01-01崔迪孟丽君葛秀丽郎朗刘博婷刘羽佳



关键词:铁皮石斛;bHLH转录因子;DobHLH51基因;生物信息学分析;非生物胁迫;表达特性分析
铁皮石斛(Dendrobium officinale Kimura et Mi-go)是兰科(Orchidaceae)石斛属(Dendrobium)多年生附生草本植物,是一种重要的药用资源,在我国已有两千多年的药用历史,早在东汉末年《神农本草经》中就有记载并被奉为“上品”。因具有“滋阴清热、益胃生津”之功效,铁皮石斛历来被视为石斛中的瑰宝。现代药理学分析表明,铁皮石斛具有调节免疫力、降血糖、抗氧化、抗肿瘤、降血脂、抗衰老、对胃肠道炎症有疗效等较高的药用和保健价值。铁皮石斛的野生资源多分布于东亚、东南亚、澳大利亚等气候温暖湿润的国家或地区,在我国则主要分布在浙江、云南、贵州、广东等地。野生铁皮石斛繁育率低,加之过度采挖和生态环境的恶化,野生资源受到严重破坏:人工种植虽然解决了野生石斛资源稀缺的问题,但由于盲目引种导致的种苗混杂以及种植技术不规范等,其品质难以保证。因此,培育铁皮石斛优良种质对保障中药材原料及品质、确保临床疗效和用药安全至关重要。
bHLH (basic/helix-loop-helix)家族是植物中第二大类转录因子家族,其结构域均由60个氨基酸组成,包含两个高度保守且功能不同的保守区域,分别位于N端和C端。其中,N端的由13~17个亲水性碱性氨基酸组成的碱性区域可以特异地与DNA序列结合,进而参与调控很多重要的转录过程:C端的疏水性碱性氨基酸组成螺旋一环一螺旋(HLH)区域,并通过与其他蛋白的疏水性氨基酸结合形成同源或异源二聚体调控下游基因表达。另外,这两个结构域之间还存在着相互联系和相互作用,共同调节细胞中许多重要基因的表达,如植物开花、种子休眠和根毛发育等的调控基因。
bHLH家族成员的作用广泛而复杂,在植物次生代谢调节过程中也起到非常重要的作用。例如,水仙花(Chinese narcLssus)NtbHLH1与Nt-MYB6相互作用能够直接促进花青素的生物合成:Aa6119和Aa7162作为典型的黄花蒿(Ar-temzsza annual)bHLH转录因子,通过两种机制协同并正向调控青蒿素在黄花蒿中的生物合成,一种是通过Aa6119与ADS启动子的直接结合来促进ADS基因的转录激活,另一种是通过Aa6119与Aa7162形成异源二聚体协同激活青蒿素生物合成基因和毛状体发育基因的表达。此外,bHLH家族成员与植物响应非生物胁迫(如低温、干旱等)有关,当植物感受到外界环境变化时,会将这种变化通过信号通路向下游传导,从而激发非生物胁迫响应相关基因的表达,提高对环境的适应能力。如花生(Arachis hypogaea)Ah-bHLH112基因的过表达提高了同源转基因植株幼苗期和成株期的抗旱能力,推测AhbHLH112可能通过增强活性氧的清除能力,调控过氧化物酶(POD)介导的H202稳态,进而参与ABA依赖的应激反应途径,在干旱胁迫响应中起积极作用;三叶橙(Trifoliate orange)的PtrbHLH66基因在拟南芥(Arabidopsis thaliana)中的过表达促进了转基因植株的种子萌发和根系生长,提高了脯氨酸和ABA含量以及抗氧化酶活性,降低了丙二醛(MDA)和活性氧(ROS)的积累,进而增强了转基因拟南芥的抗旱性;水稻(Oryza sativa)的OsbHLH57基因通过正向调控海藻糖合成、ROS代谢和CBFs/DREBs -依赖性途径提高水稻的耐寒性;在拟南芥中过表达仙人掌(FagopyrumtatarLcum)的FtbHLH2基因,能够显著增强转基因拟南芥的抗寒能力。可见,bHLH转录因子在植物非生物胁迫应答中发挥重要作用,深入研究bHLH基因功能及其表达调控机制对于提高植物的抗逆性、选育优良种质、提高作物产量等方面具有重要意义。
目前,有关铁皮石斛bHLH转录因子的研究报道相对较少。Wang等于2020年从铁皮石斛基因组中鉴定出98个bHLH基因家族成员,并将其分成18个分支。Yu等克隆了茉莉酸甲酯响应转录因子DobHLH4基因,研究发现该转录因子能够调控DoTPS10基因表达进而促进铁皮石斛花发育过程中芳樟醇的生物合成。张红瑞和理雅等分别克隆了铁皮石斛DobHLH96和DcbHLH14基因,并对这两个基因在非生物胁迫下的表达模式进行了分析,推测这两个基因可能通过依赖于ABA信号转导途径响应低温和干旱胁迫,但响应机制尚不明确。本研究从铁皮石斛中克隆了DobHLH51基因编码区(CDS)序列,并对其进行生物信息学和组织表达特异性分析,明确该基因在低温、干旱和ABA处理下的表达特性,为深入研究DobHLH51基因功能及其在非生物胁迫应答中的表达调控机制奠定理论基础,也为通过现代育种技术培育优质抗逆的药用铁皮石斛优良品种提供基因资源。
1材料与方法
1.1试验材料
供试植物材料为广东丹霞铁皮石斛,取样于韶关市石斛工程开发中心。2x Taq PCR Mix、All-in-One First-Strand Synthesis MasterMix、Fast-Pure Gel DNA Extraction Mini Kit购自北京兰博利德生物技术有限公司、T4 DNA连接酶、pMD18-T载体、Real Time PCR试剂盒均购自宝生物工程(大连)有限公司;PCR引物合成委托生工生物工程(上海)股份有限公司完成。
1.2试验方法
试验于2021-2022年在广东省韶关市韶关学院生物与农业学院分子生物学与基因工程研究室进行。
1.2.1样品采集与处理铁皮石斛蒴果消毒后接种于培养基中无菌培养6个月,选取生长状态良好且长势一致的幼苗(株高约5~6cm)用于胁迫处理。设低温处理组(4℃恒温恒湿培养箱中培养)、模拟干旱处理组(20%PEG-6000浇灌)和ABA处理组(100umol/L ABA浇灌),以未经任何处理的正常条件下的无菌幼苗为对照组。各组均在处理第1、3、6、9、12、24 h取叶组织样品,经液氮速冻后,保存于-80℃超低温冰箱,用于胁迫下的基因表达分析。分别采集处于花期的一年生铁皮石斛的根、茎、叶和花器官,经液氮速冻后超低温保存,用于基因组织表达分析。每个样品3次生物学重复。
1.2.2总RNA提取与cDNA合成采用改良的CTAB法分别提取上述样品总RNA,利用Nan-odrop微量分光光度计检测其浓度。参照All-in-One First-Strand Synthesis MasterMix剂盒说明书反转录合成单链cDNA。
1.2.3基因克隆与阳性茵斑鉴定根据NCBI(https://www.ncbi. nlm. nih. gov)数据库已知的云南铁皮石斛bHLH51同源序列(基因ID:LOC110107930)设计DobHLH51基因特异性扩增弓I物(F:5'-ATGAAGAACTCTTCAAATTTCT -3';R:5'-CTAAACAGACATATTGAAATAG-3').PCR扩增产物经琼脂糖凝胶电泳检测后,胶回收连接至pMD18-T线性载体,并转化大肠杆菌(Escherichia coli) DH5a感受态细胞,筛选单克隆菌液进行PCR鉴定后,选取阳性克隆送至委托公司测序验证。
1.2.4生物信息学分析在NCBI数据库提交克隆的DobHLH51基因的氨基酸编码序列,通过Blastp比对筛选相似性高的其他物种同源序列,利用MEGA11软件的邻接法(neighbor-joining,NJ)构建系统发育进化树。利用DNAMAN软件对DobHLH51及其同源蛋白进行氨基酸多序列比对:利用NCBI-CDD在线预测DobHLH51蛋白结构域;运用ExPASy ProtParam预测蛋白理化性质;使用SOPMA进行蛋白二级结构分析,采用ProtScale进行亲疏水性分析:采用SWISS -MOD-EL预测分析蛋白三级结构,采用PDBsum对蛋白三级结构合理性评估;使用TMHMM和SignalP5.0预测蛋白跨膜区和信号肽,利用NetPhos 3.1在线软件预测蛋白磷酸化位点。通过PlantCARE网站预测启动子顺式作用元件。
1.2.5组织表达特性分析从NCBI数据库中下载未经任何处理的8个铁皮石斛组织的表达数据,分别是花蕾、花柱、唇瓣、萼片、叶、茎、灰白根和绿根尖。基因表达量用FPKM(fragments perkilobase per million mapped reads)表示,并用TBtools软件进行可视化处理,分析DobHLH51基因在不同组织器官中的表达情况。
1.2.6不同胁迫处理下的转录组分析将经低温、干旱或ABA处理不同时间的铁皮石斛样品的cDNA模板送至深圳华大基因科技有限公司完成高通量测序。去除所得原始测序序列中的接头和低质量Rawreads.以保证信息分析质量,最终得到Cleanreads;利用Qubit 2.0将Clean reads与铁皮石斛基因组参考序列进行比对,并使用Cufflinks进行组装从而获得转录组数据,基因表达量以FPKM表示,分析铁皮石斛DobHLH51基因在低温、干旱和外源ABA处理下的表达模式。
1.3数据处理与分析
采用WPS 2019软件对数据进行处理,使用SPSS21.0进行统计分析,用Duncan's法进行多重比较,使用GraphPad Prism 9.5软件作图。
2结果与分析
2.1DobHLH51基因克隆与序列分析
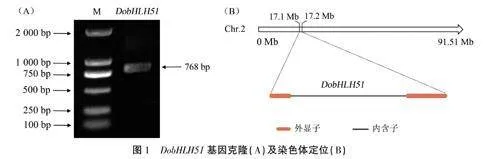
由图1、图2可见,以广东丹霞铁皮石斛叶组织cDNA为模板,利用特异性引物进行PCR扩增,克隆到目的条带。测序获得DobHLH51基因CDS序列全长为768bp,编码255个氨基酸。与参考序列对比,共有13个碱基差异,包括5个同义替换、5个非同义替换和3个碱基缺失。基因结构和染色体定位结果显示.DobHLH51基因定位于2号染色体,含有2个外显子和1个内含子。
2.2DobHLH51蛋白生物信息学分析
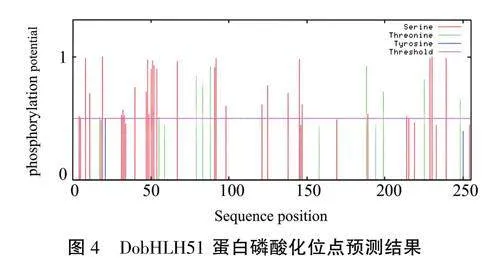
蛋白结构分析结果显示,DobHLH51蛋白含有bHLH家族保守结构域(cl00081)和ACT结构域(cd04873),属于bHLH转录因子亚家族(图3A);DobHLH51分子式为相对分子量为28.03kDa,理论等电点为9.71,不稳定指数达58.01,在第27位亮氨酸(Leu)处具有亲水性最大值(1.567),在第74位精氨酸(Arg)处具有亲水性最小值(-3.033),亲水性平均值为-0.334(图3B),说明DobHLH51属于亲水性不稳定蛋白。DobHLH51蛋白二级结构由41.57%a-螺旋、1.96%折叠、11.76%延伸链和44.71%无规则卷曲组成(图3C),主要以a-螺旋和无规则卷曲为主;其三级结构(图3D)与墨兰bHLH30转录因子相似度高达81.82%,且该模型符合立体化学的规则,蛋白构象合理可靠(图3E)。此外,DobHLH51蛋白无信号肽和跨膜结构区;但含有52个蛋白磷酸化位点,其中包括38个丝氨酸(Ser)、12个苏氨酸(Thr)及2个酪氨酸(Tyr)位点(图4)。
2.3DobHLH51蛋白的同源性和系统发育分析
以DobHLH51蛋白为目的序列,在NCBI数据库中筛选出9个不同物种的同源性较高的序列进行比对分析,其中同源性最高的是黄石斛(Den-drobium catenaturn)bHLH蛋白,相似度为97.66%;其次是金钗石斛(Dendrobium nobile)和鼓槌石斛(Dendrobium chrysotoxum)的bHLH蛋白,相似度分别为96.47%和94.12%。多重序列比对结果(图5)显示,DobHLH51氨基酸序列与其他物种bHLH蛋白的氨基酸序列同源性较高,均在N端第56~114个氨基酸残基位置含有bHLH蛋白结构域。进化树分析结果(图6)显示,DobHLH51与黄石斛和金钗石斛bHLH的聚类关系最近,与同源序列比对分析结果一致。
2.4DobHLH51基因组织表达特性分析
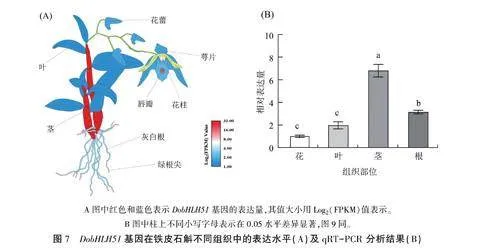
转录组分析结果(图7A)表明,DobHLH51基因在铁皮石斛不同组织器官中的相对表达量存在明显差异,在茎中的相对表达量最高,在根组织(绿根尖和灰白根)中的表达量次之,在叶中的表达量最低。qRT-PCR分析结果(图7B)进一步验证了DobHLH51基因在铁皮石斛不同组织中的表达存在差异,同样以茎中的表达量最高,是叶的3.45倍:其次是根中的,相对表达量是叶的1.6倍;在花中的表达量最低。推测该基因可能在铁皮石斛茎的生长发育中发挥着重要的调控作用。
2.5DobHLH51在低温、干旱及ABA处理下的表达特性分析
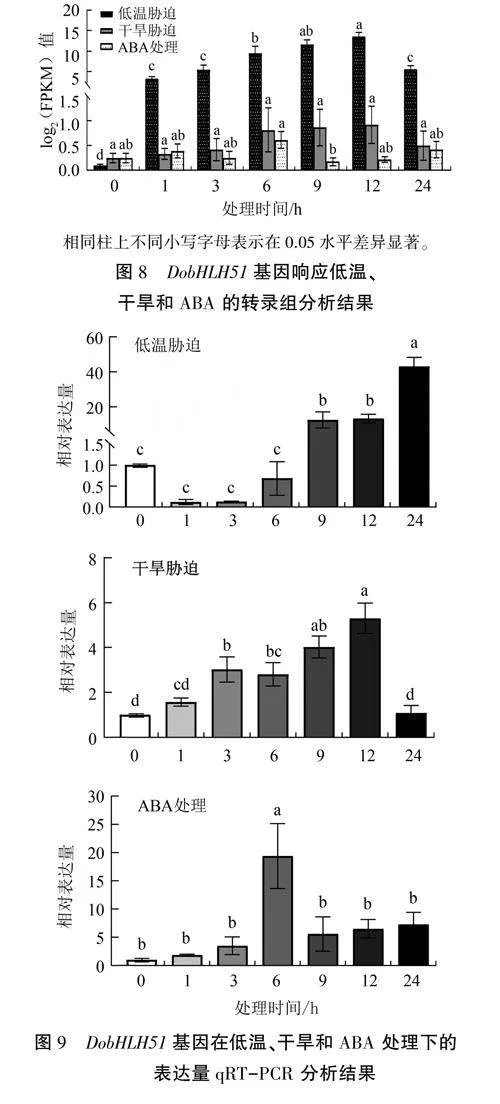
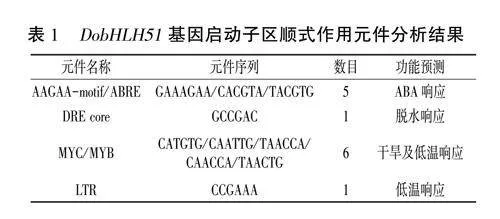
PlantCARE分析显示,DobHLH51基因起始密码子上游2kb启动子中含有脱水、干旱、低温及ABA响应等顺式作用元件(表1),因此,对DobHLH51基因在低温、干旱和ABA处理下的表达特性进行分析。转录组分析结果(图8)显示,DobHLH51基因表达量对3种处理的响应整体上均随着时间的延长呈现先升高后下降趋势,其中低温和干旱处理12h表达量达到最高,而ABA处理6h表达量就达到峰值,表明DobHLH51基因明显受到低温、干旱和ABA诱导,提示该基因可能在非生物胁迫响应过程中发挥重要功能。qRT-PCR分析结果(图9)进一步验证了DobHLH51基因在低温、干旱和ABA处理下均可被诱导上调表达,其中低温处理后DobHLH51表达量先下降后升高,处理24h达到峰值,是处理前的43倍:干旱处理后DobHLH51表达量持续升高,处理12h达到最高值,是处理前的5.3倍:ABA处理6h DobHLH51表达量最高,是处理前的19.4倍。
3讨论与结论
bHLH家族作为植物第二大类转录因子,已在多种植物中被鉴定和克隆出来,并借助拟南芥、水稻等模式植物进行了功能研究,发现bHLH转录因子在植物生长发育和非生物胁迫响应过程中发挥着重要的调控作用。铁皮石斛中共鉴定出98个bHLH基因家族成员,但对其响应非生物胁迫方面的作用及机制鲜少报道。本研究克隆了铁皮石斛DobHLH51基因,其CDS全长768bp,编码255个氨基酸,除含有bHLH保守结构域外,还具有ACT结构域,该结构域能够结合特定的小分子配体来提供变构调节,以发挥传感器蛋白功能。DobHLH51为亲水性不稳定蛋白,其三级结构与墨兰bHLH30同源性高达81.82%,且在进化上与黄石斛和金钗石斛bHLH亲缘关系最近,表明DobHLH51蛋白序列和结构在兰科植物中进化相对保守。此外,DobHLH51肽链中含有52个磷酸化位点,其中38个Ser位点,推测该蛋白发生磷酸化时可能以Ser磷酸化为主,从而在多种生物学进程和分子功能上发挥着重要的调控作用。
以往的研究显示,bHLH转录因子在植物不同发育日寸期和不同组织器官中的表达具有日寸空特性,对植物的形态建成和生长发育起着重要作用。本研究克隆的DobHLH51基因在铁皮石斛不同组织器官中的表达水平也存在明显差异,在茎中的表达量最高,提示该基因在铁皮石斛营养器官形成、养分吸收和运输等过程中发挥重要功能。此外,bHLH基因及其参与的信号通路在植物响应低温、干旱、高盐等胁迫时发挥重要调控作用。如葡萄(Vitis vinifera)VvbHLH1通过增加转基因拟南芥中黄酮类化合物的积累并增强其ABA信号传导,提高了转基因植株对非生物胁迫的耐受性;密罗木(Myrothamnus flabellifolia)MfbHLH38基因通过依赖于ABA的信号途径提高保水能力、调节渗透平衡、减轻胁迫引起的氧化损伤等以增强转基因拟南芥植株对干旱和盐胁迫的耐受性。本研究发现,DobHLH51基因启动子区含有低温、干旱和脱水响应以及ABA应答元件,且可被低温、干旱和ABA处理诱导显著上调表达:但其在低温和干旱胁迫下的表达模式一致,与ABA诱导下表达模式不同,提示该基因响应低温和干旱胁迫可能通过不依赖于ABA的信号转导途径,但具体的调控机制尚需进一步研究。
值得关注的是.有研究发现bHLH转录因子主要通过ICE-CBF途径结合并激活靶基因的表达,提高活性氧清除能力及联合光信号、激素信号等机制来增强植物的抗寒能力。本研究克隆的DobHLH51启动子中含有能够被CBFs特异结合的LTR元件,该元件是植物低温响应过程中的关键因子,推测DobHLH51基因可能通过CBFs介导的ICE-CBF信号途径参与低温应答过程,但具体响应机制尚未可知。因此,需要对DobHLH51基因响应干旱和低温胁迫的分子调控机制进行深入研究。今后本课题组将通过基因过表达和基因敲除实验对DobHLH51基因的功能做进一步分析和验证,以期为阐明该基因在铁皮石斛抗逆中的作用机制奠定基础。
