城市人工浅水湖泊底栖动物群落结构及影响因素分析
2024-01-01莫竞瑜池仕运王竹王瑞魏秘喻霈雯曾小康潘静怡胡菊香





摘要:探讨梅溪湖底栖动物群落结构及其影响因素,为城市人工湖泊水生态系统修复后的运营维护管理提供理论依据。2020-2021年在梅溪湖设置14个采样站位(敞水区7个,滨岸带7个),按季节开展底栖动物调查。研究结果表明,梅溪湖底栖动物种类达75种,滨岸带种类数高于敞水区。整个湖区全年优势种为米虾(Caridina sp.)、铜锈环棱螺(Bellamya aeruginosa)、椭圆萝卜螺(Radix swinhoei)、大脐圆扁螺(Hippeutis umbilicalis)、霍甫水丝蚓(Limnodrilus hoffmeisteri)和沼虾(Macrobrachium sp.)。敞水区和滨岸带群落结构存在显著差异,除霍甫水丝蚓外,全年优势种是导致2个区域群落结构差异的主要分类单元。梅溪湖底栖动物现存量和多样性季节间无显著差异,但敞水区和滨岸带之间差异显著,滨岸带远高于敞水区,表明水生植物群落的构建有利于底栖动物种群的壮大和多样性的提高。CCA结果表明,氨氮(AN)和营养水平(TLI)对敞水区和滨岸带的底栖动物群落结构均有较大影响。CoIA分析结果显示,米虾、大脐圆扁螺、铜锈环棱螺、沼虾、椭圆萝卜螺等物种与环境因子存在较为密切的联系,这些物种可以作为梅溪湖水环境条件变化的指示种。合理控制梅溪湖底栖动物群落结构,并维持较高的多样性水平,可有效促进其生态系统的正常运转。
关键词:滨岸带;敞水区;水生植物;底栖动物;富营养化;影响因子;城市湖泊
中图分类号:Q178.1" " " " 文献标志码:A" " " " 文章编号:1674-3075(2024)05-0058-09
城市人工湖泊具有雨水调蓄、水资源再利用、气候调节和增加生物多样性等生态服务功能,在城市景观设计和水生态文明建设规划中受到高度重视(王以尧等,2018)。为改善城市生态环境、提高城市生活品质,近年来城市人工湖泊的建设得到了快速发展(窦明等,2019)。然而与自然湖泊相比,城市人工湖泊通常较浅,水域面积和水环境容量较小,水体循环缓慢,水体自净能力较差,在人类活动的强烈扰动及运营维护中,易发生各种水生态环境问题,如污染物以及底泥内源污染物难以降解,易致水质恶化、水体富营养化和藻华暴发(窦明等,2019)。
梅溪湖是湖南长沙市的一座人工湖泊,成湖之初水质较差。2017-2018年实施生态治理工程,2020-2021年调查显示,梅溪湖水体处于中度污染和中-富营养状态,藻类年均丰度超1×107个/L,有不定期藻华暴发风险(魏秘等,2022)。已有研究表明,生态修复可以降低湖泊沉积物中的总磷和有机物,增加沉水植物生物量和沉积物中微生物多样性,改善底栖动物群落状态,加快湖泊生态系统的恢复和重建进程(Liu et al,2022)。结合鱼类控制与沉水植物重建等修复措施能有效改善水质和水生态状况(吴家乐和刘正文,2019)。在众多影响浅水湖泊底栖动物的因素(水深、水位、透明度、水温、叶绿素a、沉水植物、水动力条件、湖泊形态、水质状况以及底质类型等)中(Shrestha et al,2021;沈忱,2012;王海军,2007;2019;陆文泽等,2022; Obolewski et al,2021),底栖动物群落的时空变化主要由营养物质(如总氮和氨氮)和栖息地因素(如沉积物和沉水植物)来解释(Mao et al,2023;Li et al,2023),其中,沉水植物分布是影响底栖动物的关键因素(谢志才等,2007)。然而,有研究表明,仅靠湖泊生态系统的自我恢复或仅种植沉水植物引入外来底栖物种,对底栖动物多样性的影响极为有限(钟非等,2007)。
梅溪湖水体营造初期,投放的底栖动物种类较为单一,主要为螺贝类和虾蟹(张小春等,2021)。如今,滨岸带形成水生植物景观带,局部水域特别是西部水域形成了大片“水下森林”,但管理运营方对底栖动物群落结构缺乏深入了解。先期投放的底栖动物物种对群落结构的后续演化有何作用?底栖动物现存量和多样性处于何种水平?水生植物是否促进底栖动物群落状态的改善?这些问题均需要在深入调查的基础上进行科学解答。相比浮游生物群落,底栖动物对水体环境条件的改善更为敏感(Mao et al,2023),在湖泊生态修复中起到十分重要的作用。本研究以梅溪湖滨岸带和敞水区底栖动物作为研究对象,探讨分析其群落结构及影响因素,以期为城市人工湖泊水生态系统修复后的运营维护管理提供理论依据。
1" "材料与方法
1.1" "研究区域
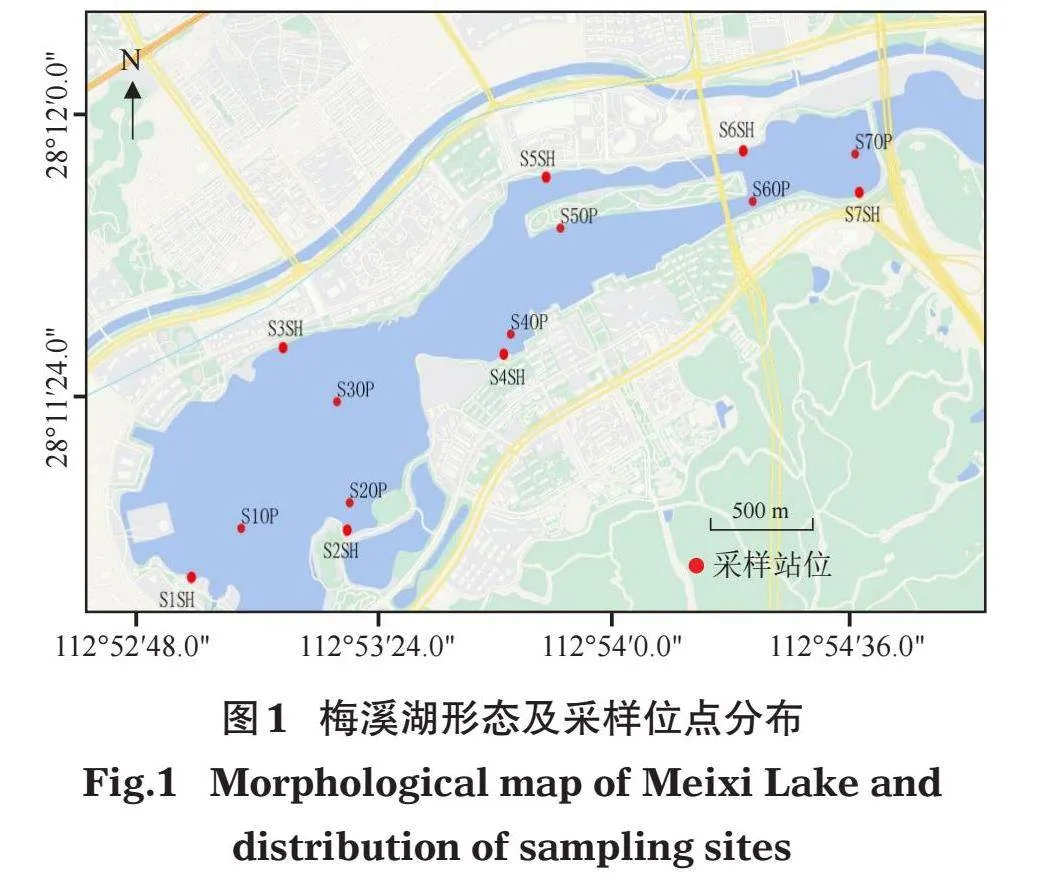
梅溪湖位于湖南长沙湘江新区,龙王港中游的右岸,是湘江新区核心区域的城市“海绵体”,具雨水调蓄、景观观赏等功能。梅溪湖最高运行水位35.40 m,正常蓄水位35.00 m,梅溪湖东湖建有泵站调控湖泊水位。正常蓄水位以下库容434.1万m3,湖面面积为200 hm2(二环以西面积约170 hm2),集雨面积12.31 km2。梅溪湖最大水深6 m,水深0~2 m的区域面积占湖区总面积32.4%。由于汇水面积、汇水量和换水量调节能力较小,梅溪湖受污染风险大,生态环境脆弱,发生富营养化的可能性较大(魏秘等,2022;张小春等,2021;2023)。
梅溪湖于2012年建成,2017-2018年实施的生态治理工程采取河蚌笼式挂放、水生植物种植和水生动物投放等措施。经生态治理,梅溪湖水质明显提升,滨岸带水生植物茂盛,初步形成了“水清岸绿”的风景(张小春等,2021)。梅溪湖湖区水生植物主要以沉水、挺水植物为主,还包括部分漂浮、浮叶植物。挺水植物多分布于西南部岸线,呈条带状;沉水植物多分布在水深0~2 m的西部湖区,东部湖区因水深gt;5 m,沉水植物相对较少。水生植物主要呈模块化分布,形成了多种单优势挺水植物群落,如香蒲群丛、梭鱼草群丛、再力花群丛、莲群丛、泽泻群丛、旱伞草群丛、水葱群丛、香菇草群丛和喜旱莲子草群丛;单优势沉水植物群落主要有苦草群丛、马来眼子菜群丛、轮叶黑藻群丛、金鱼藻群丛和菹草群丛;漂浮、浮叶植物群落主要有菱角群丛、荇菜群丛和粉绿狐尾藻群丛(张小春等,2021)。
1.2" "研究方法
采样时间为2020年11月(秋季)、2021年1月(冬季)、4月(春季)和7月(夏季)。根据梅溪湖形态和环境特征,自西向东设14个采样站位(图1),其中,敞水区7个(S1OP~S7OP),滨岸带7个(S1SH~S7SH)。敞水区(OP)采用彼得逊采泥器(面积1/16 m²)进行定量采样,每个采样站点采集 2~4个重复样合并成一个大样。滨岸带(SH)采用长柄矩形踢网采集,记录网口扫过的面积。野外将采集的粗样淘洗干净后装入封口袋中,带回实验室分拣处理。在实验室用10%的福尔马林溶液固定保存。标本鉴定参照底栖动物专业分类文献(蔡如星,1991;王洪铸,2002;Morse et al,1994),鉴定至尽可能低的分类单元,并用万分之一精度的电子天平称量湿重。采集底栖动物时同步采集水质样品,带回实验室后测定氨氮(AN)、总氮(TN)、总磷(TP)、叶绿素a(Chl-a)和高锰酸盐(CODMn),测定方法参照《水和废水监测分析方法》(魏复盛,2002)。
1.3" "数据分析
梅溪湖水体营养状态采用综合营养状态指数(TLI)进行定量评价(金相灿,1995),分级标准:TLIlt;30为贫营养;30≤TLI≤50为中营养;50lt;TLI≤60为轻度富营养;60lt;TLI≤70为中度富营养;TLIgt;70为重度富营养。选择Chl-a、TP、TN、CODMn作为评价参数,综合营养状态指数TLI计算公式为:
TLI=[j=1mWj×TLI(j)] ①
式中:TLI为综合营养状态指数;Wj为第j种参数的营养状态指数的相关权重;TLI(j)为代表第j种参数的营养状态指数。
以Chl-a作为基准参数,则第j种参数的归—化的相关权重计算公式为:
Wj = [r2ijj=1mr2ij] ②
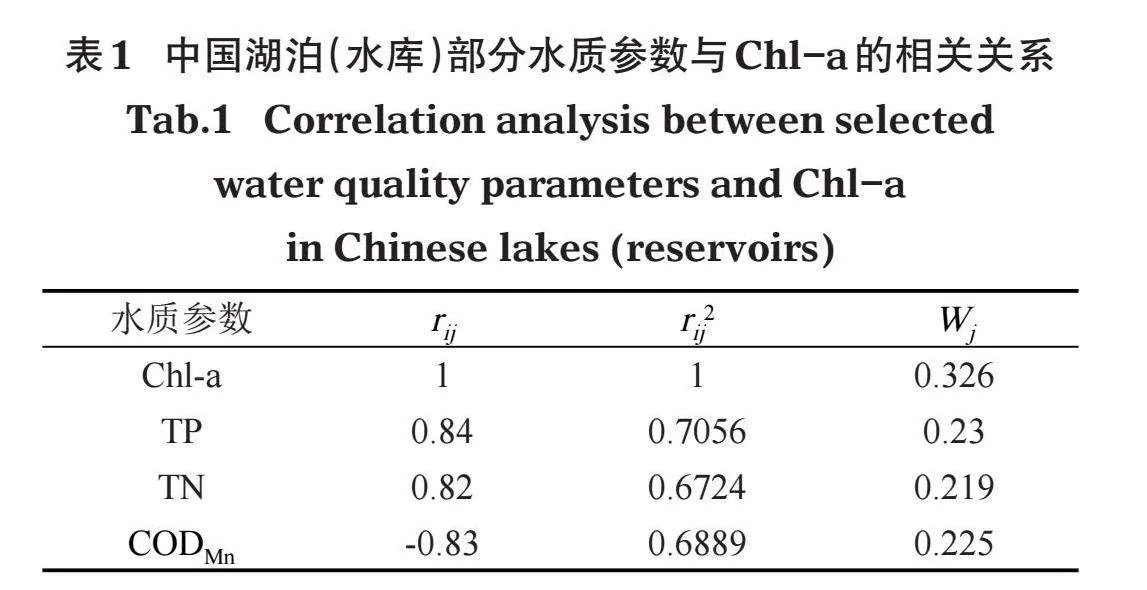
式中:rij为第j种参数与基准参数Chl-a的相关系数;m为评价参数的个数。中国湖泊(水库)的Chl-a与其他参数之间的相关关系rij及r[2ij]见表1。各参数营养状态指数的计算公式如下:
TLI(Chl-a)=10(2.5+1.086lnChl-a) ③
TLI(TP)=10(9.436+1.624lnTP) ④
TLI(TN)=10(5.453+1.694lnTN) ⑤
TLI(CODMn)=10(0.109+2.661lnCODMn) ⑥
式中:Chl-a单位为μg/L,TN、TP和CODMn单位均为mg/L。
采用CCA分析确定影响敞水区和滨岸带底栖动物群落结构的主要环境变量。分析前,物种密度数据经lg(x+1)转换。参与强制排序的环境变量包括AN、TN、TP、Chl-a、CODMn和TLI。采用协惯量分析(co-intertia anaysis,CoIA)探讨物种和环境变量的关系(Domínguez-haydar et al,2019;池仕运等,2021)。采用Mcnaughton优势度指数(Y)来确定底栖动物优势种(何勇凤等,2020)。当Y gt;0.02时,该物种为优势种;采用多响应置换过程(multi-response permutation procedures,MRPP)检验来评估不同群组之间的差异显著程度;采用相似性百分比分析(similarity percentage analysis, SIMPER)确定对组间差异贡献较大的分类单元。MRPP分析使用PC-ORD V7.0软件进行(Mccune amp; Mefford,1999)。SIMPER分析采用PAST v3.0进行。绘图以及统计分析采用OriginPro 2018软件进行。底栖动物多样性采用单站物种数、Shannon-Wiener指数和Pielou均匀度指数进行测度。CCA和CoIA分析采用Canoco V5.15进行(Šmilauer amp; Lepš,2014)。
2" "结果与分析
2.1" "环境因子和富营养化评价
梅溪湖秋冬季氨氮浓度远高于春夏季;总氮浓度春秋冬季较高,夏季最低;总磷春季最低,夏季最高,秋季次之;叶绿素秋季最高,春季次之,夏季最低;高锰酸盐4个季度差异不大,春秋季略高于夏冬季;TLI秋季最高,春季次之,夏季最低。就全年均值来看,氨氮和总磷处于地表水III类水平,总氮处于地表水IV类水平,高锰酸盐平处于地表水I类水平,梅溪湖水体总体处于中营养状态(表2)。
2.2" "群落结构特征
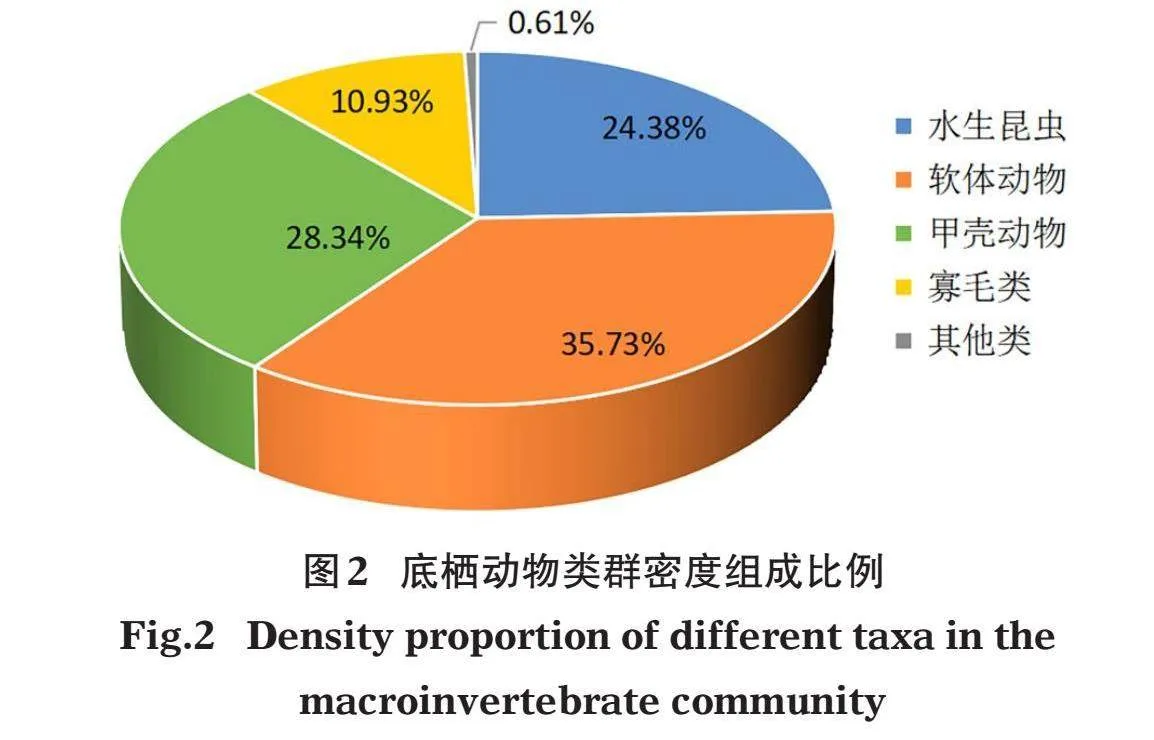
4次调查共检出底栖动物75种,隶属4门(节肢动物门、软体动物门、环节动物门和扁形动物门)9纲13目28科。节肢动物门水生昆虫有44种(其中摇蚊科34种),甲壳动物4种,蛛形纲1种;软体动物15种;环节动物门寡毛类7种,水蛭类3种;扁形动物1种。全年优势种为米虾(Caridina sp.)、铜锈环棱螺(Bellamya aeruginosa)、椭圆萝卜螺(Radix swinhoei)、大脐圆扁螺(Hippeutis umbilicalis)、霍甫水丝蚓(Limnodrilus hoffmeisteri)和沼虾(Macrobrachium sp.)。就密度占比而言,软体动物占比高达35.73%,其次是甲壳动物和水生昆虫(图2)。
春季检出底栖动物44种,夏季30种,秋季35种,冬季39种。春季优势种为霍甫水丝蚓、米虾、椭圆萝卜螺、沼虾、大脐圆扁螺和铜锈环棱螺,夏季优势种为铜锈环棱螺、米虾、赤豆螺(Bithynia fuchsiana)、纹沼螺(Parafossarulus striatulus)和椭圆萝卜螺(Radix swinhoei),秋季优势种为米虾、铜锈环棱螺、椭圆萝卜螺、云集多足摇蚊(Polypedilum nubifer)和大脐圆扁螺,冬季优势种为米虾、椭圆萝卜螺、大脐圆扁螺、沼虾、长足摇蚊(Tanypus sp.)和铜锈环棱螺。4个季度均为优势种的物种为铜锈环棱螺、米虾和椭圆萝卜螺。
滨岸带检出底栖动物61种,优势种为铜锈环棱螺、米虾、大脐圆扁螺、椭圆萝卜螺、沼虾和纹沼螺;敞水区检出42种,优势种为霍甫水丝蚓和苏氏尾鳃蚓(Branchiura sowerbyi)。经MRPP检验,滨岸带和敞水区底栖动物群落结构存在显著不同,与季节因素无关(表3)。SIMPER分析结果显示,米虾、铜锈环棱螺、霍甫水丝蚓、大脐圆扁螺、椭圆萝卜螺和沼虾是导致敞水区和滨岸带群落结构差异的主要分类单元,贡献率均超过5%,累计贡献率达55.80%。
2.3" "现存量和多样性
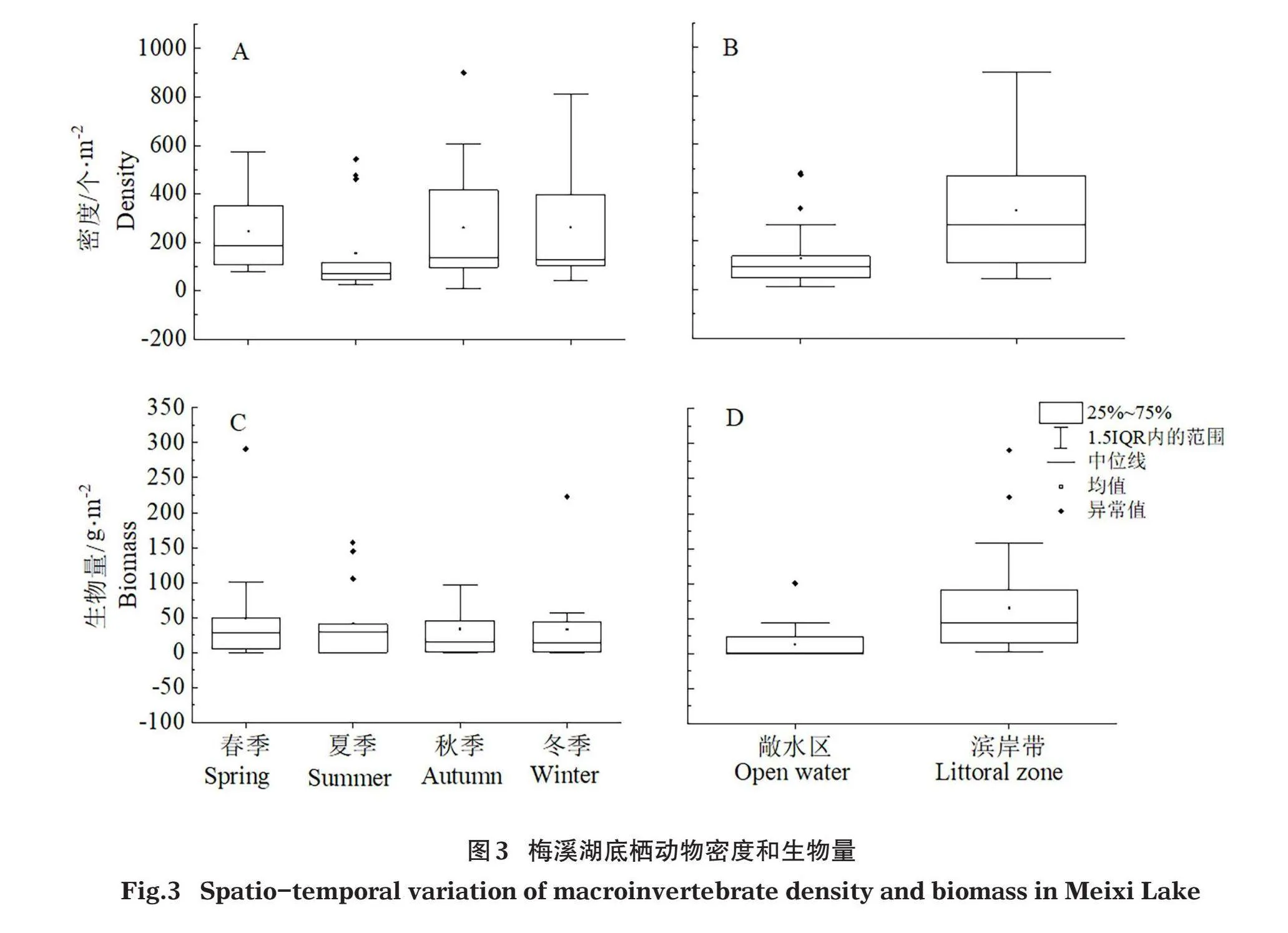
梅溪湖底栖动物密度为10.67~900个/m2,均值229.96个/m2(图3A),生物量为0.01~290.74 g/m2,均值39.67 g/m2(图3C),经单因素方差分析检验,密度和生物量季节间差异均不显著(Pgt;0.05)。梅溪湖敞水区底栖动物密度为10.67~480个/m2,均值129.21个/m2(图3B),生物量为0.01~100.45 g/m2,均值13.05g/m2(图3D);滨岸带密度为45.33~900个/m2,均值327.11个/m2(图3B),生物量为3.12~290.74 g/m2,均值65.34 g/m2(图3D),经单因素方差分析检验,敞水区和滨岸带的密度和生物量均存在显著差异(Plt;0.05)。
梅溪湖底栖动物单站物种数为1~16,均值7.45(图4A),Shannon-Wiener指数为0~2.44,均值1.49(图4B),Pielou均匀度指数为0.34~1,均值0.72(图4C),经单因素方差分析检验,单站物种数、Shannon-Wiener指数和Pielou均匀度指数季节间差异均不显著(Pgt;0.05)。梅溪湖敞水区底栖动物单站物种数为1~10,均值4.63(图4D),Shannon-Wiener指数为0~2.14,均值1.20(图4E),Pielou均匀度指数为0.43~1,均值0.82(图4F);滨岸带底栖动物单站物种数为5~16,均值10.18(图4D),Shannon-Wiener指数为1.11~2.45,均值1.77(图4E),Pielou均匀度指数为0.34~0.98,均值0.62(图4F),经单因素方差分析检验,敞水区和滨岸带的多样性指标均存在显著差异(Plt;0.05)。
2.4" "多元统计分析
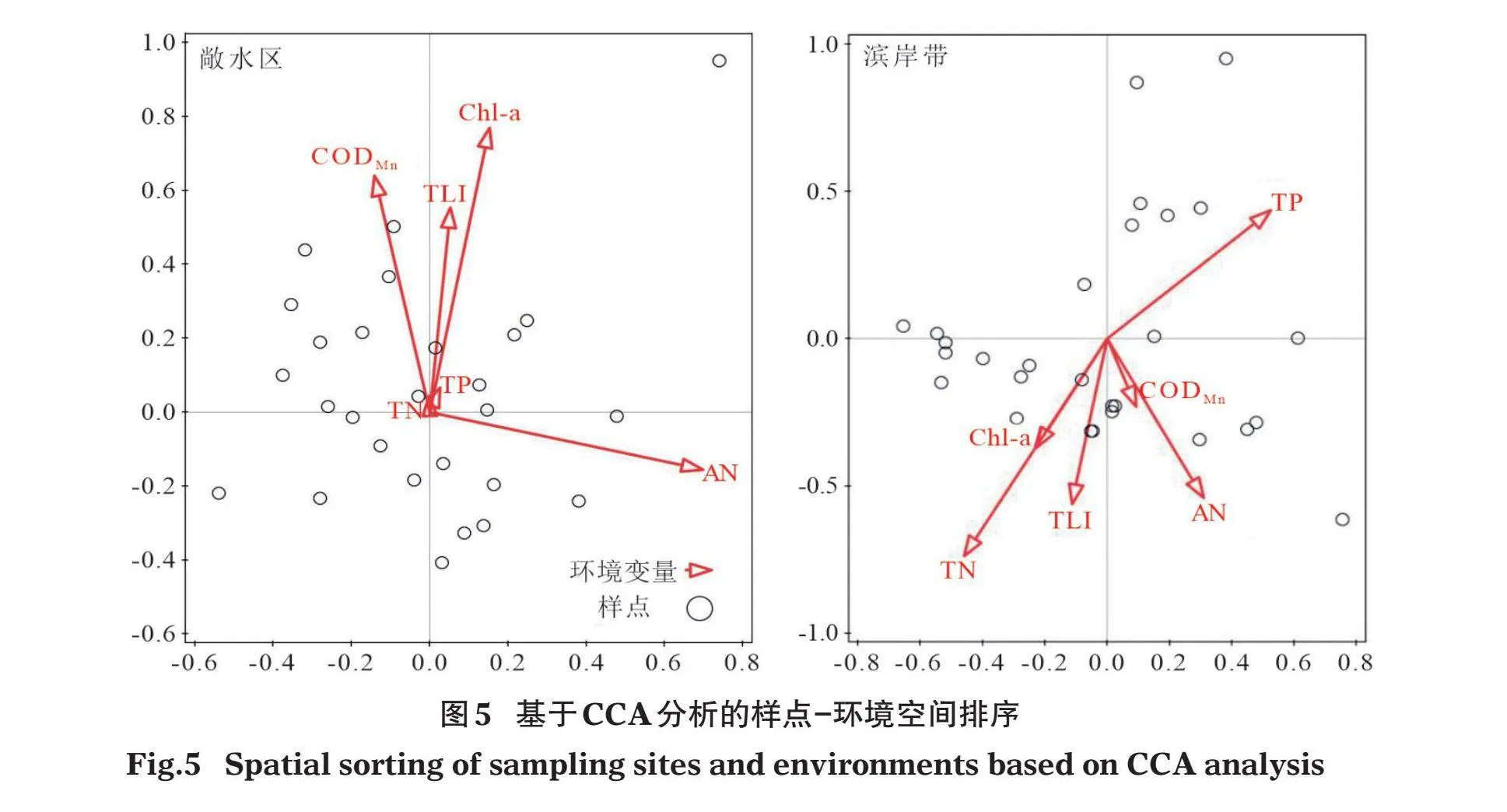
置换检验(permutation test)结果显示,敞水区和滨岸带CCA分析中环境变量所有轴均具有统计显著性(Plt;0.05),表明排序结果良好。敞水区CCA分析中,环境变量前两轴的特征值分别为0.390和0.321,累计解释了12.07%的物种变化信息,物种-环境解释拟合变化累积解释率达44.63%;滨岸带CCA分析中,环境变量前两轴的特征值分别为0.194和0.131,累计解释了15.40%的物种变化信息,物种-环境解释拟合变化累积解释率达59.62%。敞水区对底栖动物群落结构影响较大的环境因子依次为AN、Chl-a、CODMn和TLI;滨岸带对底栖动物群落结构影响较大的环境因子依次为TN、TP、AN和TLI(图5)。
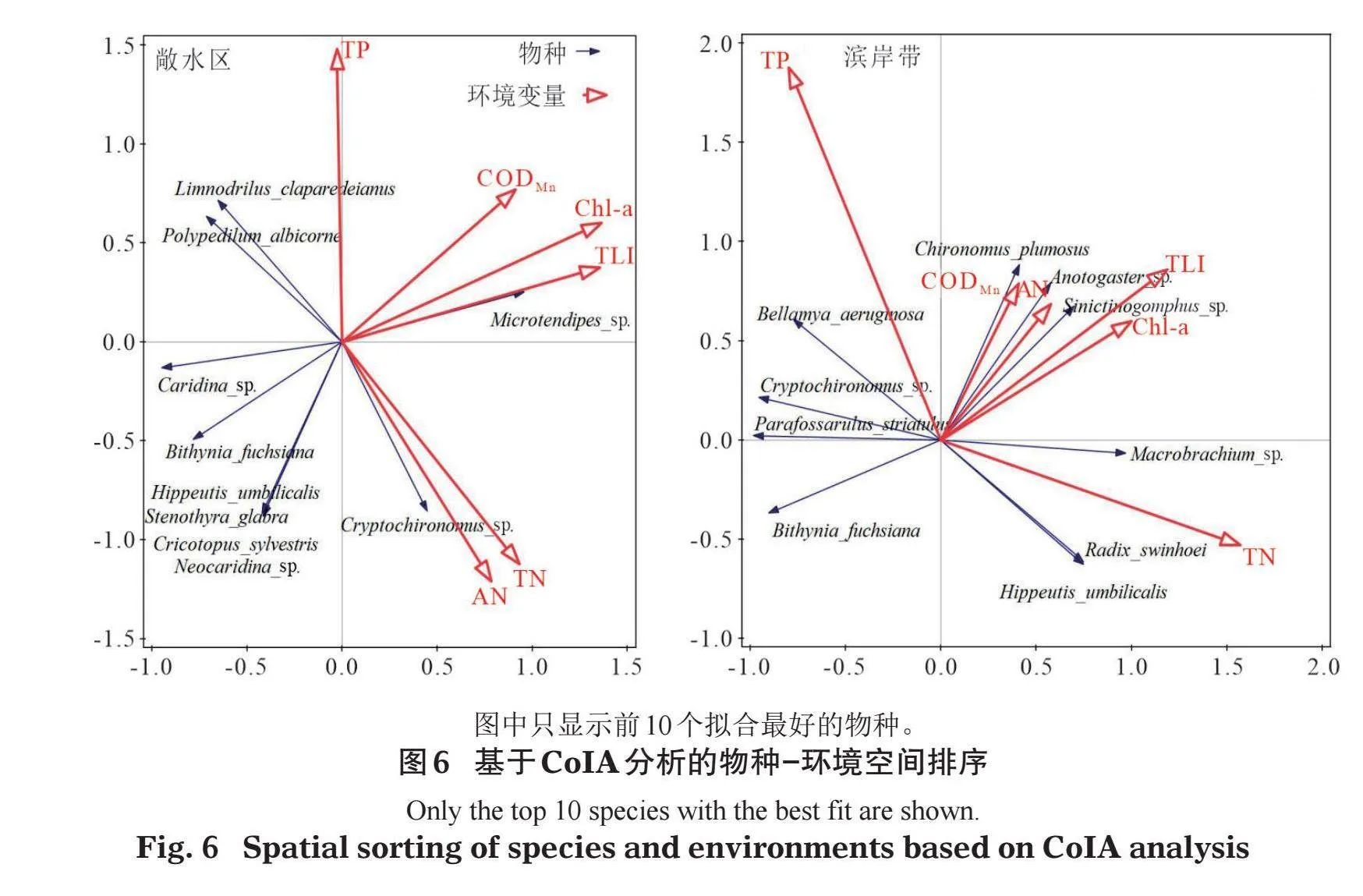
采用CoIA分析进一步探讨物种和环境变量之间的关系。敞水区的分析结果表明,隐摇蚊(Cryptochironomus sp.)随AN和TN的增加而增加,而克拉泊水丝蚓(Limnodrilus claparedeianus)和白角多足摇蚊(Polypedilum albicorne)与之相反;倒毛摇蚊(Microtendipes sp.)随CODMn、Chl-a和TLI的增加而增加,而米虾、赤豆螺、大脐圆扁螺、光滑狭口螺(Stenothyra glabra)、林间环足摇蚊(Cricotopus sylvestris)、新米虾(Neocaridina sp.)与之相反。滨岸带的分析结果表明,羽摇蚊(Chironomus plumosus)、圆臀大蜓(Anotogaster sp.)、新叶春蜓(Sinictinogomphus sp.)随CODMn、AN、TLI、Chl-a的增加而增加,而赤豆螺与之相反;沼虾、椭圆萝卜螺、大脐圆扁螺随TN的增加而增加,而铜锈环棱螺、隐摇蚊、纹沼螺与之相反;铜锈环棱螺随TP的增加而增加,而椭圆萝卜螺、大脐圆扁螺与之相反(图6)。
3" "讨论
3.1" "梅溪湖底栖动物群落结构特征
本研究发现,梅溪湖底栖动物种类丰富,群落组成较为复杂。其群落的形成壮大得益于生态工程投放的底栖动物种类(青虾、铜锈环棱螺、河蚬、河蟹等)。底栖动物种类从初期投放的几种增至75种,表明生态治理工程效果显著。研究显示,生态治理工程投放的铜锈环棱螺和青虾(沼虾)已成为全年优势种,湖区大片水草丰茂的浅水区域为米虾、大脐圆扁螺和椭圆萝卜螺提供了理想的栖息场所,是这几个物种成为全年优势种的重要因素。敞水区和滨岸带底栖动物群落结构差异显著,除霍甫水丝蚓外,全年优势种是导致敞水区和滨岸带群落结构差异的主要分类单元,说明先期投放的底栖动物种类对后续的群落结构演化起关键作用。梅溪湖运行至今,底栖动物种类大幅增加,但也带来了外来种入侵的问题,如尖膀胱螺和福寿螺。福寿螺在滨岸带水草间较为常见,出现率达到14.29%。研究表明,外来入侵种可使底栖动物群落发生根本变化,使优势种改变(Burlakova et al,2018)。对梅溪湖而言,外来入侵种的影响还需深入研究。
3.2" "环境因子对底栖动物的影响
基于直接梯度分析的CCA结果表明,AN和TLI对敞水区和滨岸带的底栖动物群落结构影响较大。虽然梅溪湖AN浓度较成湖初期大幅下降,但季节性波动较大且湖区差异明显,其对底栖动物群落影响较大。TLI结果表明,梅溪湖无富营养化现象,但浮游植物年平均丰度和季度平均丰度均达1×107个/L(魏秘等,2022),存在较高的藻华暴发风险。TLI值偏低可能与藻类小型化导致Chl-a含量较低有关,虽然TLI值处于较低水平,但在时空尺度上存在一定梯度(表1),影响底栖动物分布。CoIA分析结果显示,一些物种与环境因子存在较为密切的联系,如全年优势种米虾和大脐圆扁螺随CODMn、Chl-a和TLI的增加而减少,铜锈环棱螺随TP的增加而增加,沼虾和椭圆萝卜螺则随TN的增加而增加,这些物种可以作为有效的指示种来指示梅溪湖水环境条件的变化。研究表明,叶绿素a含量对欧洲浅水湖泊底栖动物的丰度有积极影响(Kornijow et al,2021)。本研究中,无论是敞水区还是滨岸带,叶绿素a含量对底栖动物的分布均具有较大的影响,且一些物种(图7)的丰度也与叶绿素a含量具有较为紧密的联系。
3.3" "水生植物对底栖动物的影响
在湖泊生态系统中,水生植物是天然的净化器(王同勋等,2014;杨飞和支崇远,2009),其对水体中氮磷的富集和转移具有明显的效果且具有克藻效应(李春龙, 2015)。在新建人工湖泊的成湖初期,水生植物缺乏、水体中营养物质过多以及风浪扰动都有利于藻类的生长,小风又有利于藻类的漂浮和聚集,因而很容易产生局部水域的藻华现象(杨飞和支崇远,2009)。在新建人工湖泊生态系统的重建阶段,水生植物进入对后续生态系统的演化至关重要,可以防止藻类的过度生长导致透明度下降、浊度增加、水质恶化,甚至水体发黑发臭等现象的发生(许金花等,2007)。在人工湖泊的生态系统修复过程中,水生植物的茎、叶可以改变局部的水动力特征,有效降低水中悬浮颗粒物的浓度及其吸附的重金属、磷和持久性有机物等,提高水体的透明度并改善水质。与此同时,水生植物的茎叶和根部为攀爬类和营穴类底栖动物提供了栖息地,也为这类型底栖动物和小型鱼类躲避捕食者提供重要的避难所(Lemes et al,2018)。在湖泊生态系统中,水生植物群落的发展可以加快湖泊生态系统的恢复和重建进程,这可以通过底栖动物群落的明显改善体现(Liu et al,2022)。本研究中,梅溪湖底栖动物的现存量和多样性季节间没有显著差异,但敞水区和滨岸带差异非常显著,且滨岸带远高于敞水区,表明水生植物为底栖动物提供了理想的栖息和避难场所,有利于底栖动物种群的发展壮大、群落结构的稳定恢复以及多样性的提高。
3.4" "梅溪湖运营维护建议
在湖泊生态系统中,水位变化通过影响底质的可用性来影响底栖动物群落,适度的水位有助于维持底栖动物的多样性(Lemes et al,2018;Shrestha et al,2021)。梅溪湖水位常年维持在35 m,变幅不超过40 cm,有利于滨岸带水生植物群落及底栖动物多样性的维持。
研究发现,中国长江泛滥平原33个湖泊中,水系连通和水生植物繁茂对底栖动物多样性具有正向效应(Jiang et al,2022)。梅溪湖应急补水工程等有利于底栖动物多样性的维持和提升。亚热带浅水湖泊中,小型杂食性鱼类的过度增殖会导致枝角类种群的崩溃和浮游动物小型化,进而减弱对浮游植物的控制,对底栖动物多样性造成不利影响(Pacheco et al,2021)。与此同时,底栖动物中寡毛类的过度增殖也会使沉积物中营养成分大量释放,通过上行效应和下行效应改变水生生物类群的群落组成(Yu et al,2020)。对梅溪湖而言,底栖动物群落结构合理可控且多样性维持在较高水平,可有效促进其生态系统的正常运转,合理调控鱼类结构促使底栖动物群落结构正向发展,是今后运营维护工作的重点。
参考文献
蔡如星, 1991. 浙江动物志[M]. 杭州:浙江科学技术出版社.
池仕运, 赵先富, 高少波, 等, 2021. 丹江口水库秋季底栖动物群落状态和空间分布及其与环境因子的关系[J].生态学报, 41(3):1229-1241.
窦明, 孟猛, 毛豪林, 等, 2019. 小型人工湖藻类变化特征及主要驱动因子研究[J].中国农村水利水电(5):77-81, 90.
何勇凤, 杨德国, 柴毅, 等, 2020. 长湖浮游植物优势种季节演替及生态位分析[J].水生生物学报, 44(3):612-621.
金相灿, 1995. 中国湖泊环境[M].北京:海洋出版社.
李春龙, 2015. 重庆市某人工湖泊富营养化评价及水生植物修复研究[D].重庆:重庆交通大学.
陆文泽, 任仁, 饶骁, 等, 2022. 太湖流域城市湖泊大型底栖动物群落结构及影响因素研究[J].水生态学杂志, 43(4):8-15.
沈忱,2012. 太湖湖滨生态修复区底栖动物群落结构及梯度分布[D]. 南京:南京大学.
王海军, 2007. 长江中下游中小型湖泊预测湖沼学研究[D]. 武汉:中国科学院水生生物研究所.
王洪铸, 2002. 中国小蚓类研究:附中国南极长城站附近地区两新种[M]. 北京:高等教育出版社.
王同勋, 张翠英, 蔡浩, 等, 2014. 人工湖泊中悬浮物对狐尾藻生长的影响[J].节水灌溉, (9):54-56.
王以尧, 高红, 王照丽, 等, 2018. 成都市城区主要人工湖泊水质调查与富营养化水平评价[J].四川环境, 37(5):87-93.
魏复盛, 2002. 水和废水监测分析方法[M]. 4版.北京:中国环境科学出版社.
魏秘, 王瑞, 王竹, 等, 2024. 城市人工湖泊浮游植物群落结构及环境因子分析[J]. 水生态学杂志, 45(2):92-101.
吴家乐, 刘正文, 2019. 生态修复对洋澜湖富营养化与生态状态的影响[D].广州:暨南大学.
谢志才, 马凯, 叶麟, 等, 2007. 保安湖大型底栖动物结构与分布格局研究[J].水生生物学报(2):174-183.
许金花, 潘伟斌, 张海燕, 2007. 城市小型浅水人工湖泊浮游藻类与水质特征研究[J].生态科学, 26(1):36-40, 49.
杨飞, 支崇远, 2009. 喀斯特高原地区人工湖泊富营养化治理初探:以红枫湖、百花湖为例[J].安徽农业科学, 37(9):4181-4183, 4194.
张小春, 代丹, 秦成, 等, 2023. 梅溪湖表层沉积物营养盐和有机质分布特征及污染评价[J].水生态学杂志, 44(5):107-113.
张小春, 魏秘, 王辉, 等, 2021. 生态治理技术在长沙市梅溪湖的应用研究[J].人民长江, 52(10):55-61.
钟非, 刘保元, 贺锋, 等, 2007. 水生态修复对莲花湖底栖动物群落的影响[J].应用与环境生物学报, (1):55-60.
Burlakova L E," Barbiero R P," Karatayev A Y," et al," 2018. The benthic community of the Laurentian Great Lakes: analysis of spatial gradients and temporal trends from 1998-2014[J]. Journal of Great Lakes Research," 44(4): 600-617.
Domínguez-haydar Y," Velásquez E," Carmona J," et al," 2019. Evaluation of reclamation success in an open-pit coal mine using integrated soil physical," chemical and biological quality indicators[J]. Ecological Indicators," 103: 182-193.
Jiang X M," Sun X," Alahuhta J," et al," 2022. Responses of multiple facets of macroinvertebrate alpha diversity to eutrophication in floodplain lakes[J]. Environmental Pollution," 306: 119410.
Kornijow R," Dukowska M," Leszczynska J," et al," 2021. Distribution patterns of epiphytic reed-associated macroinvertebrate communities across European shallow lakes[J]. Science of the Total Environment," 760: 144117.
Lemes da silva A L," Petrucio M M," 2018. Relationships between aquatic invertebrate communities," water-level fluctuations and different habitats in a subtropical lake[J]. Environmental Monitoring and Assessment," 190: 548.
Li Z F,nbsp; García-girón J," Zhang J," et al," 2023. Anthropogenic impacts on multiple facets of macroinvertebrate α and β diversity in a large river-floodplain ecosystem[J]. Science of the Total Environment," 874: 162387.
Liu Z S," Bai G L," Liu Y L," et al," 2022. Long-term study of ecological restoration in a typical shallow urban lake[J]. Science of the Total Environment," 846: 157505.
Mao Z G," Cao Y," Gu X H," et al," 2023. Effects of nutrient reduction and habitat heterogeneity on benthic macroinvertebrate assemblages in a large shallow eutrophic lake[J]. Science of the Total Environment," 867: 161538.
Mccune B," Mefford M J," 1999. PC-ORD. Multivariate Analysis of Ecological Data[M]. Gleneden Beach," OR," US: MJM Software Design.
Morse J C," Yang L F," Tian L X," 1994. Aquatic insects of China useful for monitoring water quality[M]. Nanjing: Hohai University Press.
Obolewski K," Glinska-lewczuk K," Sidoruk M," et al," 2021. Response of Benthic Fauna to Habitat Heterogeneity in a Shallow Temperate Lake[J]. Animals ," 11:2488.
Pacheco J P," Aznarez C," Meerhoff M," et al," 2021. Small-sized omnivorous fish induce stronger effects on food webs than warming and eutrophication in experimental shallow lakes[J]. Science of the Total Environment," 797: 148998.
Shrestha S," Tachamo-shah R D," Doody T," et al," 2021. Establishing the relationship between benthic macroinvertebrates and water level fluctuation in subtropical shallow wetlands[J]. Environmental Monitoring and Assessment," 193(8): 534.
Šmilauer P," Lepš J, 2014. Multivariate analysis of ecological data using Canoco 5[M]. Second edition. New York: Cambridge University Press.
Yu J L," Xia M L," Kong M," et al," 2020. A small omnivorous bitterling fish (Acheilognathus macropterus) facilitates dominance of cyanobacteria," rotifers and Limnodrilus in an outdoor mesocosm experiment[J]. Environmental Science and Pollution Research," 27: 23862-23870.
(责任编辑" "郑金秀)
Community Structure and Influencing Factors of Benthic Macroinvertebrates
in Artificial Urban Shallow Lakes
MO Jing‐yu1, CHI Shi‐yun2, WANG Zhu1, WANG Rui2, WEI Mi2, YU Pei‐wen1,
ZENG Xiao‐kang1, PAN Jing‐yi1, HU Ju‐xiang2
(1. Hunan Xiangxin Water Environmental Protection Investment and
Construction Co., Ltd, Changsha" "410006, P.R. China;
2. Key Laboratory of Ecological Impacts of Hydraulic-projects and Restoration of Aquatic Ecosystem,
Ministry of Water Resources, Institute of Hydrogeology, MWR amp; CAS, Wuhan" "430079, P.R. China)
Abstract:Meixi Lake is a recently constructed artificial shallow lake in the Xiangjiang New Area of Changsha, Hunan Province, serving multiple ecological functions and is playing an important role in environmental improvement and economic development of the area. Since its formation, macroinvertebrates have played a crucial role in the developing ecology of the lake. In this study, we explored and analyzed the macroinvertebrate community and its influencing factors in Meixi Lake after an ecological treatment project implemented during 2017-2018. Our aim was to provide theoretical and technical support for the restoration and management of water ecosystems in urban artificial lakes. In November 2020, January, April and July 2021, quantitative and qualitative samples of macroinvertebrates and water samples for water quality analysis were collected at 14 sites in Meixi Lake, including 7 sampling sites in the open water area and 7 in the littoral zone. A total of 75 macroinvertebrate species were identified during the four investigations in Meixi Lake, with the dominance of Caridina sp., Bellamya aeruginosa, Radix swinhoei, Hippeutis umbilicalis, Limnodrilus hoffmeisteri, and Macrobrachium sp. throughout the year. Species richness in the littoral zone (61 species) was higher than that in the open area (42 species). There were significant differences in the macroinvertebrate community structure between the littoral zone and open water, attributed to the dominant species, except for Limnodrilus hoffmeisteri. There were no significant seasonal differences in the macroinvertebrate density, biomass and community diversity, but the differences between the open water and littoral zone were significant. They were much higher in the littoral zone, indicating that construction of the aquatic plant community increased macroinvertebrate population growth and community diversity. Canonical correlation analysis of the sampling sites and environment shows that both ammonia nitrogen and the comprehensive trophic level index (TLI) had a significant impact on the macroinvertebrate communities in both the open water and the littoral zone. CoIA analysis shows that species like Caridina sp., Hippeutis umbilicalis, Bellamya aeruginosa, Macrobrachium sp., and Radix swinhoei, were closely related to environmental factors, which could serve as effective indicator species to water environment conditions in Meixi Lake. In conclusion, the macroinvertebrate community structure in Meixi Lake is reasonable and controllable, and community diversity maintains at a relatively high level, ensuring normal functioning of the aquatic ecosystem.
Key words:littoral zone; open water area; aquatic plant; macroinvertebrates; eutrophication; influencing factors; urban lake
