江西官山大样地优势种群空间分布格局与种间关联性分析
2023-12-27易伶俐戴宇峰欧阳园兰方平福彭巧华钟曲颖曾小霞丁永刚
易伶俐,方 芳,陈 琳,戴宇峰,欧阳园兰,熊 勇,方平福,彭巧华,钟曲颖,曾小霞,丁永刚*
江西官山大样地优势种群空间分布格局与种间关联性分析
易伶俐1,方 芳2,陈 琳1,戴宇峰1,欧阳园兰1,熊 勇1,方平福1,彭巧华1,钟曲颖1,曾小霞3,丁永刚1*
(1. 江西官山国家级自然保护区管理局,江西 宜春 336000;2. 江西省林业科技推广和宣传教育中心,江西 南昌 330199;3. 江西农业大学 林学院,江西 南昌 330045)
【目的】为探讨江西官山自然保护区常绿阔叶林的树种格局特征。【方法】以江西官山常绿阔叶林动态监测样地的调查数据为基础(12 hm2),根据树木点坐标信息研究优势种的分布格局、代际关系及种间关系。【结果】(1)样地中优势树种主要表现为聚集分布,在25 m以内聚集分布的树种占总树种数的90%,但随着尺度的增加各树种逐渐转为随机分布;(2)优势树种中多数常绿树种的大树与小树呈正相关,如虎皮楠、木荷;而落叶树种的大树与小树呈负相关,如赤杨叶、南酸枣;(3)优势种间主要以负关联的形式存在,呈正相关的仅有5个种对。【结论】研究结果说明官山样地群落还处于发育初期,优势种主要以聚集分布为主,且种间竞争激烈。
优势种;分布格局;代际关系;种间关联;江西官山自然保护区
【研究意义】优势种群是指对群落的结构和群落的环境有明显控制作用的植物,分析优势树种的种群特征,有助于对群落结构的认识。空间分布格局是植物自身生物学特性与外界因素综合作用的产物;是植物个体在与环境相互作用中所形成的空间配置特点,种群的分布格局分为聚集分布、随机分布、均匀分布3种类型。空间分布格局的研究对于确定种群特征、代际(种内)和种间相互作用,以及种群与环境之间的关系具有非常重要的作用,对生物多样性保护及种群的保育具有一定的指导意义[1]。不同物种在空间上的分布格局关系为种间联结[2],分为正联结、负联结、无关联3种情况。其中,正联结是指两个物种对环境资源的需求相同或相似,是群落在演替过程中各物种对环境的趋同性和竞争性引起的;负联结及无关联是指两个物种相互独立,几乎不在同一时空出现[3-4]。种间联结能够反应物种与生境因子之间的耦合关系,是群落演替机制研究的重要特征之一[5-7]。对种间联结性的研究,有助于加强人们对群落结构、功能和动态演替过程的认识。【前人研究进展】因此,许多学者对物种的分布格局、代际(种内)和种间相互作用进行了大量研究,如樊登星[8]、李立[9]、张育新[10]等分别研究了北京山区栓皮栎(),古田山甜槠()与北京东灵山辽东栎()的种内代际关系。陈绪辉等[5]、刘红润和李凤日[6]、吉颖等[7]分别以油松()、红松()、木贼麻黄()等珍稀濒危物种为研究对象,分析其与群落内主要物种的种间关联性。【本研究切入点】这些研究对种群的发育和珍稀濒危植物的保护具有重要意义,但也存在一定的局限性,研究的尺度较小,或者只分析了单一尺度下的分布格局、代际和种间关联性。种群的空间分布格局和种间关系存在着尺度的依赖性,不同尺度下分布格局、代际关系、种间联结性变化显著,在小尺度上受扩散限制和种间竞争的影响,在大尺度上主要受生境异质性的影响[11-12]。1977年Ripley提出的点格局分析方法[13-14],它是以植物种的个体在空间的坐标为基本数据[15],可以研究不同尺度的分布格局和种间关系。因此,有必要对常绿阔叶林开展大样地建设工作,并采用点格局分析方法研究物种分布格局、代际和种间关系。官山国家级自然保护区赣西北九岭山脉西段,森林群落以常绿阔叶林为主,具有较丰富的生物多样性,是开展常绿阔叶林研究的理想地点。【拟解决的关键问题】本研究以生物多样性较丰富的官山自然保护区为研究地点,对常绿阔叶林群落进行详细调查,并采用点格局中的异质性泊松模型来分析不同优势种的分布格局、代际关系及种间联结性。主要探讨以下几个科学问题:样地内优势种的空间分布格局特征、大树与小树的空间关系、哪些树种的大树会影响小树生长及优势种间的联结性的状态等。这些科学问题的解决,将有助于人们对该区域植物群落的认识和了解,更利于今后对其植物多样性的保护。
1 研究区域概况
江西官山自然保护区位于赣西北九岭山脉西段,总面积11 500.5 hm2,其中核心区3 621.1 hm2,缓冲区1 466.4 hm2,实验区6 413.0 hm2;地理坐标为28°30′~28°40′N,114°29′~114°45′E,最高海拔1 480 m(麻姑尖),最低海拔200 m;坡度30°左右的地区面积最大,最陡的区域出现在海拔500~800 m[16],年均气温16.2 ℃,年均降水量1 950~2 100 mm,四季分明、光照充足,属中亚热带暖湿东南季风气候区[17]。土壤类型共有红壤、黄壤、草甸土3个类型,依次分布于保护区海拔500 m以下、500~1 300 m、1 300 m以上的山顶。
其独特的地理位置和复杂的地貌特征,孕育着丰富的植被类型和野生动植物种类。森林覆盖率高达93.8%,高等植物2 344种,被子植物1 896种,国家级重点保护野生植物21种和国家级珍稀濒危保护植物28种,分别占江西省的45.8%、46.4%、38.1%和51.8%[18]。
2 研究方法
2.1 样地概况
为了监测本区植被组成结构及功能变化,于2014年按CTFS样地调查方法[19],在国家财政部、生态环境部“全国生物多样性野外监测示范基地修缮项目”支持下,江西农业大学和江西官山国家级自然保护区管理局共同在江西官山国家级自然保护区东河管理站附近建立了12 hm2(300 m×400 m)的亚热带常绿阔叶林动态监测样地。样地南北走向,西南角(原点)地理坐标为28°33′25″N,114°34′40″E,整个样地地形复杂,样地东北高,西南低,最高海拔645.0 m,最低海拔444.1 m,相对高差200.9 m,局部地段坡度在60°以上(图1)。
基于2014年的调查数据表明,样地内共有木本植物312种63 690株(DBH≥1 cm),其中阔叶树304种56 249株(常绿142种39 741株;落叶162种16 508株)。样地内优势种明显,重要值位于前10的依次是:杉木()、赤杨叶()、毛竹()、虎皮楠()、鹿角杜鹃()、小叶青冈()、木荷()、南酸枣()、麻栎()、红楠(),这10个树种的重要值约占总树种的一半。
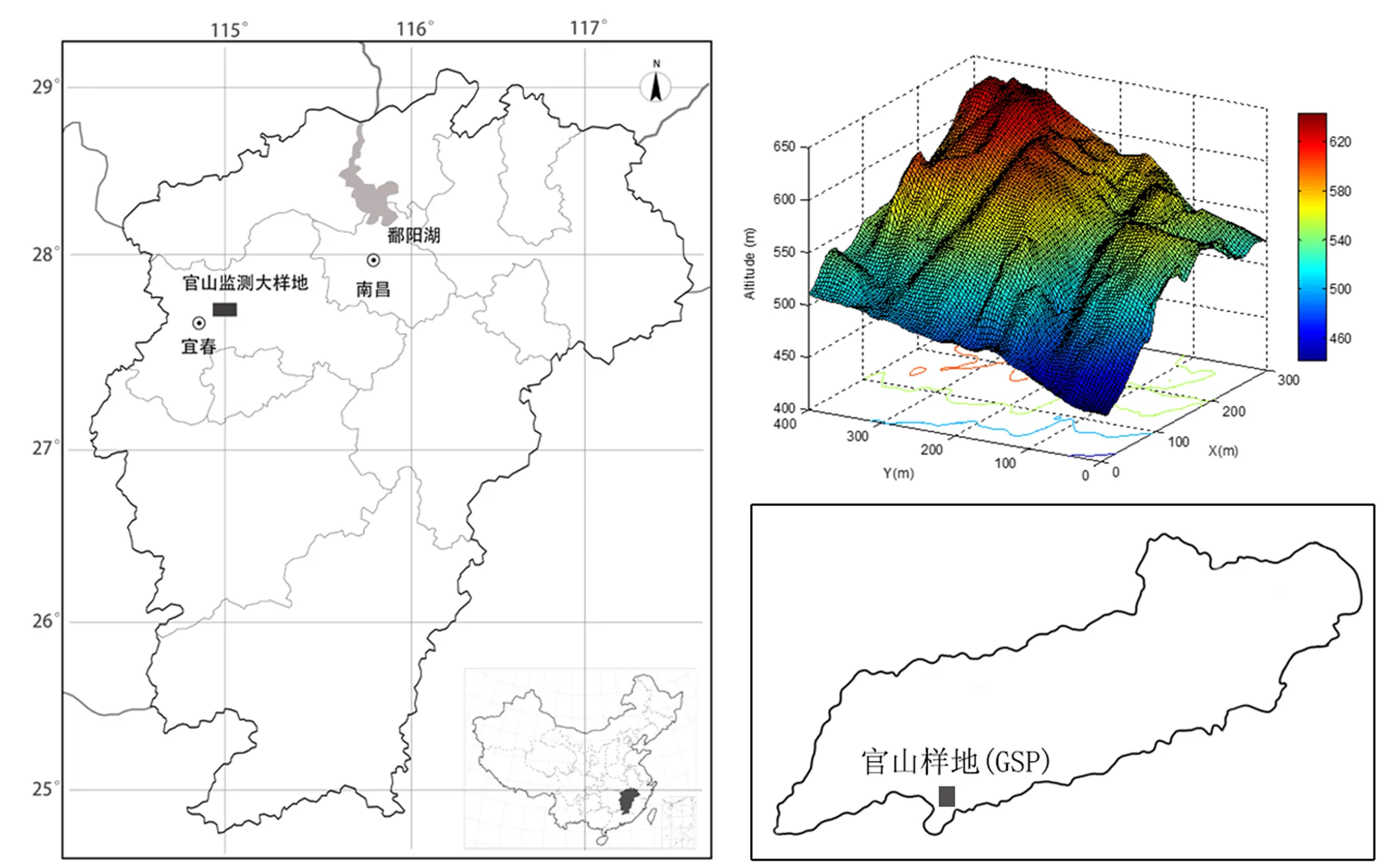
图1 官山大型森林监测样地地理位置与数据高程模型
2.2 数据处理
2.2.1 空间分布格局与种间关联性 以江西官山自然保护区大型森林监测样地中的重要值位于前10的优势种(杉木、赤杨叶、毛竹、虎皮楠、鹿角杜鹃、小叶青冈、木荷、南酸枣、麻栎、红楠)为研究对象,分析其分布格局、种内与种间关联性。
点格局分析过程中的置信区间用包迹线来表示,通过重复Monte Carlo随机模拟99次产生99%的置信区间。当树种的g11()值高于包迹线上限,则该树种在该距离为聚集分布;若在包迹线之间,则为随机分布;低于包迹线的下限,则为均匀分布[20-21]。在双变量分析中,若g12()值在包迹线之间,则种间关系在该距离不相关;若g12()值高于包迹线上限或低于包迹线的下限,则种间关系在该距离为正相关或负相关。本研究采用的零模型主要为异质性泊松模型,是空间点格局中使用最广泛的一种,该模型假设任何一个个体在研究区域内任何一个位置上出现的几率相等,同时点与点之间是彼此独立的,在生态学中常用来假设物种的空间分布不受任何生物或非生物过程影响。统计分析采用Programita软件分析实现。计算公式如下:


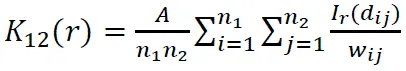
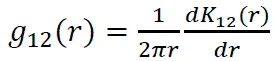
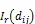
该研究所有分析采用Excel 2016、Mtalab 2018a、Origin 8.5以及Programita软件分析实现。
2.2.2 代际关系分析 按DBH(胸径)和H(树高)两个指标,将9个树种分成大树(DBH>5 cm且>5 m)与小树(DBH≤5 cm且≤5 m)。采用点格局的分析方法进行数据处理,分析优势种的种内关联性(计算方式与种间关系计算方式相同,由于毛竹不会进行次生生长,未进行种内关联性分析)。
3 结果与分析
3.1 优势种群分布格局
采用异质性泊松模型分析得出官山国家自然保护区12 hm2内,重要值位于前10的树种,在25 m以内以聚集分布为主,约占90%;45 m以外随机分布占据主导地位(图2)。10个优势树种在0~150 m的尺度内,随尺度的增加聚集分布的树种数呈先急剧下降后上升的趋势,在45 m的尺度上降为0,后缓慢上升至3左右且保持稳定;随机分布的树种数呈先急剧上升后下降的趋势,在45 m左右达到峰值9,后降低至7且趋于稳定;均匀分布在约40 m内为0,后上升到3左右且保持稳定。
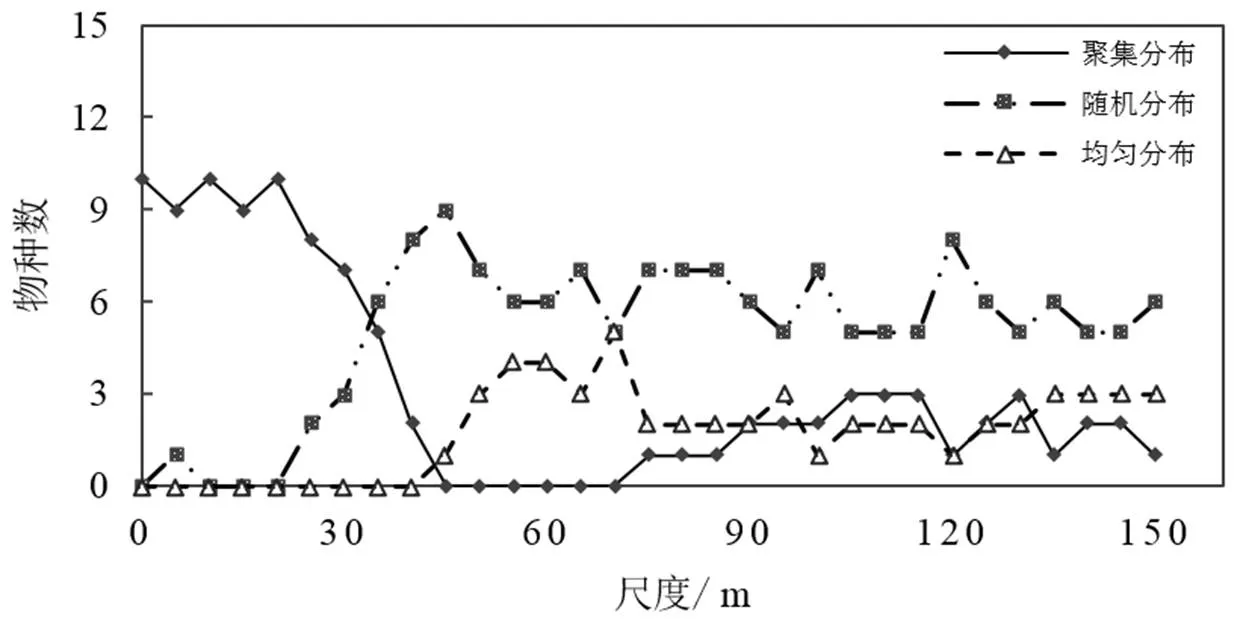
图2 不同尺度上10个优势种的分布格局
由图3中树种分布的散点图可以发现除了赤杨叶、虎皮楠、南酸枣、红楠在样地中的分布较均匀,其它树种集群现象较为明显外,杉木主要分布在山脊和缓坡周围;毛竹、鹿角杜鹃主要分布于阴坡;小叶青冈和木荷主要分布于阴坡;麻栎主要分布于阳坡。因此采用异质性泊松模型排除环境空间异质性的影响。结果显示:麻栎在整个样地基本以随机分布的状态出现。其它9个树种约在0~30 m内呈聚集分布,30 m之后,杉木和红楠呈随机分布-均匀分布-聚集分布交替出现;南酸枣、赤杨叶、木荷、虎皮楠、鹿角杜鹃大体呈随机分布;毛竹基本呈均匀分布。

实线为实际观测值,虚线之间为置信区间,子图为树种在样地中的实际分布图,子图中圆圈的大小表示植物个体胸径的大小。
3.2 优势种群代际关系
代际关系在一定程度上能够反映种群的更新特征,官山大样地9个优势树种中多数树种的大树与小树能够共存,但共存的尺度存在差异(图4)。具体表现为:杉木、虎皮楠、鹿角杜鹃在0~150 m的尺度上大树与小树基本呈现正相关关系;麻栎0~150 m的尺度上基本不相关;木荷、小叶青冈、红楠的大树与小树在0~25 m内基本呈现正相关的关系,在50~150 m内基本呈负相关;而赤杨叶与南酸枣的种内关联性在0~100 m内主要呈现负相关。
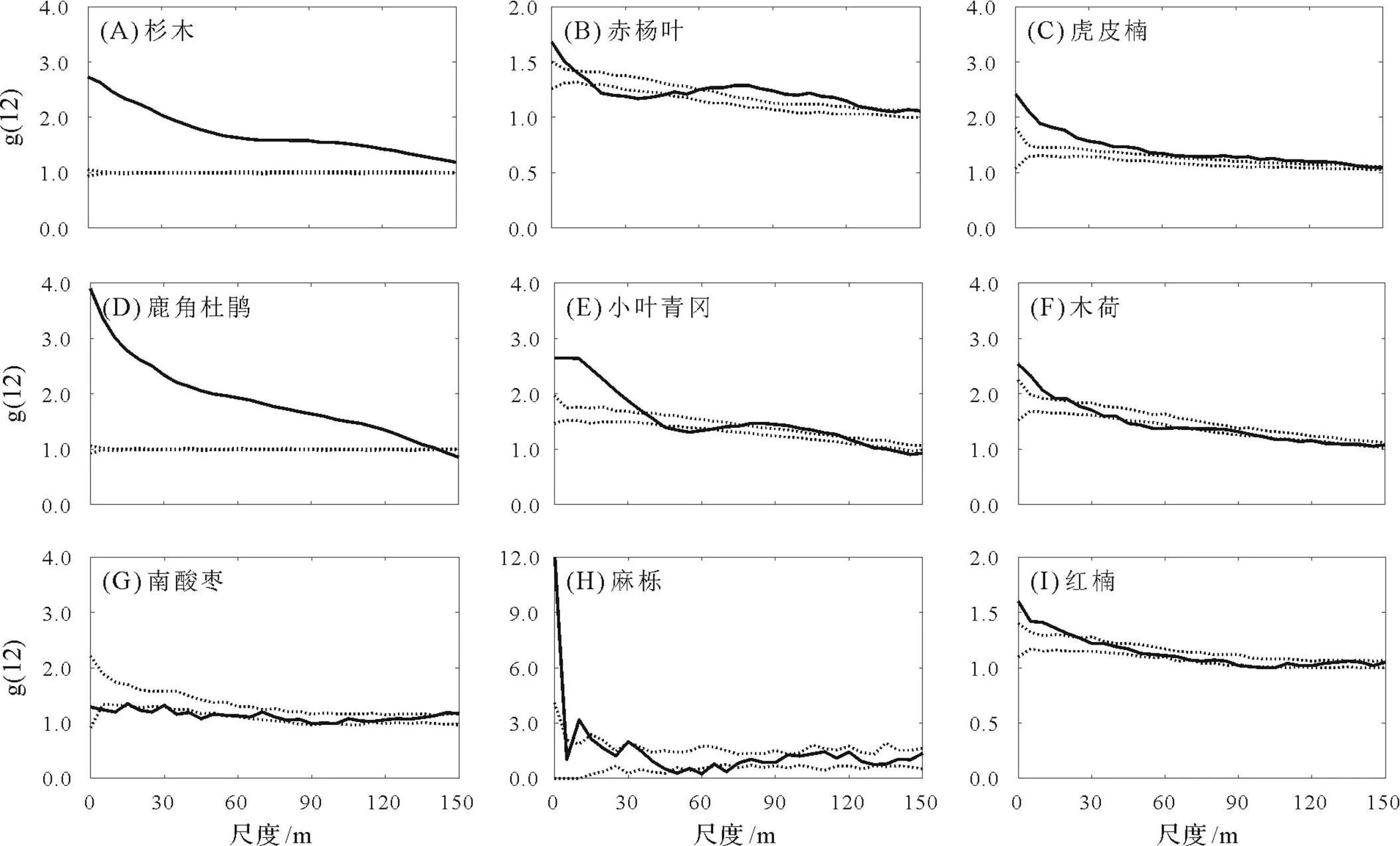
图4 优势种群的代际关系
3.3 优势种群种间关系
对样地中的杉木、赤杨叶、毛竹、虎皮楠、鹿角杜鹃、小叶青冈、木荷、南酸枣、麻栎、红楠10个树种45个种对的关联类型进行分析。结果显示:样地中大部分优势树种在空间分布上呈现负关联性,随尺度的变化负关联的种对持续高于正关联和不相关的种对,仅在150 m的尺度上略低于不相关的种对(图5)。负相关种对在约50 m的尺度上达到了其最高值(约占76%),之后呈下降趋势;正相关和不相关的树种在0~80 m的尺度上所占的比例均不超过20%,在80 m之后正相关对数和不相关对数有增加的趋势,其中正相关的树种在约145 m尺度上所占的比例最高(约占36%),不相关的树种在150 m尺度上所占的比例最高(约占38%)。
45个种对在不同尺度的种间关系如图6所示,在整个样地范围内完全呈负相关的有12对,占总对数的26.67%,分别是杉木-鹿角杜鹃、杉木-毛竹、小叶青冈-木荷、小叶青冈-杉木、木荷-虎皮楠、木荷-红楠、木荷-毛竹、木荷-鹿角杜鹃、南酸枣-小叶青冈、南酸枣-鹿角杜鹃、南酸枣-毛竹、麻栎-木荷;基本以负相关状态存在的有13对,占28.89%,如:小叶青冈-赤杨叶、南酸枣-木荷、虎皮楠-杉木等。呈正相关的仅有5个种对,分别是红楠-毛竹、小叶青冈-毛竹、鹿角杜鹃-毛竹、赤杨叶-杉木、南酸枣-杉木。几乎不相关的种对有2个,是南酸枣-虎皮楠、南酸枣-麻栎。其它13个种对,仅在部分区域存在正相关或不相关的情况,其余范围均为负相关状态,如:赤杨叶-虎皮楠、麻栎-毛竹、红楠-杉木等。

图中A、B、C表示负相关的种对;D、E、F表示正相关的种对;G、H表示不相关的种对;I表示大体负相关的种对;J、K、L表示正相关、负相关、不相关交替出现的种对。
4 讨 论
4.1 优势种群主要呈聚集分布
种群分布格局是物种与环境长期相互作用的结果,生物因子和非生物因子均可影响种群的分布格局。官山大型森林动态监测样地的优势树种在25 m以内主要以聚集分布为主,这符合自然群落种群聚集分布的一般规律。导致物种呈聚集分布的主要因素是生境异质性和限制性扩散[20-22]。在较小尺度上,物种的分布格局主要由密度制约(种内种间竞争)、种子的扩散机制决定[23];而较大尺度上的空间分布格局则更多受到生境异质性的影响[24]。
此研究目标树种中小叶青冈、南酸枣、木荷等果实较大,成熟的种子多掉落在母树周边,且种子的传播方式以动物传播为主,因此导致这些树种呈聚集分布。另外,有趣的是麻栎的果实也较大,但其分布格局在0~150 m的尺度上均呈随机分布,这与其生物学特性和种群发育阶段有关。麻栎喜阳,其幼苗在林下更新困难[25],且麻栎在本研究区域发育的较成熟,大树占比高(麻栎胸径最大的个体为83.1 cm,胸径大于20 cm的个体占总个体数的79.70%),这不利于幼树的生长,导致相邻植株由同种个体变为异种个体,因此其分布格局呈随机分布。综述所述种子扩散限制与萌蘖繁殖是官山大样地主要树种在小尺度上聚集分布的主要原因,群落发育阶段会影响种群的分布格局。
此外,样地中的优势种多数具有明显的生境偏好,不同树种在不同生境形成不同的斑块,但在采用异质性泊松模型排除环境空间异质性的影响[20,26]后,各树种在25 m的尺度上以聚集分布为主,这说明生境抑制性不是导致此研究区域树种在大尺度下聚集分布的主要原因。Getzin等[24]研究表明在大于10 m的尺度上,如果呈现聚集分布,就可以解释为是受到生境异质性的影响,导致官山大样地树种聚集分布的原因还需进一步探索。
4.2 优势种群种内竞争较弱
代际关系是影响植物空间分布格局的重要因素。本研究发现在0~25 m内虎皮楠、鹿角杜鹃、木荷等常绿阔叶树的大树与小树间均表现为正关联,而赤杨叶、南酸枣等落叶阔叶树的大树与小树在0~100 m内,基本呈现负相关的关系。这可能与它们生物学特性有关,赤杨叶、南酸枣等属于阳性落叶树种,具有喜阳且生长迅速的生态学特征[27-28],而随着群落的演替和阳性阔叶树的生长,林分郁闭度逐渐增加,不利于其林下小树的生长与更新,因此赤杨叶、南酸枣等落叶阔叶树的大树与小树间呈负相关关系。而虎皮楠、木荷等常绿阔叶树具有耐荫性,林分郁闭度的增加有利于其幼苗、幼树的生长与更新,因此他们的大树与小树间呈正相关关系。
4.3 优势种群种间竞争激烈
种间联结是用来表示不同物种在空间分布上的相互关联性的,有正关联、负关联和无关联3种形式。结果显示10个优势种种间关联,主要以负关联的形式存在,而空间分布上表现为负关联的原因主要有2个,一是物种间生态位过渡重叠而产生强烈的种间竞争;二是对生态环境的需求完全不同,生态习性不相似[29]。由分布格局可知,本研究的杉木主要分布在山脊和缓坡周围;毛竹、木荷、小叶青冈、鹿角杜鹃主要分布于阴坡;麻栎主要分布于阳坡。可见优势种间以负关联的形式存在与其对生境的偏好有关,但是否受生态位的重叠情况影响还需进一步分析。
本研究以官山12 hm2的常绿阔叶林为研究对象,具体分析了不同尺度下常绿阔叶林优势种的分布格局、代际和种间关系,发现优势种主要呈聚集分布,种内竞争较弱,种间竞争较激烈,但随着研究尺度的增加,物种的分布格局由聚集分布朝随机分布转变,负关联的种对呈增加的趋势,正关联种对呈增加趋势。这有助于了解物种间相互关系的尺度效应,为常绿阔叶林植物群落演替动态的研究提供参考依据。
[1] 袁春明, 孟广涛, 方向京, 等. 珍稀濒危植物长蕊木兰种群的年龄结构与空间分布[J]. 生态学报, 2012, 32(12): 3866-3872.
[2] 刘翔宇, 何东, 田文斌, 等. 浙江普陀山岛森林木本植物的种间关联格局[J]. 植物生态学报, 2017, 41(12): 1219-1227.
[3] 郑振宇, 龙翠玲. 茂兰喀斯特森林不同地形部位木本植物种间联结性分析[J]. 林业资源管理, 2014(4): 78-84.
[4] 张金屯, 焦蓉. 关帝山神尾沟森林群落木本植物种间联结性与相关性研究[J]. 植物研究, 2003(4): 458-463.
[5] 陈绪辉, 叶宝鉴, 潘标志, 等. 珍稀濒危植物江南油杉群落乔木层主要树种种间关联性分析[J]. 热带亚热带植物学报, 2023, 31(1): 21-30.
[6] 刘红润, 李凤日. 红松天然林种内和种间竞争关系的研究[J]. 植物研究, 2010, 30(4): 479-484.
[7]吉颖, 许强, 张钦弟, 等. 山西五鹿山自然保护区木贼麻黄群落优势种的种间关联性分析[J]. 西北植物学报, 2017, 37(5): 1004-1011.
[8] 樊登星, 余新晓. 北京山区栓皮栎林优势种群点格局分析[J]. 生态学报, 2016, 36(2): 318-325.
[9] 李立, 陈建华, 任海保. 古田山常绿阔叶林优势树种甜槠和木荷的空间格局分析[J]. 植物生态学报, 2010, 34(3): 241-252.
[10] 张育新, 马克明, 祁建, 等. 北京东灵山海拔梯度上辽东栎种群结构和空间分布[J]. 生态学报, 2009, 29(6): 2789-2796.
[11] 王慧杰, 常顺利, 张毓涛, 等. 天山雪岭云杉森林群落的密度制约效应[J]. 生物多样性, 2016, 24(3): 252-261.
[12] 梁爽, 许涵, 林家怡, 等. 尖峰岭热带山地雨林优势树种白颜树空间分布格局[J]. 植物生态学报, 2014, 38(12): 1273-1282.
[13] 梅世秀, 袁志良, 叶永忠. 宝天曼自然保护区锐齿栎种群空间分布格局研究[J]. 河南农业大学学报, 2011, 45(2): 220-225.
[14] 袁志良, 王婷, 朱学灵, 等. 宝天曼落叶阔叶林样地栓皮栎种群空间格局[J]. 生物多样性, 2011, 19(2): 224-231.
[15] 张金屯, 孟东平. 芦芽山华北落叶松林不同龄级立木的点格局分析[J]. 生态学报, 2004(1): 35-40.
[16] 徐定兰, 余泽平, 王国兵, 等. 江西官山常绿阔叶林监测样地: 灌木多样性及空间计量特征[J]. 江西农业大学学报, 2018, 40(5): 1001-1011.
[17]曹岚, 梁芳, 邹红, 等. 江西官山国家级自然保护区珍稀植物多样性研究[J]. 安徽农业科学, 2012, 40(3): 1696-1698.
[18] 刘信中, 吴和平. 江西官山自然保护区科学考察与研究[M]. 北京: 中国林业出版社, 2005.
[19] 兰国玉. 世界热带森林生态系统大样地定位研究进展[J]. 西北植物学报, 2007, 27(10): 2140-2145.
[14] 张金屯. 植物种群空间分布的点格局分析[J]. 植物生态学报, 1998(4): 57-62.
[20] 陈云, 王婷, 李培坤, 等. 河南木札岭温带落叶阔叶林群落特征及主要乔木空间分布格局[J]. 植物生态学报, 2016, 40(11): 1179-1188.
[21] 王晓雨, 于大炮, 周莉, 等. 长白山北坡林线岳桦种群空间分布格局[J]. 生态学报, 2015, 35(1): 116-124.
[22] 王志高, 张中信, 段仁燕, 等. 鹞落坪国家级自然保护区吊罐井物种分布格局与种间关联的多尺度分析[J]. 植物科学学报, 2016, 34(1): 21-26.
[23] 饶米德, 冯刚, 张金龙, 等. 生境过滤和扩散限制作用对古田山森林物种和系统发育β多样性的影响[J]. 科学通报, 2013, 58(13): 1204-1212.
[24] GETZIN S, WIEGAND T, WIEGAND K, et al. Heterogeneity influences spatial patterns and demographics in forest stands[J]. Journal of ecology, 2008, 96(4): 807-820.
[25] 卢正茂. 坡向与林分密度对麻栎林生物量的影响[J]. 技术研究, 2021(7): 72-73.
[26] 陈云, 郭凌, 姚成亮, 等. 暖温带-北亚热带过渡区落叶阔叶林群落特征[J]. 生态学报, 2017, 37(17): 5602-5611.
[27] 田有圳. 天然混交林中拟赤杨的生长规律[J]. 北华大学学报(自然科学版), 2009, 10(1): 76-79.
[28] 吴小龙. 南酸枣生长进程研究[J]. 江苏林业科技, 2003, 30(6): 6-8.
[29] WIEGAND T, MOLONEY K A. Rings, circles, and null-models for point pattern analysis in ecology[J]. Oikos, 2004, 104(2): 209-229.
Spatial Pattern and Interspecific Correlation Analysis of Dominant Species of Guanshan Evergreen Broadleaved Forest Monitoring Plot in Jiangxi Province
YI Lingli1, FANG Fang2, CHEN Lin1, DAI Yufeng1, OUYANG Yuanlan1, XIONG Yong1, FANG Pingfu1, PENG Qiaohua1, ZHONG Quying1, ZENG Xiaoxia3, DING Yonggang1*
(1. Administration of Jiangxi Guanshan National Nature Reserve, Yichun, Jiangxi 336000, China; 2. Extension and Publicity Education Center for Forestry Technology of Jiangxi, Nanchang 330199, China; 3. School of Forestry Sciences, Jiangxi Agricultural University, Nanchang 330045, China)
This study aimed to explore the population pattern characteristics of evergreen broad-leaved forest in Guanshan National Nature Reserve, Jiangxi Province.Based on the investigation data of the dynamic monitoring sample plot of evergreen broad-leaved forest in Guanshan (12 hm2), Jiangxi Province, the distribution pattern, intergenerational relationship and interspecific relationship of dominant species in the sample plot were analyzed, according to the coordinate information of tree points.(1) The dominant species were mainly concentrated, and 90% of the species were concentrated within 25 m, but with the increase of the scale, the tree species gradually turned to random distribution; (2) Among the dominant tree species, the large trees of most evergreen tree species were positively correlated with small trees, such asand; while, the large trees and small trees of deciduous species were negatively correlated, such asand; (3) The dominant species mainly existed in the form of negative correlation, and only 5 species pairs were found to be positively correlated.The community is still in the early stage of development, the dominant species are mainly concentrated and distributed, and the interspecific competition is fierce.
dominant species; distribution pattern; intergenerational relation; interspecific association; Jiangxi Guanshan National Nature Reserve
10.3969/j.issn.2095-3704.2023.04.76
S718.54+1
A
2095-3704(2023)04-0510-09
2023-06-15
2023-08-25
官山保护区植物群落类型多样性调查项目(2022)
易伶俐(1985—),女,工程师,主要从事自然保护区管理工作,274317646@qq.com;*通信作者:丁永刚,2389173159@qq.com。
易伶俐, 方芳, 陈琳, 等. 江西官山大样地优势种群空间分布格局与种间关联性分析[J]. 生物灾害科学, 2023, 46(4): 510-518.
